黄海北部春季和夏季浮游动物生态特性与时空分布
2017-12-18王爱勇王小林刘修泽
段 妍,王爱勇,王小林,刘修泽,柴 雨,陆 阳,2,王 彬,董 婧
( 1.辽宁省海洋水产科学研究院,辽宁省海洋生物资源与生态学重点实验室,辽宁 大连 116023; 2.大连海洋大学,辽宁 大连 116023 )
黄海北部春季和夏季浮游动物生态特性与时空分布
段 妍1,王爱勇1,王小林1,刘修泽1,柴 雨1,陆 阳1,2,王 彬1,董 婧1
( 1.辽宁省海洋水产科学研究院,辽宁省海洋生物资源与生态学重点实验室,辽宁 大连 116023; 2.大连海洋大学,辽宁 大连 116023 )
根据2015年3月(春季)和6月(夏季)对黄海北部辽宁近岸海域的浮游动物调查结果,分析了两个季节的浮游动物种类、生物量、丰度等群落结构特征。此次调查共鉴定出浮游动物47种,其中春季42种,夏季34种。浮游动物以浮游幼虫和小型桡足类为主。桡足类幼虫、棘皮动物幼体、桡足类幼体、洪氏纺锤水蚤、短角长腹剑水蚤、小拟哲水蚤、拟长腹剑水蚤和腹针胸刺水蚤为春季优势种;夏季浮游动物优势种依次为桡足类幼体、洪氏纺锤水蚤,拟长腹剑水蚤、短角长腹剑水蚤、桡足类幼虫、小拟哲水蚤和腹针胸刺水蚤。春季浮游动物平均生物量为512.09 mg/m3,平均丰度为15 522.18个/m3;夏季浮游动物平均生物量为218.84 mg/m3,平均丰度为7582.39个/m3;浮游动物生物量和丰度均为春季较高。黄海北部辽宁沿岸海域的浮游动物生物量、丰度较高。水温是影响浮游动物群落结构的重要环境因子。
黄海北部;浮游动物;生物量;丰度;群落结构;季节变化
浮游动物,作为浮游植物的摄食者,同时也为其他经济生物提供了丰富的饵料来源。浮游动物在初级生产者和更高营养级生物的能量传递中起到连接作用,在海洋生态系统中扮演着重要的角色[1-2],其种类组成和生物量的季节性变动对渔业资源及水域生态具有重要的调控作用;同时,某些种类的数量分布对海洋气候变化和洋流、水团的变动也具有一定的指示作用[3]。黄海北部既是黄海暖流进入渤海海域的通道,同时也是黄河沉积物向外海扩散的必经之路,因此黄海北部海域水动力及水文条件变化剧烈, 物质来源复杂[4-5]。辽宁省黄海北部沿岸海域是众多河流的入海口,水域饵料充足,营养丰富,特定的海洋环境多样性也造就了黄海北部海域浮游动物多样性的格局。
笔者根据2015年3月(春季)、6月(夏季)浮游动物的调查结果,分析了黄海北部辽宁近岸海域的浮游动物分布特征,丰度和生物量,优势种及物种多样性等的动态变化,为今后对辽宁省黄海北部海域海洋生态环境的保护及可持续利用提供基础资料与依据。
1 材料与方法
1.1 调查站位
黄海北部调查站位见图1。2015年3月(春季)和6月(夏季)在黄海北部海域进行了2次拖网调查,此次共设14个采样站位。
1.2 样品采集与分析
采用附有网口流量计的浅水Ⅱ型浮游生物网由底层至表层垂直拖网采集样品。用体积分数为5%的甲醛溶液固定保存于塑料样品瓶中,带回实验室进行分析。计数称量质量得出浮游动物的丰度和生物量[6]。
1.3 数据分析
采用香农—威纳指数(H′)表示多样性指数[7];采用Margalef指数(dMa)表示物种丰富度指数[8];采用Pielou指数(J)表示物种均匀度[9];采用优势度(Y)表示优势种类[10]。计算公式如下:
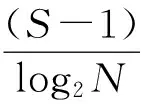
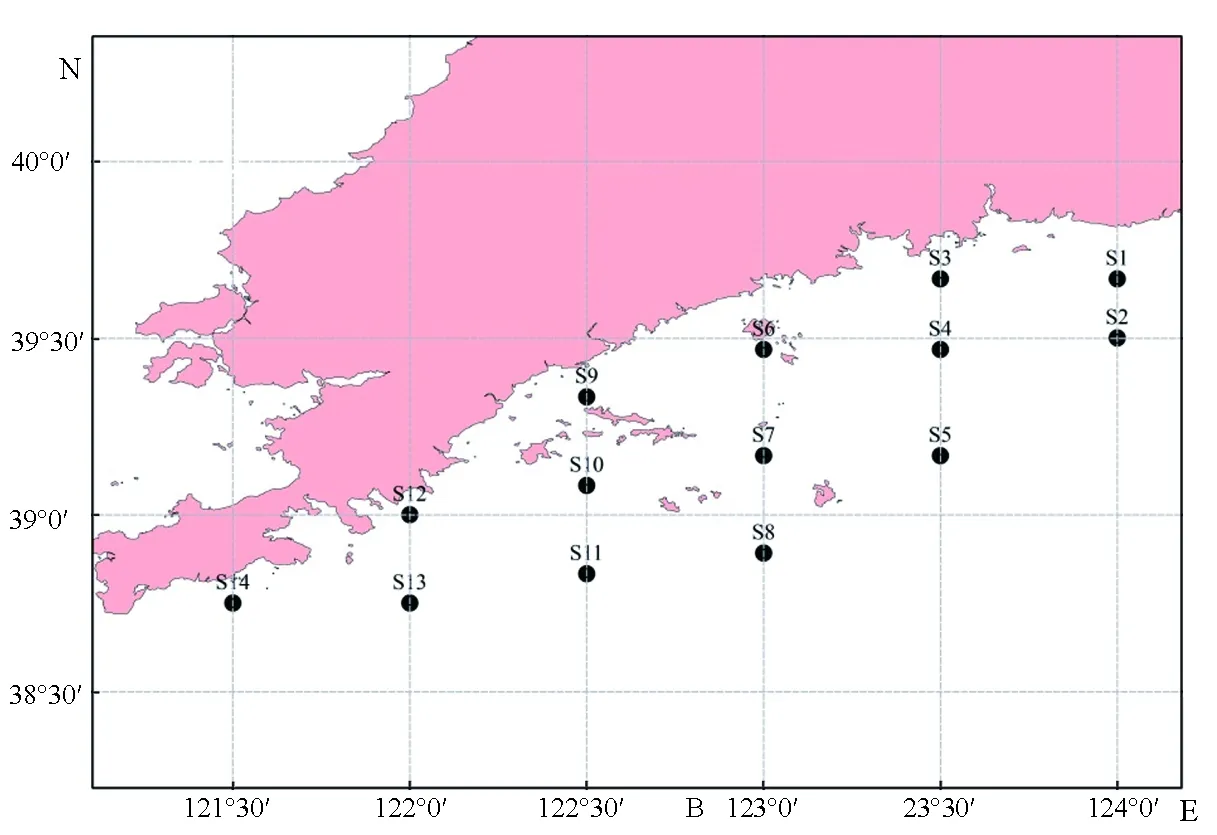
图1 调查站位图
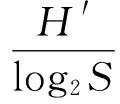
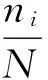
式中,N为某个样品中浮游动物的总个数;S为种类数;Pi为第i种的个体数在总个数中的比例,ni为第i种的个体数量,fi为第i种生物的出现频度,Y≥0.02的种类可视为优势种。
2 结果与分析
2.1 种类组成
本次调查共鉴定浮游动物8门9大类群47种。浮游动物群落中,桡足类为最优势类群,有20种,占浮游动物总种数的43.48%;浮游幼虫和水母类各8种,分别占总种类数的17.02%;原生动物共鉴定出6种,占12.77%;枝角类、介形类、被囊类、端足类和毛颚类各1种,分别各占2.13%。桡足类以拟哲水蚤属(Paracalanus)、胸刺水蚤属(Centropages)、纺锤水蚤属(Acartia)和长腹剑水蚤属(Oithona)为优势类群。浮游幼虫则包括桡足类幼体、棘皮动物幼体、软体动物幼体、环节动物幼体等(表1)。
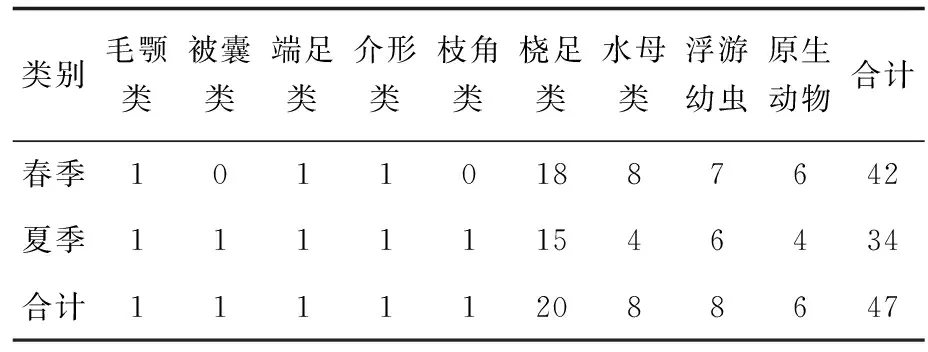
表1 春季和夏季浮游动物各类别种类数
从季节看,此次调查的浮游动物在春季出现42种,夏季34种。鉴定结果显示,两个季节的浮游动物均以浮游幼虫和桡足类为主,桡足类春季共18种,占春季总种类数的42.86%,浮游幼虫7种,占16.67%;夏季共鉴定出桡足类15种,占夏季总种类数的44.12%,浮游幼虫6种,占17.65%。从季节变化来看,黄海北部海域春季浮游动物种类数多于夏季。
2.2 丰度变化
春季浮游动物的丰度为1798.72~44 167.59个/m3,最低值出现在S5站位,最高值出现在S8站位,平均丰度为15 522.18个/m3。夏季浮游动物的丰度为425.32~32 314.81个/m3,最低值出现在S4站位,最高值出现在S14站位,平均丰度为7582.39个/m3(图2)。黄海北部海域调查站位的丰度变化较大。
从季节变化上看,辽宁省黄海北部沿岸春季浮游动物丰度远高于夏季,春季位于庄河和丹东交接的近岸海域,及大连湾口沿岸的南部海区的浮游动物丰度较低;夏季位于庄河和丹东交接的近岸海域及长海县近岸的浮游动物丰度较低。从平面分布上看,长海县附近及庄河和丹东交接的近岸海域是春夏季节浮游动物丰度的低值区。
2.3 生物量变化
浮游动物春季的生物量为111.46~1352.14 mg/m3,S4站位最高,S3站位最低,平均为512.09 mg/m3。夏季浮游动物的生物量为19.46~928.51 mg/m3,最高值出现在S14站位,最低值出现在S4站位,夏季平均生物量为218.84 mg/m3(图3)。黄海北部海域浮游动物生物量的变化趋势与丰度的变化趋势一致,从季节上看春季生物量高于夏季;从平面变化上看,长海县附近海域浮游动物生物量在春夏季节均偏低。
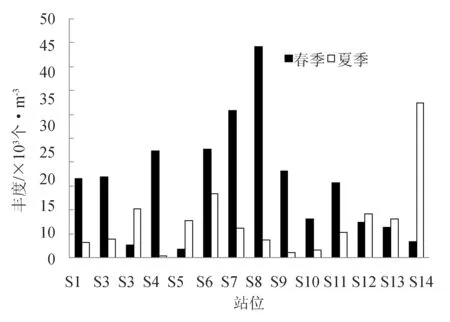
图2 春季和夏季各站位丰度
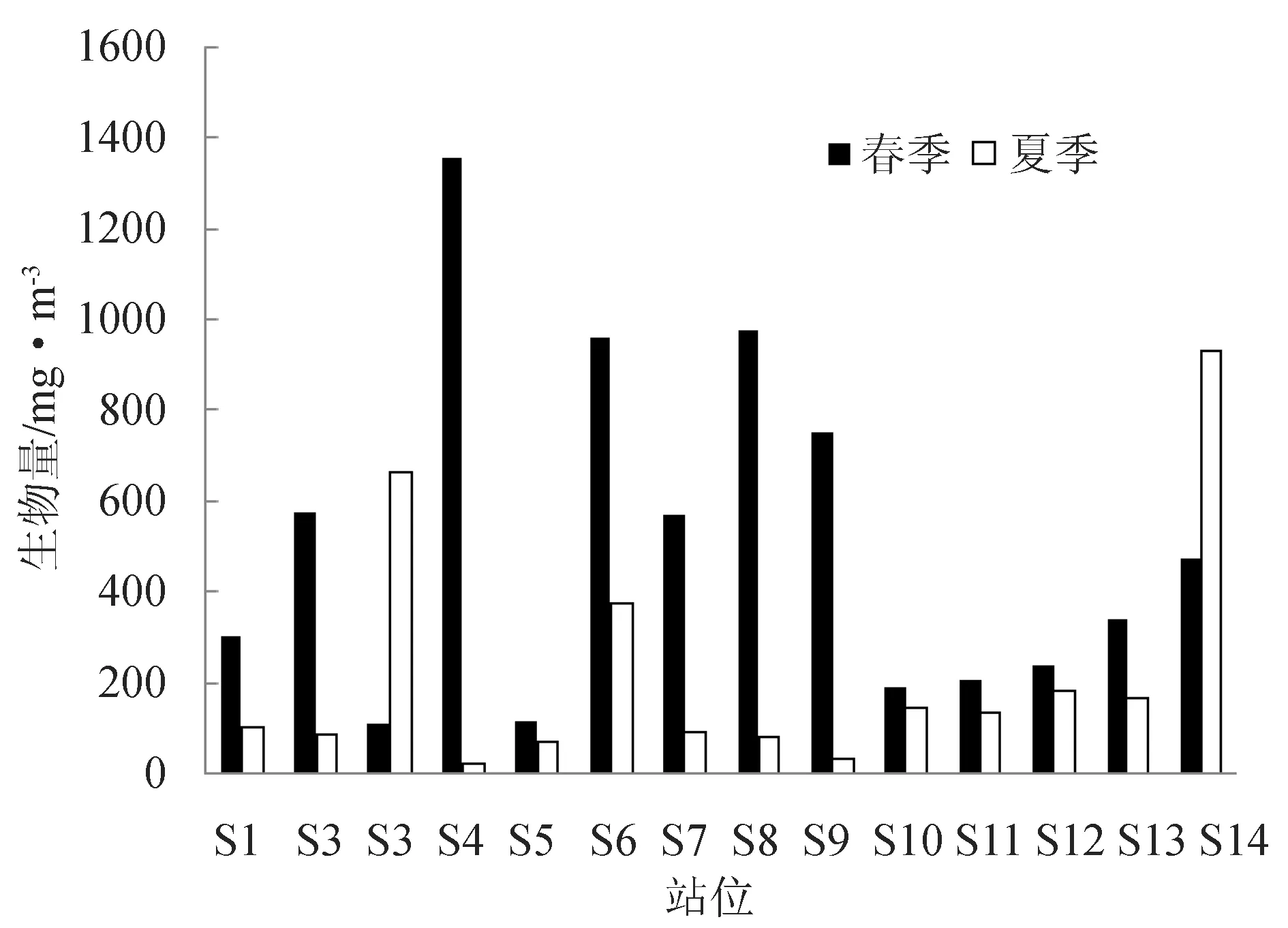
图3 春季和夏季各站位生物量
2.4 浮游动物优势种及季节变化
春季和夏季浮游动物的优势种类均为桡足类和浮游幼虫,占春夏浮游动物平均丰度的97.68%,箭虫类,水母类等数量较少。从季节上看,浮游幼虫在春季的丰度比例较大为62.04%,桡足类占35.02%,其他类占2.85%,箭虫类占0.09%。夏季,浮游幼虫丰度比例下降(29.51%),桡足类的丰度比例上升(68.79%),其他类占1.08%,箭虫类占0.61%(图4)。其他类在春季主要由原生动物、水母类、介形类和端足类组成;夏季主要由枝角类、水母类、端足类、原生动物、介形类和被囊类组成。
黄海北部春季浮游动物优势种依次为桡足类幼虫、棘皮动物幼体、桡足类幼体、洪氏纺锤水蚤(A.hongi)、短角长腹剑水蚤(O.brevicornis)、小拟哲水蚤(P.parvus)、拟长腹剑水蚤(O.similis)、和腹针胸刺水蚤(C.abdominalis);夏季浮游动物优势种依次为桡足类幼体、洪氏纺锤水蚤,拟长腹剑水蚤、短角长腹剑水蚤、桡足类幼虫、小拟哲水蚤、和腹针胸刺水蚤(表2)。
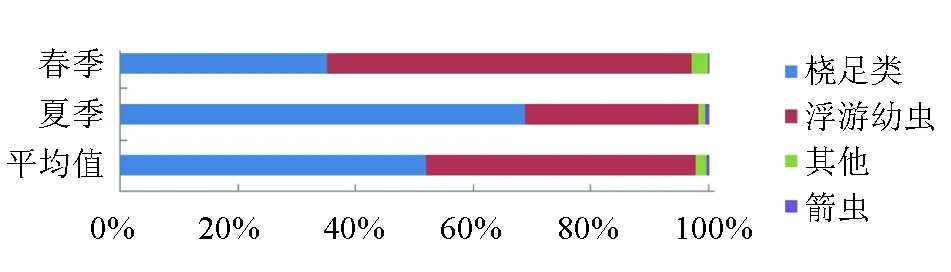
图4 各类群组成及季节变化
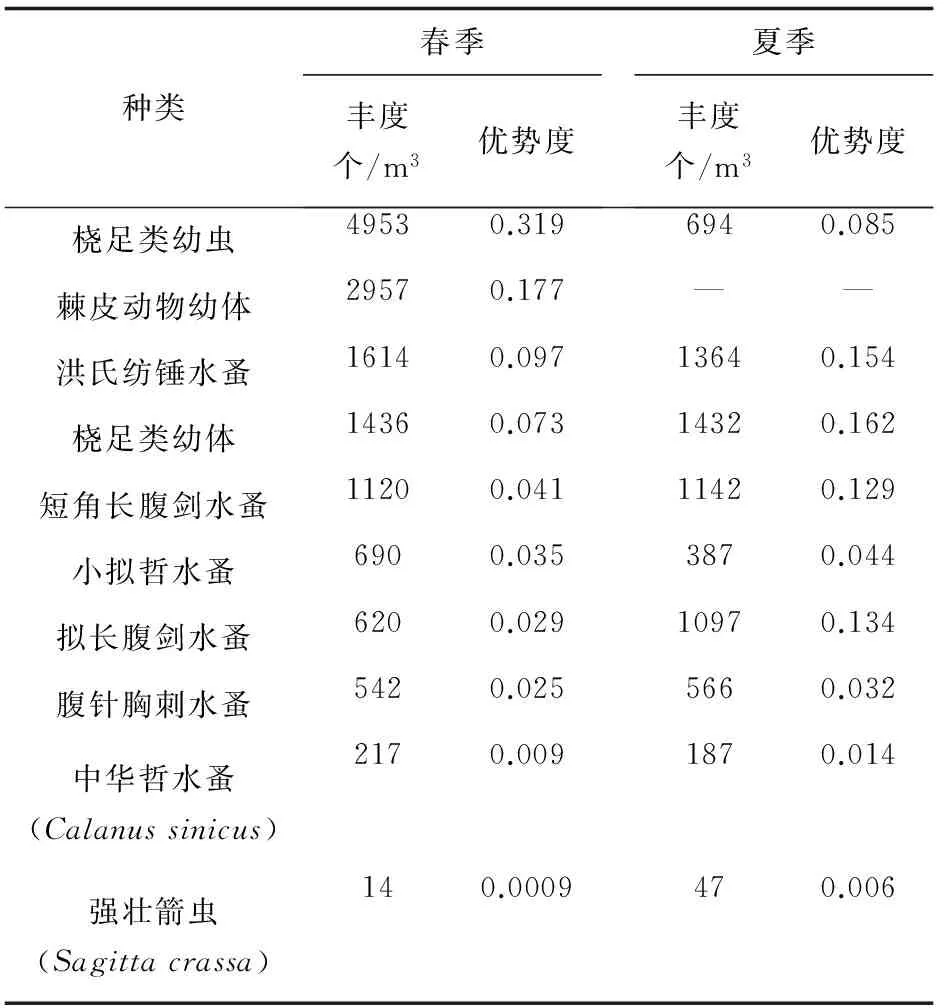
种类春季丰度个/m3优势度夏季丰度个/m3优势度桡足类幼虫49530.3196940.085棘皮动物幼体29570.177——洪氏纺锤水蚤16140.09713640.154桡足类幼体14360.07314320.162短角长腹剑水蚤11200.04111420.129小拟哲水蚤6900.0353870.044拟长腹剑水蚤6200.02910970.134腹针胸刺水蚤5420.0255660.032中华哲水蚤(Calanussinicus)2170.0091870.014强壮箭虫(Sagittacrassa)140.0009470.006
桡足类幼虫,春季丰度为769~10 000个/m3,平均丰度为4953个/m3,占浮游动物丰度的31.91%,为春季第一优势种;夏季0~1818个/m3,平均丰度为694个/m3。
棘皮动物幼体,春季丰度为0~12 222个/m3,平均丰度为2957个/m3,占浮游动物丰度的19.05%;在夏季未形成优势种类。
桡足类幼体,春季丰度为0~6250个/m3,平均丰度为1436个/m3;夏季0~7963个/m3,平均丰度为1431个/m3,占夏季浮游动物的丰度比例为18.88%,为夏季第一优势种。
洪氏纺锤水蚤在黄海沿岸均有分布。春季丰度为0~4861个/m3,平均丰度为1614个/m3;夏季0~5833个/m3,平均丰度为1364个/m3。
短角长腹剑水蚤是我国沿岸广泛分布的一种小型桡足类,尤其在近岸河口的数量尤为可观。在黄海北部沿岸,春季丰度为0~7820个/m3,平均丰度为1120个/m3,夏季0~4629个/m3,平均丰度为1142个/m3。
小拟哲水蚤春季丰度为0~2250个/m3,平均丰度为690个/m3,夏季0~1583个/m3,平均丰度为387个/m3。
拟长腹剑水蚤春季丰度为0~2500个/m3,平均丰度为620个/m3,夏季0~6417个/m3,平均丰度为1097个/m3。
腹针胸刺水蚤属于沿岸河口种类,一般栖息于港湾的表层。春季丰度为0~3333个/m3,平均丰度为542个/m3,夏季0~7314个/m3,平均丰度为566个/m3。
中华哲水蚤为暖温带外海种类,为大中性浮游动物的优势种。春季丰度为0~1667个/m3,平均丰度为216个/m3,夏季0~1944个/m3,平均丰度为187个/m3。
强壮箭虫为近岸低盐种,在黄海北部浮游动物生物组成中占重要地位。春季丰度为1~36个/m3,平均丰度为15个/m3,夏季0~333个/m3,平均丰度为47个/m3。
2.5 物种多样性,丰富度和均匀度
浮游动物多样性指数和均匀度指数均为夏季高于春季,但物种丰富度指数春季较高。春季浮游动物的多样性指数为1.53~2.99,平均值2.46,夏季浮游动物的多样性指数为0.97~3.04,平均值2.51;春季浮游动物丰富度指数为0.44~1.14,平均值0.76;夏季浮游动物丰富度指数为0.46~0.90,平均值0.72。春季物种均匀度指数为0.59~0.78,平均值0.71,夏季物种均匀度指数为0.31~0.96,平均值0.77(表3)。
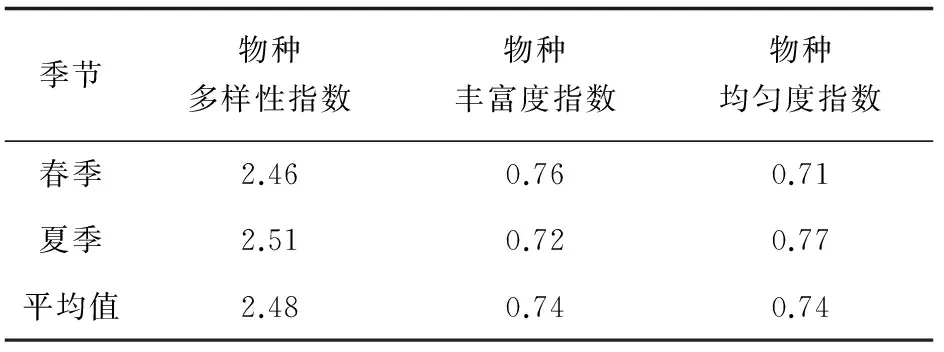
表3 黄海北部沿岸浮游动物多样性指数、丰富度指数、均匀度指数
3 讨 论
3.1 浮游动物数量变动及种类组成
浮游动物,特别是桡足类是海洋鱼类的基础饵料,作为生态系统中初级生产向次级生产转换的主要媒介,有关海洋浮游动物的调查研究对于渔业、养殖业发展具有重要的实践意义[11]。黄海北部海域为辽东半岛、山东半岛及朝鲜半岛之间呈半封闭状态的暖温带海域[3],因此,研究该海域浮游动物对环境变化的响应模式能够为气候变化的区域响应机制提供参考数据。
调查海域春季浮游动物总种类数多于夏季,桡足类、水母类、浮游幼虫及原生动物种类数均为春季较多。研究发现,氨氮含量与浮游动物种类数之间有极显著的负相关性[12],夏季水温的升高会使氨氮值上升,同时夏季沿岸人为干扰增加,生活污水,农业用水等氨氮含量较高的废水流入沿岸海域,人为作用和自然条件的共同影响可能引起夏季浮游动物种类数下降[12-13]。
辽宁省黄海北部沿岸海域受辽南沿岸流影响显著,不同季节数量变动差异较大。该海域浮游动物春季丰度及生物量均显著高于夏季。黄海北部海域重要水文特征之一是黄海暖流,其向黄海北部海域输送了大量高温高盐的外海水[14],同时也带来大量的营养物质,促进了浮游植物的大量繁殖,引起浮游动物数量增加。黄海暖流强弱变化呈季节性波动,秋季形成,夏季最弱且局限于北黄海的南端[15],夏季高温高盐外海水补充量的降低,有可能是春季浮游动物生物量和丰度高于夏季的原因之一;另一方面,温度作为影响动物生长繁殖的最关键因素,对桡足类的生活也存在显著的影响,春季为大部分生物繁殖的季节,在适合的温度范围内(6.5~20 ℃),温度越高,无节幼虫、桡足幼体的繁育速度越快,但当温度继续升高时,其发育速度反而减慢[11]。春季浮游幼虫丰度显著增加,约占浮游动物总丰度的50%以上,也可能是导致春季浮游动物生物量及丰度高于夏季的另一重要原因。同时,渤海及黄海海域从6月开始进入伏季休渔期,伏季休渔期间一些以浮游动物为食的游泳生物数量有所增加,使得浮游动物被捕食的压力增大,这也可能是导致浮游动物生物量及丰度在夏季降低的原因。
大量的研究资料显示,全球气候的变化对海洋浮游动物的丰度及生物量、多样性等的时空分布产生巨大的影响[3]。孙松等[16]研究发现,近20年来,胶州湾浮游动物生物量和丰度均呈现增加的趋势,尤以春季为最。对比黄海北部2009年调查数据[17]显示,2009年春季浮游动物平均丰度为6063.8个/m3,夏季平均丰度为3014.4个/m3,春夏季节浮游动物丰度均有升高,春季升高较明显,是2009年的2.6倍。浮游动物的显著增加可能与饵料生物增加及捕食压力降低有关。Kang等[18]调查发现,20世纪末,黄海中华哲水蚤丰度与鳀鱼(Engraulisjaponicus)数量呈现负相关,鳀鱼作为中华哲水蚤的捕食者,其资源量自20世纪末起开始降低[19],捕食者数量的减少能够促进浮游动物丰度的增加。同时,全球水温的不断上升[20],浮游植物等饵料生物增加,促进了暖温性浮游种类丰度的增加和暖水性种类分布范围的北移[18]。因此,浮游动物丰度及生物量的长期变化与气候及人类活动密切相关。近10年来,黄海北部浮游动物生物量和丰度都呈现增加的趋势,同时胶质类浮游动物的数量亦有所增加,特别是春季水母数量增加显著。在全球范围内,水母类无论从数量还是种类上都出现了上升趋势,全球海洋胶质化现象明显[14,21],胶质水母类数量和种类的变化与海洋环境条件的改变密切相关[22-24]。
3.2 优势种及群落结构
本次调查结果显示,黄海北部海域浮游动物以桡足类和浮游幼虫为主,分别占浮游动物总丰度的51.9%和47.8%。春夏季优势类群季节变换明显。春季是桡足类及其他海洋生物开始繁殖的季节,因此,海区内的桡足类幼虫及其他浮游幼虫数量增加,桡足类幼虫成为海区的第一优势种,浮游幼虫丰度占夏季总丰度的62%。夏季是浮游动物迅速生长的季节,水温升高,浮游植物大量繁殖,其数量直接影响浮游动物的丰度,海区内桡足类幼体成为第一优势种,同时桡足类成体数量不断增多,洪氏纺锤水蚤成为海区第二优势种,桡足类丰度占夏季总丰度的69%,远超其他种类。浮游动物的种类组成具有明显的季节性变化特征。
强壮箭虫和中华哲水蚤是黄海北部海域大中型浮游动物的优势种。中华哲水蚤和强壮箭虫属于近岸性暖温种,在黄海北部海域春、夏季均为大中型浮游动物优势种。春季,中华哲水蚤平均丰度较夏季略有升高,但最高丰度略有下降,各站位波动幅度不大。温度被认为是限制中华哲水蚤分布的最主要原因[25],此次调查时间为3月和6月,温度较适宜中华哲水蚤生长,因此,春夏季丰度差别不大。而强壮箭虫丰度从春季到夏季持续升高,春季的平均丰度及最高丰度都显著低于夏季。研究发现,强壮箭虫可以适应较高的温度上限[26],其丰度的变化与环境因子的相关性不高,但受桡足类丰度影响显著[27]。强壮箭虫以桡足类为主要食物来源[28],桡足类丰度的增加,为强壮箭虫提供了丰富的饵料。因此夏季调查海区适宜的环境条件,充足的食物来源使得强壮箭虫丰度显著增加。
与其他环境因子相比,水温是影响浮游动物群落结构特征的重要因素之一, 在浮游动物适宜的温度范围内,其多样性及均匀度会随着水温的升高而增加。因此,辽宁沿岸黄海北部海域夏季物种均匀度及多样性高于春季,但是由于春季浮游幼体丰度的增加,使得春季物种丰富度高于夏季。研究发现,海域内具有一定程度的干扰频率,会增加物种多样性,干扰强度过低,竞争能力强的物种将成为优势种;但当干扰强度过高时,生存能力强的物种会成为优势种,因此,只有在中等强度的干扰条件下,物种的生存机会均等,此时,群落多样性较高[29]。本次调查为黄海北部辽宁近岸海域,同时又是多条河流的入海口,春夏季入海径流增加,因此该海域受人类活动和自然环境条件的影响显著,其群落结构及生态特性在各种因素的综合作用下,物种均匀度、生物多样性及物种丰富度等指标均较高,总体群落结构较成熟、稳定。
[1] 马璐, 曹文清, 张文静, 等. 北部湾北部海域夏季微型浮游动物对浮游植物的摄食压力[J]. 生态学报, 2014, 34(3):546-554.
[2] Calbet A,Landry M R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems[J]. Limnology and Oceanography, 2004, 49(1):51-57.
[3] 杨青, 王真良, 樊景凤, 等. 北黄海秋、冬季浮游动物多样性及年间变化[J]. 生态学报, 2012, 32(21): 6747-6754.
[4] 蓝先洪, 李日辉, 张志珣, 等. 渤海东部与黄海北部表层沉积物的元素地球化学记录[J]. 地球学报, 2015, 36(6):718-728.
[5] Liu J, Saito Y, Wang H, et al.Sedimentary evolution of the Holocene subaqueous clinoform off the Shandong Peninsula in the Yellow Sea[J]. Marine Geology, 2007, 236(3):165-187.
[6] 国家海洋局. GB 17378—2007,海洋监测规范[S].北京:中国标准出版社,2008.
[7] Shannon C E, Weaver W. The Mathematical Theory of Communication[M].Urbana I L:University of Illi-nois Press,1949:1-125.
[8] Margalef R. Information theory in ecology[J].Gen Syst,1958,3(1):36-71.
[9] Pielou E C. An Introduction to Mathematical Ecology[M]. New York:Wiley-Interscience,1969.
[10] 方良, 李纯厚, 杜飞雁, 等. 大亚湾海域浮游动物生态特征[J]. 生态学报, 2010, 30(11):2981-2991.
[11] 高敬. 海洋桡足类生态学研究进展[J]. 河南科技, 2015(11):85-86.
[12] 高原, 赖子尼, 李捷,等. 连江浮游动物多样性的空间分布[J]. 生物多样性, 2013, 21(6):699-708.
[13] 靳萍, 代克岩, 杨程,等. 牧野湖浮游动物群落结构及其与环境因子的相关性研究[J]. 水生态学杂志, 2013, 34(2):53-61.
[14] 杜明敏, 刘镇盛, 王春生,等. 中国近海浮游动物群落结构及季节变化[J]. 生态学报, 2013, 33(17):5407-5418.
[15] 鲍献文,李娜,姚志刚,等.北黄海温盐分布季节变化特征分析[J].中国海洋大学学报:自然科学版, 2009, 39(4):553-562.
[16] 孙松, 李超伦, 张光涛, 等. 胶州湾浮游动物群落长期变化[J]. 海洋与湖沼, 2011, 42(5):625-631.
[17] 王彬,于旭光,刘春洋,等.辽宁省黄海北部沿岸浮游动物的群落结构及季节变化[J].水产科学, 2009, 28(增刊):62-66.
[18] Kang J H, Kim W S,Jeong H J, et al. Why did the copepodCalanussinicusincrease during the 1990s in the Yellow Sea?[J]. Marine Environmental Research, 2007, 63(1):82-90.
[19] 赵宪勇.黄海鳀鱼种群动力学特征及其资源可持续利用[D].青岛:中国海洋大学,2006.
[20] Lin C, Ning X, Su J, et al. Environmental changes and the responses of the ecosystems of the Yellow Sea during 1976—2000[J]. Journal of Marine Systems, 2005, 55(3):223-234.
[21] 刘镇盛. 长江口及其邻近海域浮游动物群落结构和多样性研究[D]. 青岛:中国海洋大学,2012.
[22] Lynam C P, Hay S J, Brierley A S. Interannual variability in abundance of North Sea jellyfish and links to the North Atlantic Oscillation[J]. Limnology and Oceanography, 2004, 49(3):637-643.
[23] Brodeur R D, Mills C E, Overland J E, et al. Evidence for a substantial increase in gelatinous zooplankton in the Bering Sea, with possible links to climate change[J]. Fisheries Oceanography, 1999, 8(4): 296-306.
[24] Lindley J A, Batten S D. Long-term variability in the diversity of North Sea zooplankton[J]. Journal of the Marine Biological Association of the UK, 2002, 82(1):31-40.
[25] 王丽, 王保栋, 陈求稳, 等. 三峡三期蓄水后长江口海域浮游动物群落特征及影响因子[J]. 生态学报, 2015, 36(9):2505-2512.
[26] 刘青,曲晗,张硕.强壮箭虫对温度、盐度的耐受性研究[J].海洋湖沼通报,2007(1):111-116.
[27] 王晓, 姜美洁, 刘萍,等. 秋季南黄海浮游动物分布及其影响因素[J].海洋学报, 2016(10):125-134
[28] 杨纪明,李军.渤海强壮箭虫摄食的初步研究[J]. 海洋科学, 1995(6):38-42.
[29] Connell J H. Diversity in tropical rain forests and coral reefs[J]. Science, 1978, 199(4335):1302-1310.
SeasonalVariationandCommunityStructureofZooplanktoninNorthernYellowSeainLiaoninginSpringandSummer
DUAN Yan1, WANG Aiyong1, WANG Xiaolin1, LIU Xiuze1,CHAI Yu1, LU Yang1,2, WANG Bin1, DONG Jing1
( 1.Key Laboratory of Marine Biological Resources and Ecology, Liaoning Ocean and Fisheries Science Research Institute, Dalian 116023, China; 2.Dalian Ocean University, Dalian 116023, China )
The seasonal variation and community structure of zooplankton in northern Yellow Sea in Liaoning province was studied based on seasonal investigation data obtained in spring (March of 2015) and in summer (June of 2015). Forty-seven species were found including forty-two species in spring and thirty-four species in summer. The copepod and pelagic larvae were found to be the dominant groups. Nauplius, ophiopluteus larva, copepodite,Acartiahongi,Oithonabrevicornis,Paracalanusparvus,O.similes, andCentropagesabdominaliswere dominant species in the spring, and copepodite,Acartiahongi,O.similes,O.abrevicornis, nauplius,P.parvus, andC.abdominaliswere dominant species in the summer. The results showed that the biomass of zooplankton was 512.09 mg/m3and the abundance was 15 522.18 ind/m3in spring, and the biomass of zooplankton was 218.84 mg/m3and the abundance was 7582.39 ind/m3in summer. There was higher biomass and abundance of zooplankton in spring than those in summer, indicating that the seasonal change could affect the biomass and abundance of zooplankton. Copepods were the most dominant group in number of species and abundance in the community structure of zooplankton in northern Yellow Sea.
northern Yellow Sea; zooplankton; abundance;biomass;community structure; seasonal variation
10.16378/j.cnki.1003-1111.2017.04.009
S932.2
A
1003-1111(2017)04-0456-06
2016-11-08;
2016-12-05.
国家海洋公益性行业科研专项(201405010).
段妍(1984-),女,助理研究员,博士;研究方向:资源增殖学及鱼类行为学. E-mail: chongstar@163.com.通讯作者:董婧(1966-),女,研究员;研究方向:水母生态学及渔业资源.E-mail:dj660228@tom.com.
