雷帕霉素对衰老细胞生物节律关键基因的调控作用观察
2017-12-12刘佐君刘宝华孟凡彪
刘佐君,刘宝华,孟凡彪
(深圳大学,广东深圳518060)
雷帕霉素对衰老细胞生物节律关键基因的调控作用观察
刘佐君,刘宝华,孟凡彪
(深圳大学,广东深圳518060)
目的观察雷帕霉素对衰老细胞生物节律关键基因的调控作用。方法通过传代诱导人皮肤成纤维细胞复制性衰老,获得衰老的人皮肤成纤维细胞。加入50%的马血清刺激诱导细胞同步化,将细胞分为观察组和对照组,分别加入100 mmol/L的雷帕霉素和DMSO培养24 h。采用荧光定量PCR法检测生物节律关键基因芳烃受体核转位蛋白3(BMAL1)、节律周期蛋白2(PER2)、蓝光受体蛋白1(CRY1)、孤儿核受体α(Rev-erbα)、孤核受体α(Rorα)mRNA表达,利用JTK-CYCLE软件分析各基因的节律周期,Western blotting法检测BMAL1、PER2、CRY1蛋白表达。结果观察组PER2、CRY1、Rev-erbα、Rorα mRNA表达高于对照组(P均<0.05)。对照组BMAL1、CRY1、Rev-erbα、Rorα基因的节律周期为24 h,PER2基因的节律周期为28 h。观察组BMAL1、CRY1、Rorα基因的节律周期为24 h,Rev-erbα基因的节律周期为27.5 h,PER2的节律周期为24 h。观察组CRY1蛋白表达高于对照组(P<0.05),PER2蛋白表达低于对照组(P<0.05),两组BMAL1 mRNA及蛋白表达比较无统计学差异。结论雷帕霉素可以提高衰老的人皮肤成纤维细胞生物节律关键基因BMAL1、PER2、CRY1、Rev-erbα、Rorα mRNA的表达,缩短PER2的节律周期,对紊乱的节律钟功能具有调控作用。
生物节律;雷帕霉素;生物节律关键基因;芳烃受体核转位蛋白3;节律周期蛋白2;蓝光受体蛋白1;孤儿核受体α;孤核受体α;哺乳动物雷帕霉素靶蛋白
生物节律是生物为了适应外界环境(如光照、温度以及营养物质等)而产生的内在的、具有节律性的生命活动。人体的许多生理活动都具有生物节律,包括体温、睡眠、激素分泌、代谢以及免疫系统等。生物节律紊乱不仅会导致一系列的代谢疾病如糖尿病、肥胖等,而且与衰老以及衰老相关的心血管疾病等的发生息息相关[1,2]。研究[3,4]显示,生物节律关键基因芳烃受体核转位蛋白3(BMAL1)和节律调控蛋白(CLOCK)缺失会导致小鼠寿命缩短,并且伴随出现早衰的相关表型。人体的生物节律由两个主要的反馈环调节,其中一个反馈环是由BMAL1和CLOCK形成的异源二聚体激活下游的节律周期蛋白(PERs)和蓝光受体蛋白(CRYs)基因家族,PERs和CRYs基因反馈抑制BMAL1的转录活性,形成一个负反馈调控环;BMAL1还可以激活孤儿核受体(Rev-erb)和孤核受体(Ror)基因家族,Rev-erb和Ror基因反馈抑制BMAL1的转录活性,形成另外一个反馈环。哺乳动物雷帕霉素靶蛋白(mTOR)是一类进化保守的丝氨酸/苏氨酸类的蛋白激酶,与寿命有关,可以直接调控生物节律。抑制mTOR信号通路可以延长酵母[5]、线虫[6]、果蝇[7]、老鼠的寿命[8]。2015年1月~2017年1月,我们利用mTOR抑制剂雷帕霉素处理衰老的人皮肤成纤维细胞,检测细胞内生物节律关键基因BMAL1、节律周期蛋白2(PER2)、CRY1、Rev-erbα、Rorα的表达变化,探讨雷帕霉素对衰老细胞生物节律的调控作用。
1 材料与方法
1.1 材料 人皮肤成纤维细胞由本实验室冻存。胎牛血清(FBS)购自PAN-Biotech公司,雷帕霉素、巴佛洛霉素A1和氯喹购自Sigma公司,TRIzol和马血清购自Invitrogen公司,逆转录试剂盒和荧光定量试剂盒购自大连宝生物公司,PER2、CRY1、BMAL1抗体购自Abcam公司,细胞β-Galactosidase染色试剂盒购自Cell Signaling Technology公司。
1.2 细胞培养与分组处理 向人皮肤成纤维细胞中加入含有10% FBS、100 μg/mL青霉素、100 μg/mL链霉素的DMEM培养基。通过传代(从第16代培养到第30代)诱导细胞的复制性衰老,并通过β-gal染色确定细胞衰老。β-gal染色阳性(呈蓝色)细胞为衰老细胞。经测定,第30代细胞β-gal染色阳性细胞比例为60%±6%,符合实验要求。细胞生长到100%的细胞密度后,加入50%的马血清处理2 h诱导细胞同步化。将细胞分为两组,观察组加入100 mmol/L雷帕霉素,对照组加入DMSO。培养24 h后开始取样,每隔4 h取样1次,共取6次。
1.3 细胞内BMAL1、PER2、CRY1、Rev-erbα、Rorα mRNA表达检测 采用荧光定量PCR法。马血清处理24 h后开始收集样品,分别收集24、28、32、36、40、44 h的细胞样品。向细胞中加入TRIzol裂解液,提取mRNA,逆转录合成cDNA。引物序列:BMAL1上游引物5′-CATTAAGAGGTGCCACCAATCC-3′,下游引物5′-TCATTCTGGCTGTAGTTGAGGA-3′;CRY1上游引物5′-CTCCTCCAATGTGGGCATCAA-3′,CRY1下游引物5′-CCACGAATCACAAACAGACGG-3′;PER2上游引物5′-CTTCAGCGATGCCAAGTTTGT-3′,PER2下游引物5′-CGGATTTCATTCTCGTGGCTTT-3′;Rev-erbα上游引物5′-TCATGCTTGCGAAGGCTGTAA-3′,Rev-erbα下游引物5′-CGCTTAGGAATACGACCAAACC-3′;Rorα上游引物5′-TGCGAGGAACAATAGACCCAA-3′,Rorα下游引物5′-ATGGCCTATGTGTGCGTTGTA-3′。PCR反应条件:95 ℃ 2 min;60 ℃ 15 s;72 ℃ 20 s;72 ℃ 5 min。荧光定量PCR通过BIO-RAD CFX ConnectTMReal-time PCR仪器进行检测和分析。读取基因扩增的Ct值,以2-ΔΔCt法计算基因的相对表达量。利用JTK-CYCLE软件分析各基因的相对震荡幅度和节律周期[13]。
1.4 细胞内PER2、CRY1、BMAL1蛋白表达检测 采用Western blotting法。马血清处理24 h后开始收集样品,分别收集24、28、32、36、40、44 h的细胞样品。向细胞中加入SDS上样缓冲液裂解,煮沸10 min。经过SDS-PAGE电泳转膜到含有正电荷的PVDF膜。经过TBST含有5%的脱脂奶粉封闭1 h,加入一抗4 ℃过夜,TBST冲洗,加入羊抗兔的二抗孵育1 h,TBST冲洗,化学发光显影。利用Image J软件对蛋白条带进行半定量分析,以目标蛋白与β-actin灰度值的比值表示目标蛋白的相对表达量。
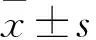
2 结果
2.1 两组BMAL1、PER2、CRY1、Rev-erbα、Rorα mRNA的相对震荡幅度比较 观察组BMAL1 mRNA的相对震荡幅度与对照组比较无统计学差异,PER2、CRY1、Rev-erbα、Rorα mRNA的相对震荡幅度较对照组升高(P<0.05或<0.01)。见表1。

表1 两组BMAL1、PER2、CRY1、Rev-erbα、Rorα mRNA相对震荡幅度比较
注:与对照组比较,*P<0.05,**P<0.01。
2.2 两组BMAL1、PER2、CRY1、Rev-erbα、Rorα mRNA的节律周期比较 对照组BMAL1、CRY1、Rev-erbα、Rorα mRNA的节律周期为24 h,PER2 mRNA的节律周期为28 h;观察组BMAL1、CRY1、Rorα mRNA的节律周期为24 h,Rev-erbα mRNA的节律周期为27.5 h,PER2 mRNA的节律周期缩短到24 h。
2.3 两组BMAL1、PER2、CRY1蛋白表达水平比较 观察组PER2蛋白表达低于对照组,CRY1蛋白表达高于对照组,BMAL1蛋白表达无明显变化。见表2。
表2 两组BMAL1、PER2、CRY1蛋白表达水平比较
注:与对照组比较,*P<0.05,**P<0.01。
3 讨论
生物节律控制着人体的睡眠、觉醒、进食等生理活动。生物节律紊乱会导致多种疾病的产生。机体衰老伴随着生物节律功能的失调,最明显的就是老年人的睡眠质量和睡眠规律性下降。Chang等[9]研究认为,衰老导致生物节律功能失调的分子机制可能是由生物节律关键基因BMAL1、PER2、CRY1、Rev-erbα、Rorα表达水平以及震荡幅度降低所致。研究显示,生物节律关键基因BMAL1、CLOCK突变会导致小鼠出现早衰的表型[3,4]。因此,上调生物节律基因表达可能有助于提高生物节律基因的震荡幅度,进一步恢复老年人的生物节律功能。研究[10~13]显示,mTOR与生物节律关键基因BMAL1有相互调控的关系,mTOR下游的蛋白质激酶S6K1磷酸化BMAL1,BAML1的磷酸化修饰进一步调控真核细胞的翻译过程。抑制mTOR信号通路可以显著延长酵母、线虫、果蝇、老鼠的寿命,但是否可以影响人衰老细胞的生物节律尚无定论。
雷帕霉素是mTOR的抑制剂,虽然mTOR信号通路调控生物节律的研究已经有相关的报道,但是直接利用雷帕霉素处理衰老细胞以及在动物体内的研究还鲜有报道。节律基因的震荡幅度是研究生物节律的一个重要指标,其主要表示一个节律周期内基因的最高表达量与最低表达量的相对比值。节律钟关键基因BMAL1和CLOCK突变都会导致节律基因的震荡幅度的丧失[3,4]。而衰老老鼠的节律钟基因的震荡幅度是降低的[9]。本研究通过传代获得衰老的人皮肤成纤维细胞,加入mTOR抑制剂雷帕霉素,发现雷帕霉素能够上调生物节律关键基因PER2、CRY1、Rev-erbα、RORα mRNA的震荡幅度,这表明雷帕霉素可能有助于延缓衰老导致的生物节律功能的下降;从蛋白质水平上,雷帕霉素处理显著降低了PER2蛋白的表达。由于PER2可以与BMAL1直接相互作用,并且抑制BMAL1的转录活性,因此雷帕霉素处理导致PER2蛋白表达降低,进一步解除了PER2对BMAL1下游基因的抑制作用,从而导致PER2、CRY1、Rev-erbα、Rorα mRNA表达上升。
衰老会伴随生物节律周期的延长,年轻老鼠的生物节律周期约为23.6 h,而老年老鼠约为24 h[9]。本研究显示,衰老的人皮肤成纤维细胞PER2 mRNA的节律周期大于24 h,而雷帕霉素处理可以缩短PER2的节律周期。雷帕霉素能够降低PER2蛋白表达,由于PER2蛋白稳定性的降低会明显缩短节律的周期[14],因此雷帕霉素可能是通过加速PER2蛋白的降解进而恢复PER2的节律周期。衰老与PER2蛋白的积累有非常紧密的联系[15],降低PER2表达可以明显延缓衰老导致的造血干细胞功能下降,进而延长老鼠寿命[16],雷帕霉素对于衰老的延缓作用是否通过PER2起作用有待进一步研究。研究发现,持续过表达PER2蛋白会显著缩短Rev-erbα基因的节律周期,并且抑制Rev-erbα基因的节律震荡水平。本研究结果显示,雷帕霉素处理会延长Rev-erbα基因的节律周期,增加Rev-erbα基因的节律震荡水平。其机制可能与PER2蛋白水平降低有关[17]。
综上所述,雷帕霉素可以提高衰老的人皮肤成纤维细胞生物节律关键基因BMAL1、PER2、CRY1、Rev-erbα、Rorα的表达,而且缩短PER2的节律周期,这意味着衰老所导致的节律钟功能的下降有可能被回复或逆转。这为雷帕霉素进一步应用于机体生物节律调控的研究提供了依据。
[1] Hood S, Amir S. The aging clock: circadian rhythms and later life[J]. J Clin Invest, 2017,127(2):437-446.
[2] Sahar S, Sassone-Corsi P. Metabolism and cancer: the circadian clock connection[J]. Nat Rev Cancer, 2009,9(12):886-896.
[3] Dubrovsky YV, Samsa WE, Kondratov RV. Deficiency of circadian protein CLOCK reduces lifespan and increases age-related cataract development in mice[J]. Aging (Albany NY), 2010,2(12):936-944.
[4] Kondratov RV, Kondratova AA, Gorbacheva VY, et al. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock[J]. Genes Dev, 2006,20(14):1868-1873.
[5] Bjedov I, Toivonen JM, Kerr F, et al. Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster[J]. Cell Metab, 2010,11(1):35-46.
[6] Vellai T, Takacs-Vellai K, Zhang Y, et al. Genetics: influence of TOR kinase on lifespan in C. elegans[J]. Nature, 2003,426(6967):620.
[7] Harrison DE, Strong R, Sharp ZD, et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice[J]. Nature, 2009,460(7253):392-395.
[8] Powers RW 3rd, Kaeberlein M, Caldwell SD, et al. Extension of chronological life span in yeast by decreased TOR pathway signaling[J]. Genes Dev, 2006,20(2):174-184.
[9] Chang HC, Guarente L. SIRT1 mediates central circadian control in the SCN by a mechanism that decays with aging[J]. Cell, 2013,153(7):1448-1460.
[10] Lipton JO, Yuan ED, Boyle LM, et al. The circadian protein BMAL1 regulates translation in response to S6K1-mediated phosphorylation[J]. Cell, 2015,161(5):1138-1151.
[11] Khapre RV, Kondratova AA, Patel S, et al. BMAL1-dependent regulation of the mTOR signaling pathway delays aging[J]. Aging (Albany NY), 2014,6(1):48-57.
[12] Cao R, Li A, Cho HY, et al. Mammalian target of rapamycin signaling modulates photic entrainment of the suprachiasmatic circadian clock[J]. J Neurosci, 2010,30(18):6302-6314.
[13] Matsumoto CS, Almeida LO, Guimaraes DM, et al. PI3K-PTEN dysregulation leads to mTOR-driven upregulation of the core clock gene BMAL1 in normal and malignant epithelial cells[J]. Oncotarget, 2016,7(27):42393-42407.
[14] Toh KL, Jones CR, He Y, et al. An hPer2 phosphorylation site mutation in familial advanced sleep-phase syndrome[J]. Science, 2001,291(5506):1040-1043.
[15] Asher G, Gatfield D, Stratmann M, et al. SIRT1 regulates circadian clock gene expression through PER2 deacetylation[J]. Cell, 2008,134(2):317-328.
[16] Wang J, Morita Y, Han B, et al. Per2 induction limits lymphoid-biased haematopoietic stem cells and lymphopoiesis in the context of DNA damage and ageing[J]. Nat Cell Biol, 2016,18(5):480-490.
[17] Chen RM, Schirmer A, Lee Y, et al. Rhythmic PER abundance defines a critical nodal point for negative feedback within the circadian clock mechanism[J]. Mol Cell, 2009,36(3):417-430.
Regulatoryeffectofrapamycinonkeygenesofcircadianrhythminsenescentcells
LIU Zuojun, LIU Baohua, MENG Fanbiao
(Shenzhen University, Shenzhen 518060, China)
ObjectiveTo investigated the effect of rapamycin on the cell autonomous circadian rhythm of senescent cells.MethodsWe induced replicative senescence of human dermal fibroblasts by series passage. The senescent human dermal fibroblasts cells were identified by SA-β-gal staining. The senescent human dermal fibroblasts cells were synchronized by 50% horse serum, and treated with DMSO or 200 nmol/L rapamycin. The oscillation profiles of BMAL1, PER2, CRY1, Rev-erbα and Rorα were examined by fluorogenic quantitative PCR. The circadian rhythm of genes was analyzed by JTK-CYCLE software. The protein expression of BMA1, CRY1, and PER2 was detected by Western blotting.ResultsThe mRNA expression of PER2, CRY1, Rev-erbα and Rorα in the observation group was higher than that of the control group (allP<0.05). The biorhythm cycle of BMAL1, CRY1, Rev-erbα, and Rorα genes in the control group was 24 h, and the biorhythm cycle of PER2 gene was 28 h. The biorhythm cycle of BMAL1, CRY1, and Rorα genes in the observation group was 24 h, and the biorhythm cycle of Rev-erbα gene was 27.5 h. The expression of CRY1 protein in the observation group was significantly higher than that in the control group, while PER2 protein expression was significantly lower than that in the control group (bothP<0.05). No significant difference was found in BMAL1 mRNA and protein expression between these two groups.ConclusionRapamycin can increase the expression of BMAL1, PER2, CRY1, Rev-erbα and Rorα mRNA in the aging human dermal fibroblasts and shorten the rhythmic cycle of PER2, which plays a regulatory role in the disturbance of circadian clock function.
biorhythm; rapamycin; key gene of biorhythm; aryl hydrocarbon receptor nuclear translocator 3; PER2; cryptochrome 1; orphan nuclear receptor α; orphan nuclear receptor α; mammalian target of rapamycin
10.3969/j.issn.1002-266X.2017.42.007
Q419
A
1002-266X(2017)42-0025-04
国家自然科学基金资助项目(81601215)。
刘佐君(1982-),男,博士后,主要研究方向为生物节律钟调控衰老的分子机制。E-mail: ijing1982@szu.edu.cn
2017-04-18)
