人工老化对小麦种子活力及醇溶蛋白组成的影响
2017-12-01,,,2,,,
, , ,2, , ,
(1.河南科技学院, 河南 新乡 453003; 2.新乡市第二十八中学, 河南 新乡 453000)
人工老化对小麦种子活力及醇溶蛋白组成的影响
张自阳1,马景周1,王智煜1,2,朱启迪1,茹振钢1,刘明久1
(1.河南科技学院, 河南 新乡 453003; 2.新乡市第二十八中学, 河南 新乡 453000)
为揭示种子活力与醇溶蛋白组成变化的关系,以小麦品种百农矮抗58、周麦18、新麦208、郑麦9023和杂交小麦(BNS/05525)为材料,高温(43±1)℃、高湿(95%相对湿度)老化处理0,4,6,9,12,15 d和20 d,对老化处理种子的活力及醇溶蛋白组成进行了分析。结果表明,随着老化时间的延长,小麦种子的发芽势、发芽率、发芽指数和活力指数逐渐降低,老化20 d的小麦种子发芽势、发芽率、发芽指数和活力指数均降为0,种子活力丧失。在同一老化程度下,杂交小麦种子的发芽势、发芽指数、活力指数均高于百农矮抗58、周麦18、新麦208、郑麦9023,说明杂交小麦的抗老化能力较强;电导率在种子老化的0~12 d内变化较小,种子老化到15 d时电导率迅速升高;5个小麦品种种子老化20 d后,发芽率从100%下降到0,醇溶蛋白的表达均出现了带型的变化,ω区醇溶蛋白出现了谱带的丢失、新蛋白的增加及蛋白表达量的上升和下降。种子活力丧失与蛋白的丢失、新谱带的产生都是在同一时间(种子老化20 d),说明种子活力的丧失与ω区域醇溶蛋白的消失和新增加的蛋白质有关;种子老化程度的加剧,导致了某些存活能力差的基因型消失,消失的基因可能与种子活力的表达相关。
人工老化; 小麦; 种子活力; 醇溶蛋白
种子是农业生产中的基本资料,种子的质量决定了农业生产的质和量,而种子活力是种子质量的重要指标,它与田间出苗、幼苗生长势和健壮度关系密切。种子在贮藏过程中的劣变或老化[1]不可避免,农业生产中经常会使用经过贮藏的种子,储藏不当的种子发芽率下降,田间播种导致断垄缺苗,给农业生产造成巨大损失。研究表明,种子活力与种子贮藏蛋白的关系密切[2],许多研究将种子蛋白质的变化作为种子活力的研究热点[3-5]。在种子老化过程中,种子贮藏物质如可溶性糖、蛋白质等经历了一个动态变化的过程。种子贮藏蛋白中的醇溶蛋白在种子萌发后为幼苗的生长提供各种氨基酸和含氮化合物,对种子的萌发和早期生长作用重大[6-8]。但是,对于种子活力、种子萌发与其醇溶蛋白组成和表达量变化关系尚不明确,对于种子活力丧失的机制尚不完全清楚,笔者于2014年研究了老化20 d的小麦种子醇溶蛋白组成的变化,发现老化20 d小麦种子醇溶蛋白ω区域的蛋白质出现了丢失和部分增加的现象,初步推断丢失的蛋白质与种子活力存在相关性[9]。为了进一步验证试验结果,探讨种子活力下降过程中醇溶蛋白的变化,探究种子活力与醇溶蛋白组成变化的关系,本试验以5个小麦品种为材料,通过人工老化的方法逐步降低小麦的种子活力,研究种子活力下降过程中种子活力及醇溶蛋白组成的变化,进一步揭示种子活力下降与醇溶蛋白组成变化的关系。
1 材料与方法
1.1 材 料
供试材料为百农矮抗58、新麦208、郑麦9023、周麦18、杂交小麦(BNS/05525),由河南科技学院小麦中心提供。
1.2 方 法
1.2.1 人工加速老化处理
人工加速老化处理采用张自阳等[9]的方法,试验材料放于LH-150 S种子老化箱[温度:(43±1 )℃;相对湿度:95%]中分别处理20,15,12,9,6,4 d,处理后自然晾干,放入4 ℃冰箱备用,以未老化处理的种子为对照。
1.2.2 种子发芽率、发芽势、发芽指数、活力指数的测定
采用张自阳等[10]的方法做标准发芽试验,每个品种100粒种子,重复4次,在20 ℃人工气候箱中培养,每天记载发芽数,第4天统计发芽势,第7天统计发芽率,发芽结束测定正常单株幼苗鲜重,并计算发芽势、发芽率、发芽指数和活力指数。
发芽势(%)=第4天发芽种子数/检测种子总数×100%;
发芽率(%)=第7天发芽种子数/检测种子总数×100%;
发芽指数(GI)=∑(Gt/Dt);
活力指数(VI)=GI×S;
式中:Gt为当天的发芽数,Dt为当日天数,S为发芽结束时单株幼苗鲜重。
1.2.3 电导率的测定,醇溶蛋白提取及聚丙烯酰胺凝胶电泳
采用张自阳等[9]的方法测定种子电导率、提取醇溶蛋白进行聚丙烯酰胺凝胶电泳。
2 结果与分析
2.1 老化处理对小麦发芽势、发芽率的影响
从图1可知,5个小麦品种的种子随着老化时间的延长,种子发芽势、发芽率随着老化天数增加呈逐渐下降趋势,老化20 d的小麦种子发芽率、发芽势均为0,表明种子已完全失去活力。老化4~9 d的种子发芽率下降缓慢,老化15 d的种子发芽率呈急剧下降的趋势;百农矮抗58、周麦18、杂交小麦在老化4~9 d内,种子发芽势下降缓慢,老化到第12天时发芽势急剧下降。新麦208、郑麦9023在老化至9 d时发芽势骤降。
2.2 老化处理对小麦发芽指数、活力指数的影响
由图2可知,5个小麦品种的发芽指数、活力指数随着老化程度的加深而逐渐降低。当老化处理时间达到20 d时发芽指数、活力指数为0。5个小麦品种的发芽指数在老化0~6 d时下降缓慢,当老化至9 d时发芽指数下降迅速。发芽指数在种子老化至6 d时表现为急速下降。
从图2结果还可知,杂交小麦在不同的老化程度下,发芽指数、活力指数均高于百农矮抗58、周麦18、新麦208、郑麦9023,表明杂交小麦的抗老化能力较强。
2.3 老化处理对小麦种子电导率的影响
由图3可知,5个小麦品种的种子在老化0~12 d内,电导率变化比较平缓,当种子老化15 d时,种子电导率迅速增大。
2.4 老化处理对小麦种子醇溶蛋白组成的影响
由图4,5,6,7,8可知,与未老化处理种子相比,5个小麦品种种子经酸性聚丙烯酰胺凝胶电泳后,人工老化4,6,9,12 d和15 d的小麦种子谱带没有发生变化,人工老化20 d的种子醇溶蛋白出现了谱带丢失、增加、表达量的上升和下降的现象。
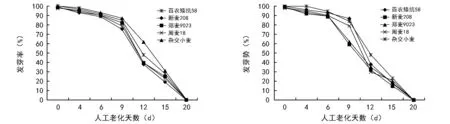
图1 人工老化对不同小麦品种发芽率、发芽势的影响

图3 人工老化对不同小麦品种电导率的影响
由图4可以看出,与对照相比,老化20 d后,百农矮抗58醇溶蛋白ω蛋白区ω 2、ω 3完全消失。在老化的4~15 d内,随老化天数的增加百农矮抗58的醇溶蛋白ω蛋白区ω 4、ω 6、ω 7、ω 8、γ蛋白区的γ 1表达量有所增加。
由图5可知,新麦208在老化20 d后醇溶蛋白ω 3谱带丢失、同时出现了新的ω 4、ω 8 2条谱带。
由图6可知,郑麦9023在老化20 d后醇溶蛋白ω 1表达量升高,ω 2蛋白谱带丢失,新出现了ω 3、ω 4、ω 6 三条醇溶蛋白谱带。
由图7可以看出,老化20 d的周麦18种子醇溶蛋白ω区ω 2谱带消失,ω 4表达量严重下降,几乎消失,同时出现了1条表达量比较弱的ω 3谱带。
由图8可以看出,杂交小麦在老化20 d后醇溶蛋白ω 3、ω 4谱带消失,ω 2表达量降低,新出现了1条ω 0谱带。
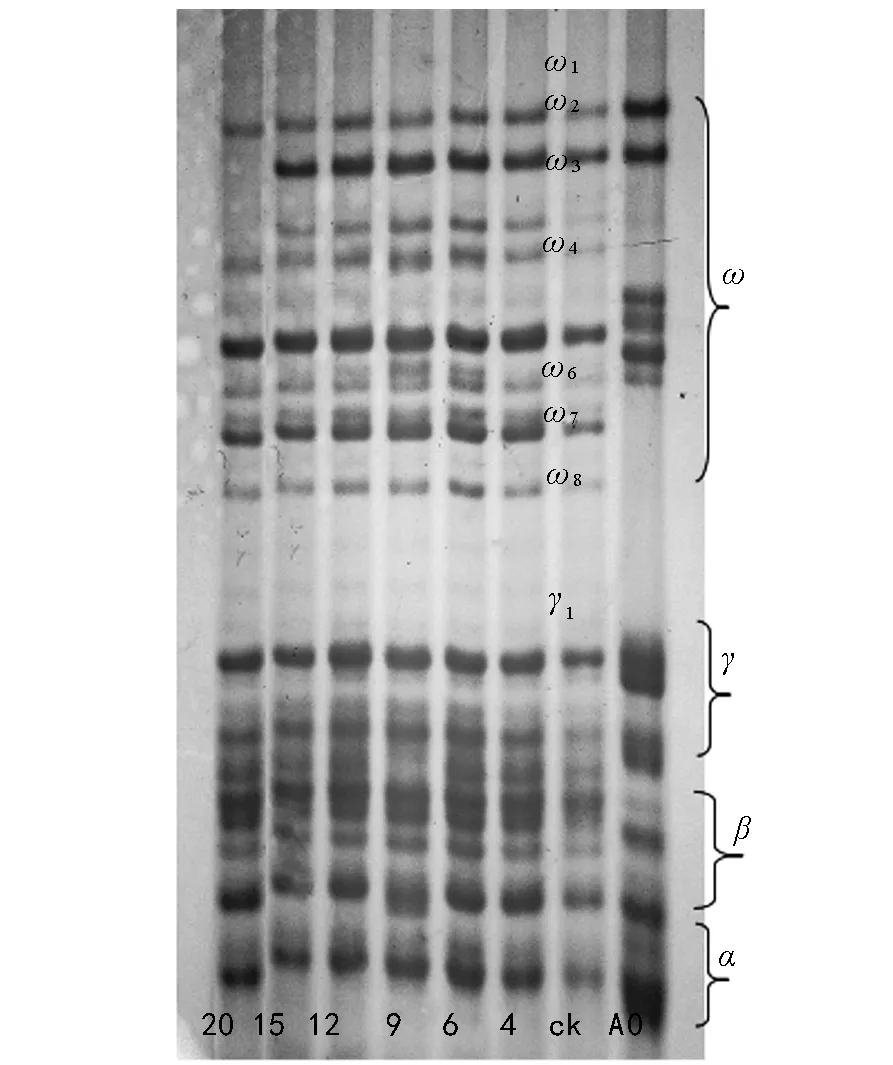
注:A0为中国春;ck为未老化种子;4,6,9,12,15,20 分别代表老化4,6,9,12,15,20 d的种子。下同。图4 不同老化程度百农矮抗58种子醇溶蛋白电泳图谱
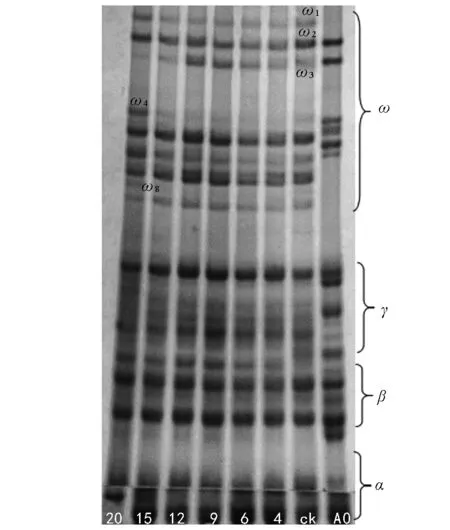
图5 不同老化程度新麦208种子醇溶蛋白电泳图谱
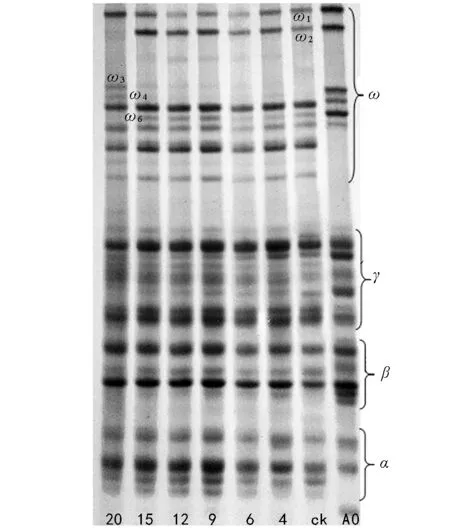
图6 不同老化程度郑麦9023种子醇溶蛋白电泳图谱
3 讨 论
由于高温、高湿环境可以加速种子衰老,同时又是研究种子活力衰退的一种快速方法。由结果分析可知,5个小麦品种种子经过人工老化后,发芽率、发芽势、发芽指数、活力指数均随老化时间的延长呈现下降趋势,说明人工老化导致了小麦种子活力下降。反映种子活力变化的发芽指标变化幅度依次为活力指数>发芽指数>发芽势>发芽率,说明活力指数能更灵敏地反映种子活力水平和老化程度,这与刘旭欢等[12-13]研究结果一致。同时可以看出,杂交小麦与百农矮抗58、周麦18、郑麦9023、新麦208相比,在老化程度一样的情况下,杂交小麦种子的发芽率、发芽势、发芽指数、活力指数均高于其它4个常规品种,说明杂交小麦的抗老化能力较强。
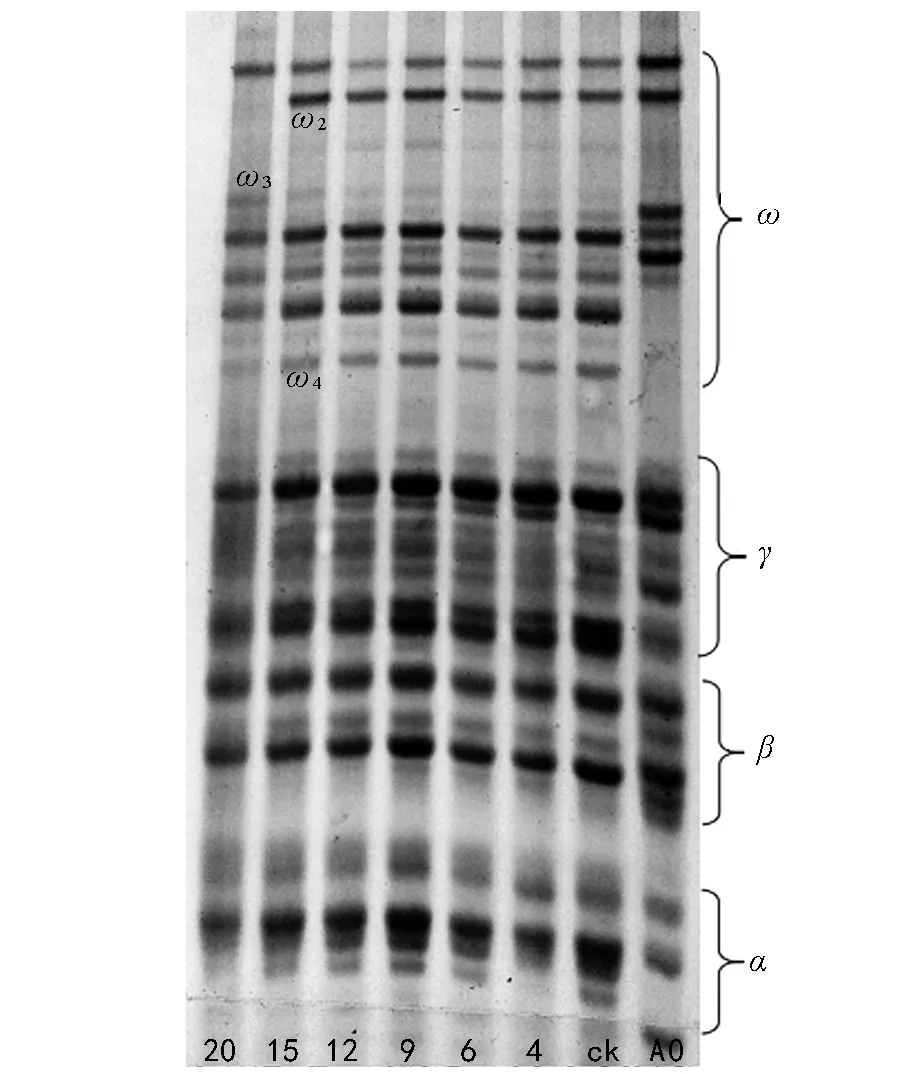
图7 不同老化程度周麦18种子醇溶蛋白电泳图谱

图8 不同老化程度杂交小麦种子醇溶蛋白电泳图谱
电导率测定法是测定种子活力较简易、快速的方法,有研究表明,电导率与人工老化种子活力之间密切相关[14-16]。本研究结果表明,5个小麦品种的种子在老化0~12 d内,电导率变化较小,当种子老化15 d时,种子电导率才迅速增大,种子活力的下降与电导率的变化相关性较小。
种子老化过程中内部会发生了一系列的生理化学反应,诱导相关基因表达的蛋白质的种类、数量或存在形式也因此发生改变[17]。种子中的贮藏蛋白在种子老化期间下降[18-19],甚至会出现新的蛋白质[9,20]。本研究结果表明,5个小麦品种的种子老化20 d,发芽率从100%下降到0,醇溶蛋白的表达出现了带型的变化,即谱带(醇溶蛋白)的丢失及新蛋白的增加,这与张晗等[21]的研究结果相似。百农矮抗58的醇溶蛋白ω 4、ω 6、ω 7、ω 8、γ蛋白区的γ 1表达量有所增加;新麦208 ω蛋白区ω 1表达量增加,同时出现了ω 4、ω 8两条新谱带;郑麦9023 ω蛋白区ω 1表达量升高,新出现了ω 3、ω 4、ω 6 3条谱带;周麦18种子醇溶蛋白ω区ω 4表达量严重下降,几乎消失,出现了1条表达量比较弱的ω 3谱带;杂交小麦在老化20 d后醇溶蛋白ω 3、ω 4谱带消失,ω 2表达量降低,新出现了1条ω 0谱带。老化20 d的小麦种子发芽率为0,种子活力丧失,种子失活与蛋白的丢失、新谱带(醇溶蛋白)的产生是在同一时期,说明种子活力与消失和增加的蛋白质有关,这与吴晓亮等的研究结果相似[22-23]。种子萌发需要蛋白质的合成[24],蛋白表达量的下调引起种子不能合成新的蛋白质,导致植物体发芽所需的能量供应不足,从而降低了种子发芽率。蛋白的消失使种子萌发失去能量来源导致种子失活,种子贮藏物质中蛋白质分解应是种子老化或种子活力丧失的显著特征及最可能的原因。醇溶蛋白是种子发育特定时期的基因产物。它的基因定位在第1和第6部分同源染色体的短臂上,全部γ和ω醇溶蛋白基因位于第1部分同源染色体短臂上,编码基因为Gli-1。种子老化到一定程度时外在逆境使遗传组成可能发生了变化,随着种子老化的加剧,导致了某些存活能力差的基因型的消失[25],消失的基因可能与种子活力的表达相关。
[1]Elena A G Hank V A,Folkert A H.Membrane chemical stability and seed longevity[J].European Biophysics Journal,2010,39(4):657-668.
[2]刘军,黄上志,傅家瑞,等.种子活力与蛋白质关系的研究进展[J].植物学通报,2001,18(1):46-51.
[3]王继红,陈绍宁,白冰,等.人工老化对玉米种子蛋白质组的影响[J].河南农业大学学报,2009,43(3):232-235.
[4]周峰.种子贮藏蛋白质丰度研究进展[J].河南农业科学,2015,44(1):1-3,76.
[5]舒英杰,陶源,王爽,等.高等植物种子活力的生物学研究进展[J].西北植物学报,2013,33(8):1 709-1 706.
[6]Muntz K,Belozersky MA,Dunaevsky YE,et al.Stored proteinases and the initiation of storage protein mobilization in seeds during germination and seedling growth[J].Journal of Experimental Botany,2001,52(362):1 741-1 752.
[7]Tiedemann J,Neubohn B,Muntz K. Different functions of vicilin and legumin are reflected in the histopaettern of globulin mobilization during germination of vetch (ViciasativaL.)[J].Planta,2000,211(1):1-12.
[8]Tiedemann J,Schlereth A,Muntz K.Differential tissue-specific expression of cysteine proteinases forms the basis for the fine-tuned mobilization of storage globulin during and after germination in legume seeds[J].Planta,2001,212(5-6):728-738.
[9]张自阳,茹振钢,刘明久,等.人工老化小麦种子的活力与醇溶蛋白、谷蛋白组成分析[J].河南农业科学,2014,43(2):24-27.
[10]张自阳,朱俊华,程媛,等.不同成熟度小麦种子活力及其与生理性状的相关性研究[J].河南农业科学,2014,43(12):6-9.
[11]张自阳,姜小苓,茹振钢,等.人工老化对杂交小麦种子生理特性和种子活力变化的影响[J].江苏农业科学,2013,41(2):81-83.
[12]刘旭欢,贾永红,阿里别里根·哈孜太.人工老化对春小麦种子萌发及根尖超微结构的影响[J].西北植物学报,2015,35(4):761-765.
[13]刘旭欢,加得拉·努尔兰,童婷,等.不同老化时问和收获期对春小麦种子活力的影响[J].新疆农业科学,2014,51(8):1 394-1 399.
[14]梁海荣,邢侗姬,周小梅.不同活力的小麦种子生理生化及其修复的研究[J].山西大学学报(自然科学版),2010,33(2):286-290.
[15]张玲丽,郭月霞,宋喜悦.不同类型小麦品种人工老化处理后种子活力特性的研究[J].种子,2008,27(10):52-55.
[16]朱银,颜伟,杨欣,等,电导法测定小麦种子活力[J],江苏农业科学,2014,42(9):78-80.
[17]Hochholdinger F,Sauer M,Dembinsky DN,et al.Proteomic dissection of plant development[J].Proteomics,2006,6(14),4 076-4 083.
[18]Ching TM,Schoolcraft I.Physiological and Chemical Differences in Aged seeds[J].Crop Science,1968,8(4):407-409.
[19]范国强,秦文静,刘玉礼.花生种子人工老化过程中发芽率和蛋白质的变化[J].河南农业大学学报,1995(4):337-340.
[20]王继红,陈绍宁,白冰,等.人工老化对玉米种子蛋白质组的影响[J].河南农业大学学报,2009,44(3):232-235.
[21]张晗,卢新雄,张志娥,等.种子老化诱导小麦染色体畸变及大麦醇溶蛋白带型频率变化的研究[J].植物遗传资源学报2004,5(1):56-61.
[22]吴晓亮,陈晓玲,辛萍萍,等.超干燥处理对豌豆种子抗氧化系统酶及热稳定蛋白的影响[J].园艺学报,2006,33(3):523-528.
[23]董贵俊.向日葵(HelianthusannuusL.)种质资源的基础研究[D].北京:中国科学院,2006.
[24]Rajjou L,Gallardo,K,Debeaujon L,et al.The effect of α-amanitin on the Arabidopsis seed proteome highlights the distinct roles of stored and neosynthesized mRNAs during germination[J].Plant Physiology,2004,134(4):1598-1613.
[25]Parzies H K,Spoor W,Ennos R A.Genetic diversity of barley landrace accessions (Hordeumvullgaressp.vulgare)conserved for different lengths of time in ex situ gene banks.Heredity,2000,84(4):476-486.
Effects of Aging Treatment on Seed Vigor and Constitutes of Gliadin in Wheat
ZHANGZiyang1,MAJingzhou1,WANGZhiyu1,2,ZHUQidi1,RUZhen’gang1,LIUMingjiu1
(1.He’nan Institute of Science and Technology,Xinxiang He’nan 453003,China; 2.Xinxiang No.28 Middle School,Xinxiang He’nan 453000,China)
In order to investigate the relationship of seed vigor and gliadin composition,Bainong aikang 58,Zhoumai 18,Xinmai 208,Zhengmai 9023 and hybrid wheat seed(BNS/05525) were studied by using artificial accelerated aging(temperature:(43±1)℃,RH:95%) with aging 0,4,6,9,12,15 and 20 days.The seeds vigor and gliadin composition which were aged were analyzed.The result show that wheat seed germination potential,germination rate,germination index,vitality index were gradual decrement trend,the wheat seed losing germination ability when it aging 20 days.The germination germination potential,germination rate,germination index,vitality index of hybrid wheat seed were higher than Bainong aikang 58,Zhoumai 18,Xinmai 208,Zhengmai 9023 under the same aging degree,which illustrate hybrid wheat with a strong anti-aging ability.The conductivity of seeds were changes slightly within the seed aging 0-12 days,it rise rapidly when the seed aging to 15 days.The gliadin composition were changed and the loss of the band,the new protein increased and the rise and fall of the amount of protein expression,the seed germination percentage decreased from 100% to 0 after aging of 20 d.The loss of seed vigor were connected with the disappearance of the gliadin and the appearance of new proteins,because they happened at the same time. Because of the intensification of seed aging,some poor survivability genotype were disappeard,which the disappearance of genes may be associated with the expression of seed vigor.
artificial aging; wheat; seed vigor; gliadin
2017-01-10
科技部创新方法工作专项项目(2015 IM 010400);河南省科技计划项目(162300410142);河南科技学院青年教师资助计划(201010714004)。
张自阳(1984—),男,河南舞钢人;实验师,硕士,主要从事作物遗传育种研究;E-mail:lanfeng0615@163.com。
刘明久(1966—),男,河南获嘉人;教授,主要从事种子学的教学和科研工作;E-mail:liumingjiu@hist.edu.cn。
10.16590/j.cnki.1001-4705.2017.08.042
S 512.1
A
1001-4705(2017)08-0042-06
