浙江省无花果种质资源亲缘关系的SRAP分析评价
2017-12-13刘亚群王燕飞段柳会王丽玲张飞英韩素芳
刘亚群,王燕飞,段柳会,王丽玲,张飞英,韩素芳
浙江省无花果种质资源亲缘关系的SRAP分析评价
刘亚群1,王燕飞2,段柳会3,王丽玲1,张飞英1,韩素芳1
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 龙泉市林场,浙江 丽水 323700;3. 开化县音坑乡林业站,浙江 衢州 324309)
利用SRAP分子标记技术,结合物候期对从美国、英国、意大利、法国、中国等地收集的适合浙江省种植的15个无花果品种的亲缘关系进行分析。结果表明,不同品种物候期存在不同程度差异,萌动期均为3月中下旬;展叶期在4月中下旬;座果期基本为5月初;果实成熟期6月底、7月中旬、7月底,8月中旬、9月均有;筛选出SRAP引物34个,扩增出204条多态性条带,多态率为62.25%,表明无花果种质资源的种内变异较为丰富;15个品种间的遗传相似系数在0.691 ~ 0.922,其中‘波姬红’与‘玛斯义陶芬’,‘美丽亚’与‘金傲芬’之间的遗传相似系数最大,为0.922,亲缘关系最近;‘红矮生’和‘日本紫果’遗传相似系数最小,为0.691。聚类分析表明在遗传距离为0.735处,不同品种分为两个大类群,其中‘红矮生’单独聚为一类,其余品种聚为一类。物候期观察表明,同一生长环境下,物候期与品种亲缘关系相关。
无花果;种质资源;亲缘关系;SRAP;分子标记
无花果Ficus carica为桑科Moraceae榕属Ficus植物,其隐花果味甜可食,富含活性多糖、黄酮等功能性成分。早在《神农本草经》就记载其果实有健胃清肠、消食解毒功效,具有降血压、降血脂、抗氧化、增强免疫力等多种功能,在临床上可用于治疗糖尿病、咽喉肿痛、肿瘤及免疫功能低下等疾病,堪称“圣果”[1-2]。无花果对土壤要求不严,在砂土、微酸性及盐碱地均可种植,新疆、山东、江苏、浙江均有广泛栽培[3]。中国无花果品种有1 000多个,但具有推广价值的不超过100个[4]。无花果可分为野生类型和栽培类型,后者又根据结实是否需授粉分为普通类型、斯密尔那类型、中间类型和原生类型,目前世界范围内栽培品种多为普通类型无花果,不经授粉即可形成可食用的果实。根据果实成熟时期将无花果分为夏果专用种、秋果专用种、夏秋果兼用种。根据无花果果皮和果肉颜色可分为绿色品种,如‘青皮’、‘绿抗 l号’;红色品种,如‘波姬红’、‘日本紫果’、‘玛斯义陶芬’等;黄色品种,如‘丰产黄’、‘布兰瑞克’、‘金傲芬’等。但上述标准过于简单,难以区分数以百计的品种资源[5]。2003年国际植物遗传资源研究所和地中海高级农业国际研究中心共同编辑《Descriptors for Fig(Ficus carica L.)》,对无花果表型性状有了更细致的描述,但由于描述的性状数量不多,在应用上仍有一定的局限性[6]。浙江省金华、嘉兴、湖州等地都有规模种植,主栽品种有‘玛斯义陶芬’、‘绿抗1号’等十几个品种。无花果多用扦插繁殖,2015年本研究组采用扦插方式建立了种质资源保存圃,并对不同无花果品种大田扦插的生长特性进行了研究[7]。随着经济增长和人民生活水平的提高,无花果的营养、药用和保健价值日益受到重视,社会需求越来越多,浙江省栽培面积也逐渐扩大。不同环境条件下的繁殖栽培,也会造成种内变异的多样性[8],因此建立在DNA水平之上的分子标记为无花果遗传图谱构建、基因定位、亲缘关系分析、品种分类鉴定及种质资源保护等开辟了新途径。
相关序列扩增多态性(Sequence-related amplified polymorphism,SRAP)技术具有简便、稳定、中等产率、高共显性、便于克隆测序目标片段、在基因组中分布均匀等特点,已广泛应用于樱桃Cerasus pseudocerasus,梅Armeniaca mume,苹果 Malus pumila,柑橘 Citrusreticulata,阳芋 Solanum tuberosum,稻 Oryza sativa,芸苔 Brassica campestris,蒜Allium sativum,莴苣Lactuca sativa,旱芹Apium gravelens,草棉Gossypium herbaceum等植物和稻瘟病(病原菌:Magnaporthe grisea)的遗传图谱构建比较基因组学、遗传多样性分析、基因定位、杂种优势预测等方面的研究[9-14]。本文利用SRAP标记技术结合物候期对适合浙江省栽培的15份无花果种质资源材料进行了亲缘关系分析评价,以期为无花果种质资源的保存、保护及分子辅助选育提供理论基础。
1 材料与方法
1.1 种质资源圃概况
无花果种质资源圃位于浙江省武义县桑和水果专业合作社,119°49′ E,28°54′ N,海拔91 m,酸性红壤,中亚热带季风气候,年平均气温16.9℃,年降水量1 445.7 mm,年日照时数1 963.7 h。该圃建于2015年,占地0.13 hm2,15个种质引自浙江嘉兴神农无花果果园,每份种质资源25株,为1年生裸根苗,平均高55 cm,地径1 cm。按行株距2.5 m×1.0 m分行定植。种源原产地分别为美国、意大利、英国、西班牙、中国,详见表1。
1.2 材料
2016年3月,每个品种随机选取5株,每株采集植株顶端初展开的完全叶1 ~ 2片,同一品种的嫩叶合并放入装有硅胶干燥剂的密封袋中,带回实验室备用。
1.3 试验方法
1.3.1 基因组DNA的提取 基因组提取采用新型快速植物基因组DNA提取试剂盒(离心柱型)(BioTeke,北京),DNA提取后通过1.5%琼脂糖凝胶电泳和Nano Drop 2000C微量核酸蛋白测定仪检测完整性、纯度及浓度。D260/D280在1.8 ~ 2.0之间的DNA样品用于后续PCR扩增。
1.3.2 SRAP引物筛选和PCR扩增 从15份样品中随机选择5份样品进行SRAP引物筛选,共获得34对扩增条带稳定、清晰的多态性引物(见表2)。利用这34条引物对15份无花果样品进行SRAP-PCR扩增。SRAP扩增反应采用20 μL体系:10.0 μL 2×Power Taq PCR MasterMix,模板DNA 60 ng,SRAP上游引物1 μL (10 μmol·L-1),下游引物 1 μL(10μmol·L-1),ddH2O 补足 20 μL。PCR 扩增反应在 TC-XP 型扩增仪(杭州博日)上进行。SRAP-PCR扩增程序为94℃预变性4 min;94℃变性1 min,35℃退火30 s,72℃延伸1.5 min,5个循环,94℃变性1 min,50℃退火30 s,72℃延伸1.5 min,35个循环,循环结束后72℃延伸7 min,4℃保存。反应结束后,将PCR扩增产物用1.5%琼脂糖凝胶电泳,检测PCR扩增片段的数量、片段分子量大小以及多态性。

表1 15 个无花果品种特征信息[3,15]Table 1 Information of 15 cultivarsof Ficus carica

表2 SRAP引物Table 2 SRAP primers
1.3.3 数据处理与统计分析 PCR扩增产物用琼脂糖凝胶电泳分析后,根据实验结果,分析记录同一水平位置重复出现的条带。在相同的迁移率上,有带用1表示,无带用0表示。采用NTSYS-pc2.10e软件进行遗传多样性分析,使用软件中的SimQual程序计算样品间的DICE遗传相似系数(GS,Genetic similarity)和遗传距离(GD,Genetic distance),用SHAN程序进行非加权成对算术平均法聚类分析(UPGMA,Unweighted pair-group method with arithmetic means),通过Tree plot模块生成聚类图。
1.4 物候观测
2016年2-10月,每个品种随机选取5株进行萌动期、展叶期、座果期及果实成熟期观测,每3 d记录1次,取平均值作为结果。
2 结果与分析
2.1 物候期观测分析
对无花果种质资源圃15个品种萌动期等物候期进行观测,结果见表3。由表3可知,所有品种的萌动期均在3月中下旬。除‘B1011’,‘日本紫果’,‘B110’及‘青皮’,其余都在4月中下旬进入展叶期。除‘日本紫果’在6月底,‘AA1213’,‘红矮生’在5月下旬,其其余座果期均均为5月初。果果实成熟期则则差别较大,分为6月底、7月中旬、7月底,8月中旬、9月初初五类。从物候候期上看,品品种间存在不不同程度的差异异。其中‘金金傲芬’和‘美美丽亚’除萌萌动期相差155 d,其余均比较接近;‘波姬红’及及‘玛斯义陶陶芬’的座果期一致,其余余差别都在115 d以内。从从形态上看,‘波姬红’和和‘玛斯义陶陶芬’果实颜颜色相近,叶片裂度也相似似,‘金傲芬芬’和‘美丽丽亚’果实颜色(图 1)及及叶片特征也也均十分相似似,需通过基因组学来判别别。
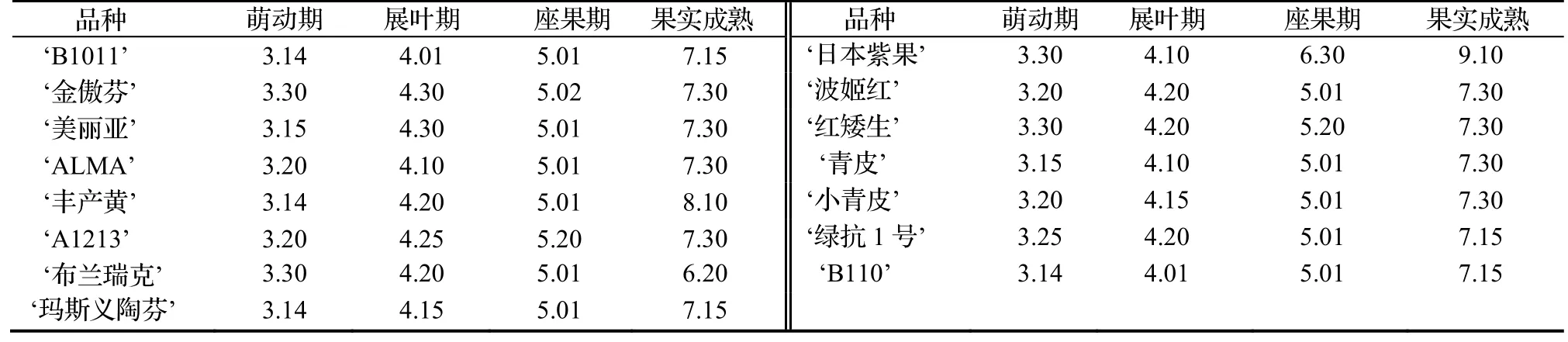
表3 15个无花果种质的物候期观测结果Table 3 Phelological period of 15 cultivars of F. carica
2.2 扩增产物的多态性分析
利用筛选选出的34个SSRAP引物对对无花果基因组DNA进行行PCR扩增,琼脂糖凝胶胶电泳结果显示均能扩增出清晰稳定、重复性和多多态性较高的条带。图2是引物MEE1 ~ EM8 对15个无花果品种的扩增增图谱。34条条引物扩增共共获得204条条带,条带带介于 100 ~2 000 bp,其其中多态性条带127条,多态率62.225%,表明无无花果种质间的基因差异异明显。

图1 ‘玛斯义陶芬’、‘‘波姬红’、‘金傲芬’、‘美丽亚’叶片片图Figure 11 Leaf of ‘Masuu i Dauphine’, ‘A1132’, ‘A212’ andd ‘A134’

图2 引物物 ME1 ~ EM88扩增图谱Figure 2 Amplification profile of primer ME1-EM8
2.3 遗传相似性分析
将34条条引物扩增出的条带作为原始矩阵,利利用NTSYS-pc软件计算15个品种间的的Dice遗传相似系数,得到遗传相似系数在0.691 ~ 0.922,平均为0.409,说明各个无花果品种间亲缘关系都比较近。其中‘波姬红’与‘玛斯义陶芬’、‘美丽亚’与‘金傲芬’之间的遗传相似系数最大为0.922,表明两者间的亲缘关系最近,与观测到的品种形态特征结果一致;其次为‘青皮’与‘绿抗1号’,其遗传相似系数为0.902。亲缘关系最远的为‘红矮生’和‘日本紫果’,其遗传相似系数最小为0.691;其次为‘日本紫果’与‘B110’,其遗传相似系数为0.701。15个无花果品种间的遗传相似系数详见表4。
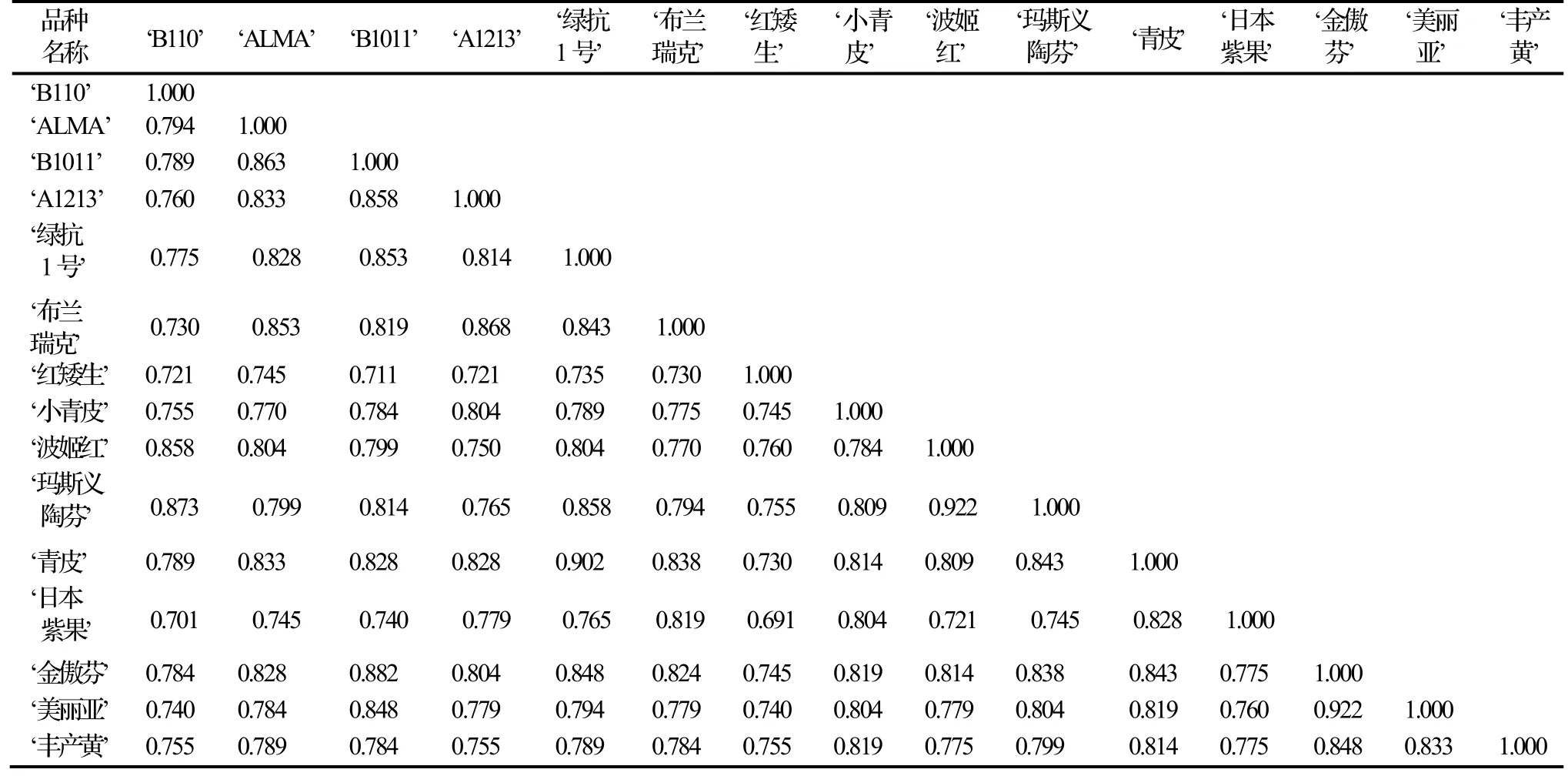
表4 15个无花果品种间遗传相似系数Table 4 Genetic similarity coefficient among 15 F. carica cultivars
2.4 聚类分析
利用UPGMA法对15个无花果品种进行聚类分析,建立聚类分支树状图(图3)。从图3可知,在遗传相似系数为0.735处可将15个无花果品种分为2大类。其中‘红矮生’单独聚为一类,其余品种聚为一类。表明‘红矮生’与其他品种之间的遗传差异较大,即亲缘关系较远。第 I大类可分为 3个小类。第一小类包含‘B110’、‘波姬红’及‘玛斯义陶芬’,且‘波姬红’及‘玛斯义陶芬’在最开始即相聚,表明两者之间亲缘关系较近。此外,‘小青皮’与‘日本紫果’虽然聚为一类,但是相聚时遗传相似系数为 0.8左右,表示两者之间还是存在一定的遗传差异。其余品种聚为第三类。总体而言,15个品种间的遗传差异性不大。

图3 15个无花果品种UPGMA聚类图Figure 3 UPGMA dendrogram of 15 cultivars of F. carica
3 结论与讨论
物种的遗传多样性是其长期进化的结果,也是其生存、适应和发展的前提,遗传多样性在物种形态、分子水平上均有所表现,丰富的遗传多样性是宝贵的基因资源。本研究利用SRAP分子标记技术,对采自浙江武义的15个无花果品种进行聚类分析,结合物候及果实性状比较不同无花果品种之间的遗传差异性。结果表明,利用筛选出的34条SRAP引物对所有样品进行PCR扩增,共获得204条条带,其中多态性条带127条,多态率为62.25%,且扩增条带清晰,样品多态性较高。
遗传相似性系数是用来比较群体或个体间相似程度的度量参数,平均相似系数越高,说明样品之间的差异性越小,即亲缘关系越近。本实验结果表明,15个样品之间的遗传相似系数变化范围在0.691 ~ 0.922。相似系数最高的是‘金傲芬’和‘美丽亚’、‘波姬红’和‘玛斯义陶芬’,为0.922;最低的是‘红矮生’与‘日本紫果’,为0.691。‘红矮生’为盆栽无花果专用品种,小灌木,树冠矮小,枝条节间短,分枝多,叶中大,掌状五裂;果紫红色,从形态上与其他无花果品种有显著差别,与遗传相似系数最低符合,总体而言15个样品间的遗传差异性不大。
进一步的聚类分析结果表明,‘金傲芬’与‘美丽亚’、‘青皮’和‘绿抗1号’、‘波姬红’及‘玛斯义陶芬’三类品种亲缘关系较近。其中‘金傲芬’与‘美丽亚’原产地均为美国加利福利亚州,都为黄色系品种;而‘青皮’和‘绿抗1号’产自中国的山东和江苏,此聚类结果将两者聚在一起,与样品的原产地的地理位置接近相符合。一般而言,在同一地区采集的样品遗传相似度较高[8]。‘青皮’和‘绿抗 1号’是极易混淆的品种,SRAP分子标记技术发现二者遗传相似系数接近,为下一步品种鉴别奠定基础。‘波姬红’及‘玛斯义陶芬’最初就聚集在一起,‘波姬红’原产地为美国德克萨斯州,而‘玛斯义陶芬’原产地为美国加利福尼亚州,地理位置接近;根据形态学观察,‘波姬红’果形为长卵形,果肉为浅红,果皮为紫红;‘玛斯义陶芬’果形为倒圆锥形,果肉为桃红色,成熟时果皮为紫褐色;从形态学和地理分布而言,两个无花果品种遗传差异性非常接近,遗传相似系数达到0.922。
物候期观察结果表明,同一生长条件下亲缘关系最近的‘金傲芬’与‘美丽亚’除萌动期相差15 d,展叶期、座果期及果实成熟期表现一致;而‘波姬红’及‘玛斯义陶芬’的座果期一致,其余差别亦在15 d以内。亲缘关系最远的‘红矮生’与‘日本紫果’除萌动期一致,展叶期呈现10 d的差别,座果期及果实成熟期相差40 d,差别较大。亲缘关系较远的‘日本紫果’和‘B110’萌动期等物候也表现出不同程度的差别,差别最大的为相差60 d座果期。同一生长环境下,物候期与品种类别呈现相关性,因此可考虑结合遗传多样性及更详尽的物候分析,判断品种亲缘关系远近,为无花果优良种源选育和推广应用提供一定理论基础。
[1] 浙江植物志编辑委员会. 浙江植物志(第二卷)[M]. 杭州:浙江科学技术出版社,1992.
[2] 马会勤. 从无花果产业看中国特色农业发展之路[J]. 中国农村科技,2015(2):32-33.
[3] 曹尚银. 无花果无公害高效栽培[M]. 北京:金盾出版社,2003.
[4] 姜卫兵. 无花果主要品种介绍[J]. 山西果树,1990,(4):27-32.
[5] 吴子江,马翠兰,郭阳彬,等. 无花果生产与研究进展[J]. 亚热带农业研究,2013,9(3):151-157.
[6] CABRITA L F,AKSOY U,HEPAKSOY S,et al.Suitability o fisozyme,RAPD and AFLP markersto assess genetic differences and relatedness among fig(Ficus carica L.)clones[J]. Sci Hor,2001(87):261-273.
[7] 刘亚群,韩素芳,张飞英,等. 不同无花果品种大田扦插的生长特性研究[J]. 浙江林业科技,2016,36(6):8-13.
[8] 王亮,王彩虹,田义轲,等. 山东省无花果种质资源多样性的RAPD分析[J]. 植物遗传资源学报,2007,8(3):303-307.
[9] LI G,QUIROS C F.Sequence-related amplified polymorphism(SRAP),a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica[J]. Theor Appl Gen,2001(103):455-461.
[10] 林忠旭,张献龙,聂以春,等. 棉花SRAP遗传连锁图构建[J]. 科学通报,2003,48(15):1676,1679.
[11] LI G,GAO M,YANG B,et al. Gene for gene alignment between the Brassica and Arabidopsis genomes by direct transcriptome mapping[J].Theor Appl Gen,2003,107(1):168-180.
[12] FERRIOL M,PICO B,NUEZ F. Genetic diversity of a germplasm collection of Cucurbitapepo using SRAP and AFLP markers [J]. Theor Appl Gen,2003,107(2):271-282.
[13] RIAZ A,LI G,QURESH Z,et al. Genetic diversity of oilseed Brassica napus inbred lines based on sequence-related amplified polymorphism and its relation to hybrid performance[J]. Plant Breed,2001,120(5):411-415.
[14] 吴伟怀,王玲,程贯忠,等. 稻瘟病菌群体的分子遗传学研究—广东省与云南省稻瘟病菌群体遗传及致病型结构的时空变化分析[J]. 中国农业科学,2004,37(10):1468-1473.
SRAP Analysis on Genetic Relationship of Germplasm Resources of Ficus carica in Zhejiang Province
LIU Ya-qun1,WANG Yan-fei2,DUAN Liu-hui3, WANG Li-ling1,ZHANG Fei-ying1,HAN Su-fang1
(1. Zhejiang Academy of Forestry, Hangzhou 310023, China; 2. Longquan Forest Farm of Zhejiang, Lishui 323700, China;3. Forestry Station of Yinkeng Town of Kaihua county, Quzhou, Zhejiang, 324309)
Fifteen culitvars of Ficus carica from the United States, the United Kingdom, Italy, France and China were planted in Wuyi, Zhejiang province in 2015, leaves were collected in March 2016 for evaluation of genetic relationships among these cultivars by SRAP markers and observations on phenological periods. The results showed that there were differences in phenological period among 15 germplasm resources, with sprouting period from middle to late March, leaf expansion from middle to late April, fruit setting at early May, and fruit mature from late June to the early September. 34 SRAP primers were selected, 204 polymorphic bands were obtained by PCR with the polymorphic rate of 62.25%, indicating abundant intraspecific variation. The genetic similarity coefficient among 15 cultivars was 0.691-0.922, with the maximal coefficient of 0.922 of‘A132’ and ‘Masui Dauphine’, and the minimal of 0.691 of ‘Violette Solise’ and ‘Hongaisheng’. Cluster analysis showed that tested germplasm could be divided into two groups at genetic distance of 0.735. ‘Hongaisheng’ was clustered alone, the others were clustered into one group. Observation on phenological period of tested cultivars planted in the same location showed that their phenology had relation with their provenances..
Ficus carica; germplasm resources; genetic relationship; SRAP; molecular marker
S663.3
A
1001-3776(2016)05-0023-06
10.3969/j.issn.1001-3776.2017.05.004
2017-03-25 ;
2017-07-08
浙江省科研院所专项(2015F30002)
刘亚群,高级工程师,从事林业土壤、植物的分析检测工作;E-mail:liuyaqun2005@aliyun.com。通信作者:韩素芳,副研究员,从事经济林栽培研究;E-mail:hansufang2004@126.com。
