洪泽湖河蚬空间分布及资源量研究
2017-11-30张彤晴唐晟凯李大命刘燕山刘小维杨俊虎朱忠穆欢黄越峰
张彤晴+唐晟凯+李大命+刘燕山+刘小维+杨俊虎+朱忠+穆欢+黄越峰


摘要:2014年5月至2015年2月对洪泽湖河蚬(Corbicula fluminea)种群动态和资源量进行了周年调查。洪泽湖河蚬周年平均密度和生物量分别为5.72 ind./m2和28.82 g/m2;资源量呈现明显的季节变化,夏季达到最大值(4.15×107 kg);湖区资源量分布呈西高东低态势,即老子山-临淮片区>成子湖片区>洪泽片区,而空壳率则表现相反的变化特征。分析表明,河蚬资源量的季节变动主要是受温度的影响,夏末秋初为捕捞河蚬的最佳季节;而水深和食物来源是影响洪泽湖河蚬空间分布的主要因素,人工增殖时应选择水深在1.3~2.2 m、食物来源丰富的水域;大部分湖区总氮已经超出渔业用水标准,且总氮已经对河蚬的资源量分布造成了一定的负面影响;洪泽湖河蚬的年资源量在0.82×107~4.15×107 kg,与历史数据相比,年资源量持续降低。
关键词:洪泽湖;河蚬;空间分布;资源量
中图分类号: S931.1 文献标志码: A 文章编号:1002-1302(2017)20-0180-07
河蚬(Corbicula fluminea)为瓣鳃纲双壳类软体动物,是江苏湖泊重要的经济水产品之一。洪泽湖有优质的河蚬种质资源,不仅是我国河蚬的重要产区,亦是河蚬增殖放流苗种的重要原产区。然而近十年来随着采沙、过度捕捞等人类活动的增加,湖泊水生态环境发生变化,河蚬的生存环境遭到不同程度的破坏,致使河蚬呈现出资源量锐减、种质退化、繁殖力下降、空壳率高等趋势,因此亟需采取有力手段对河蚬资源进行保护。目前,国内有关河蚬的研究主要集中在其生物学[1]、营养价值[2-3]、遗传多样性[4-5]及作为环境指示生物[6]等方面。关于洪泽湖底栖生物群落结构的报道较多[7-10],已有学者研究了其他水域河蚬的时空分布[11-12],而有关洪泽湖河蚬资源量、空壳率、规格的等级、时空分布以及环境因子的关系未见报道。笔者于2014年5月至2015年2月对洪泽湖河蚬进行了周年采集,探索洪泽湖河蚬的种群时空分布变化情况,以期为湖泊底栖动物种群动态及合理利用提供理论依据。
1 材料与方法
根据洪泽湖环境空间异质性,对洪泽湖河蚬集中分布区域(面积900 km2),采用ArcGIS均匀布设采样点29个,分为3片区。其中洪泽片区调查10个点,编号为1~10;老子山-临淮片区调查11个点,编号为11~21;成子湖片区调查8个点,编号为22~29。于2014年5月(春)、8月(夏)、11月(秋)和2015年2月(冬)对洪泽湖河蚬进行了4次定量采样。采样工具为渔民捕捞河蚬用的划耙(图1),耙齿间距为8.00 mm,宽70 cm,采样距离10.0 m,样方7.0 m2(0.7 m×10.0 m)。每点采集1次,重复1次。河蚬采样的同时采集水样。划耙采集样品带回实验室,经60目纱网筛沥干后,称总质量,随机取样称质量,挑出空壳称质量,计算空壳率。每个采样点随机取样30个(采样点若少于30个活样本,则全部取样),置于解剖盘中分检,用游标卡尺(恒量,0.01 mm,0.00~150.00 mm)测量壳长(L,0.10 mm),用电子天平(CANYMP1002,0.01 g,0.00~100.00 g)称取体质量(m,0.01 g)。值得注意的是,河蚬测量和称质量前,先用滤纸将待测标本体表的水分吸去,直至滤纸上没有湿的印迹为止。按HJ 636—2012[13]测水样总氮,按GB/T 11893—1989[14]测水样总磷。
2 结果与分析
2.1 河蚬的密度与生物量周年变化
洪泽湖河蚬周年平均密度为5.72 ind./m2,周年平均生物量为28.82 g/m2。图2为河蚬生物量与密度季节变化情况,河蚬生物量和密度均呈现明显的季节变化,夏季其密度和生物量值均为最大,分别达到9.57 ind./m2和 46.15 g/m2;冬季均降至最低,分别为1.57 ind./m2和9.08 g/m2。分析表明,春季和夏季河蜆的生物量和密度均显著高于秋季和冬季(P<0.05)。
2.2 河蚬壳长与湿质量的关系
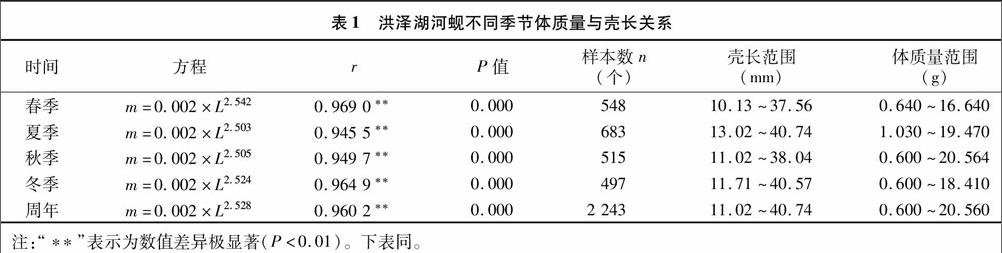
用幂函数关系公式“m=a×Lb”来拟合河蚬体质量(湿质量:m,g)与壳长(L,mm)的关系。河蚬的壳长和体质量之间呈显著的正相关关系,m=0.002×L2.528(r2=0.922 0)。共测定2 243个河蚬个体,壳长和体质量范围分别为11.02~4074 mm和0.60~20.56 g。
2.3 洪泽湖河蚬的空间分布格局
2.3.1 洪泽湖河蚬年均分布格局 利用地图汇软件对洪泽湖河蚬年平均生物量和年平均密度的空间等级分布格局进行表征(图3)。可以看出洪泽湖河蚬的生物量空间分布不均(图3左),呈现从东到西逐渐升高态势,以洪泽湖东部和东南部湖区河蚬的生物量较少,成子湖片区次之,老子山-临淮片区最高,其中大于90 g/m2的点有15号、16号和26号点(分别为117.21、125.95、92.50 g/m2)。洪泽湖河蚬年平均密度的分布格局图显示(图3右),亦为老子山-临淮片区最高,其高值点主要有15号和16号2个(分别为 23.17 ind./m2和 27.39 ind./m2),其次为12号、18号、26号点(分别为17.79、14.50、18.13 ind./m2),各点的密度均小于10 ind./m2。
2.3.2 洪泽湖河蚬生物量不同季节的空间分布 从不同季节生物量空间分布格局(图4)可以看出,春季(5月)洪泽湖河蚬的生物量以老子山-临淮片区为高值区(大于 100 g/m2),15号、16号、17号、18号点集中连片分布,分别为110.40、189.19、103.43、147.84 g/m2。周边的12号、19号、20号、21号、26号点的生物量介于40~100 g/m2,分别为9290、46.46、60.20、53.07、63.31 g/m2,呈现高值区向周边站位逐渐降低状态,其余20个点生物量均在40 g/m2以下。夏季(8月)洪泽湖河蚬的生物量仍以老子山-临淮片区为高值区,且高值区域进一步扩大,生物量也持续升高,继续呈现区域连片高值态势。其中12号、13号、15号、16号、18号、21号点分别达到175.07、107.16、163.96、256.90、108.06、11066 g/m2。19号、20号、26号点也达到63.49、62.87、9915 g/m2。其余20个点均小于40 g/m2。而秋季(11月)河蚬生物量则在老子山-临淮片区和成子湖片区出现2个高值点,高值区面积缩小为斑块分布,其中15号、26号点最高,分别为176.67、191.42 g/m2;11号、28号次之,为54.94、4379 g/m2,其余25个点均低于40 g/m2。冬季(2月)是一年中河蚬生物量最低的时期,各片区均未出现大于100 g/m2的区域,但最高值仍出现在老子山-临淮片区和成子湖片区,16号、21号点最高分别为52.76、50.27 g/m2,其余各点均小于20 g/m2。把生物量分为7个等级,可以看出7个等级在不同季节出现的站位数各不相同,即所覆盖的水域面积也不相同。随着本调查的季节变动,小于 10 g/m2 的区域呈现逐步扩大-平稳-稍下降趋势。根据采样站位所代表水域面积换算资源现存量,得出2014年5月为3.41×107 kg,8月为 4.15×107 kg,11月为1.99×107 kg,2015年2月为 0.82×107 kg,呈现先增长后下降的变化趋势。endprint
2.3.3 洪泽湖河蚬密度不同季节的空间分布 同样把不同季节密度分为5个等级对河蚬生物量空间分布格局进行表征(图5)可以看出,洪泽片区及成子湖片区除26号点出现密度等级波动(14.50~19.21~37.00~1.79 ind./m2)外,其余站位密度均小于10 ind./m2,等级密度季节变化较明显的水域主要为老子山-临淮水域。5月大于40 ind./m2的仅有16号点(49.43 ind./m2),8月为16号点(49.71 ind./m2)和12号点(42.29 ind./m2),到了11月和2月该密度等级的站点均未出现;30 ind./m2-40 ind./m2密度等级在5月未出现,到了8月为15号点(35.14 ind./m2)和18号点(34.71 ind./m2),11月为15号点(30.43 ind./m2)和26号点(37.00 ind./m2),呈点状分布。到了2月全湖密度均小于10 ind./m2。根据站位所代表水域面积换算资源数量,得出5月为7 210.64×106 ind.,8月为8 614.73×106 ind.,11月为3 353.50×106 ind.,2月为1 409.85×106 ind.,呈先增长后降低的季节变化趋势。
2.3.4 洪泽湖河蚬空壳率不同季节的空间分布 把不同季节空壳率分为5个等级进行表征,从不同季节空壳率空间分布格局(图6)可以看出,洪泽片区和成子湖片区水域的空壳率比老子山-临淮水域的高,与生物量区域等级的分布相反,从东到西呈现逐步降低态势。随着本调查的季节变换,高空壳率(大于80%)呈现由北到东南逐步扩散的态势,根据站位所代表水域面积换算空壳质量,得出:5月为1.72×107 kg,8月为3.66×107 kg,11月为0.79×107 kg,2月为0.71×107 kg,亦呈先增长后降低的变化趋势。
2.3.5 洪泽湖河蚬规格不同季节的空间分布 把不同季节规格分为5个等级进行表征(图7),从不同季节规格空间分布格局可以看出,5月规格等级以2~4 g/ind.的站点较多,达10个;到了8月、11月和2月4~6 g/ind.的站点较多,分别达11个、8个和11个;5月以8号、23号、25号点规格最小,呈现以这3个点向周围站点规格逐渐增大的布局,规格较大的分布在监测的边缘站点;8月和11月以成子湖片区相对规格较小,洪泽片区以有5个较大规格(6 g/ind~8 g/ind.)的点位而高于老子山-临淮片区的河蚬规格;2月小规格的河蚬分布于成子湖和洪泽片区,老子山-临淮片区则较高。
2.3.6 洪泽湖河蚬调查站位不同季节总氮浓度的空间分布 参照地表水环境质量标准基本项目标准(GB 3838—2002),Ⅲ类水为渔业水域用水标准,Ⅲ类水TN指标是 1.0 mg/L,Ⅳ类水为1.5 mg/L,Ⅴ类水为2.0 mg/L,把不同季节站位的总氮浓度分为5个等级进行表征,从不同季节总氮浓度的空间分布格局(图8)可以看出,总氮浓度总体呈现成子湖片区<老子山-临淮片区<洪泽片区。5月4个站点达Ⅲ类水,5个站点达Ⅳ类水,8个站点为Ⅴ类水,还有12个站点为劣Ⅴ类水。8月水质是一年中最好的,劣Ⅴ类水只有3个点位,大多数点位是Ⅲ~Ⅴ类水,成子湖水域的3个点(27号、28号、29号)达到Ⅲ类水标准;11月劣Ⅴ类水域面积继续扩大,增加到14个点位,主要集中在位于湖中部的洪泽片区和老子山-临淮片区。到了2月除少数点位外,大多数点位的总氮浓度进一步增加。
2.3.7 洪泽湖河蚬调查站位不同季节总磷浓度的空间分布 参照地表水环境质量标准基本项目标准(GB 3838—2002),Ⅲ类水为渔业水域用水标准,Ⅲ类水的TP指标是0.05 mg/L,Ⅳ类水为0.1 mg/L,Ⅴ类水为0.2 mg/L,把不同季节站位的总磷浓度分为5个等级进行表征,从不同季节总磷浓度的空间分布格局(图9)可以看出,5月水质为Ⅲ~Ⅳ类水,Ⅲ类水点位占大多数;8月大多点位总磷等级增加,以Ⅳ~Ⅴ类水为主;11月浓度有所降低,成子湖和老子山-临淮片区下降较多,洪泽片区则变化不大;到了2月洪泽片区有所下降,而成子湖和老子山-临淮片区变化不明显,总体水质改善为Ⅲ~Ⅳ類水。
2.3.8 洪泽湖河蚬资源量各因子之间相关分析 由表2可知,洪泽湖河蚬的生物量和密度之间显著正相关(r=0.963,P=0.000<0.01),其生物量与空壳率之间(r=-0.282,P=0.002<0.01),密度与空壳率之间(r=-0.244,P=0.008<0.01)以及规格与空壳率之间(r=-0.454,P=0.000<0.01)则呈现极显著负相关,而总氮、总磷与生物量、密度、规格、空壳率之间在统计学上的相关性均不显著。
3 小结与讨论
3.1 河蚬捕捞工具的优化
使用不同的采样工具往往会影响资源调查结果的准确性,因此采样工具的选择尤为重要。目前,采集河蚬所用工具有彼得森采泥器和划耙。彼得森采泥器的优势是可以采集到小规格的河蚬,缺点是代表性不够,随机性大; 而用划耙采集样品,采集面积大,样本全,更具有代表性,更加接近于实际资源量。因此本次调查采用划耙工具,一次性采样面积达到 7 m2,是常规底栖生物调查工具彼得森采泥器(1/16 m2)的112倍,所采集河蚬的规格为11.02~40.07 mm,范围远大于蔡炜采用彼得森采泥器的采集的太湖河蚬规格(1.50~21.43 mm)[11],以及稍大于毕婷婷等用划耙采集的洪泽湖河蚬规格(8.5~31.4 mm)[10],毕婷婷等所用的划耙耙间距(60 mm)亦小于本次调查(8.0 mm)。在计算壳长与体质量关系时,有文献报道,当计算样本包括所有壳长范围的河蚬时,b值趋近于3[11],而本研究的生长指数b值为2.528,尽管与毕婷婷等的报道(2.44)[10]相近,但表明用划耙采样仍有缺陷。综上所述,划耙与彼得森采泥器结合使用是采集河蚬更好的方法,且彼得森采泥器的采样次数要足够多,才能更加全面准确地反映一个水域的河蚬资源量。endprint
3.2 洪泽湖河蚬资源量分布和季节变化分析
影响河蚬资源量和分布的因素很多,概括起来可分为3类:一是非生物因素,包括水流、底质、水深、总磷、氨氮、COD等理化因子,以及铜、镉等重金属元素;二是生物因素,包括细菌、浮游植物等食物,及水生植被、螺类、鱼类等竞争者;三是人为因素,包括捕捞、增殖放流等。
洪泽湖是一个大型浅水湖泊,湖底平坦,平均水深 1.77 m,最大水深4.37 m,而河蚬生长的最适水深为1.3~2.2 m[15];且底质类型为泥沙质底质[16],易受河流和风力影响而扰动,氧气含量较高,因此该湖适宜河蚬的生长。研究结果显示,洪泽湖河蚬的空间分布表现出明显的差异性,这种差异性可能受到多种因素的综合影响。本研究中,洪泽湖河蚬年生物量呈现从东到西逐渐升高态势,洪泽片区河蚬的生物量较少,成子湖片区次之,老子山-临淮片区最高,而空壳率则表现出相反的变化特征,这可能是由于河蚬的分布和水深有关,当水深在一定范围内河蚬的资源量随水深增加逐渐降低[17],这与洪泽湖水位呈东高西低的态势[18]相符。同时食物来源也是影响河蚬资源量的重要因素[11],以淮河为主的入湖河流主要在湖泊的西侧,该片区水体营养丰富,浮游植物含量也较高,因此老子山-临淮片区叶绿素含量相对较高,浮游藻类丰富,能够为河蚬提供更加充足的饵料,因此该片区河蚬资源量也相对较高;而成子湖片区含有丰富的水草,至使水体较为清澈,底质受扰动较小,浮游植物和悬浮颗粒物含量较低,河蚬的食物来源也相对较少[19],因此该片区河蚬资源量也相对较低。综上所述,水深和食物来源是影响河蚬资源量空间分布的主要因素。因此笔者建议,在进行河蚬的人工增殖放流时,应选择水深在1.3~2.2 m,且食物来源较丰富的水域。
研究表明,洪泽湖河蚬资源量呈明显的季节变化,春季和夏季显著高于秋季和冬季,这可能是受水温的影响。春季水温上升,河蚬开始正常摄食,且浮游植物等食物来源也开始增多,生物量开始增大,至夏季持续增长至最大值,而秋季资源量显著下降,冬季降到低谷,是由于该时期水温持续降低,导致河蚬逐渐停止摄食,且浮游植物等食物来源也逐渐减少,从而主要依靠消耗自身能量维持自身代谢,这与蔡炜等的研究结果[11]相似。因此,夏末秋初洪泽湖河蚬最肥,视为捕捞河蚬的最佳季节。
有研究表明,总磷、氨氮等尤其是COD的变化可能会对一个湖区河蚬的分布、密度等造成较大影响[20]。然而本研究结果显示洪泽湖水域的总氮、总磷与生物量、密度、规格、空壳率之间在统计学上的相关性均不显著,这可能是由于洪泽湖的总氮、总磷均未达到影响河蚬资源分布的阈值,且各片区相对差异不大。但从本研究结果中仍可看出总氮与这4个因子之间呈负相关,总磷除了与空壳率呈负相关外,与其余3个因子之间呈正相关。表明大多数站点总氮的浓度已经超出渔业用水标准,对河蚬资源量的分布已经造成了一定程度的负面影响,总磷浓度超出渔业用水标准的情况要好于总氮浓度,从统计学角度分析,还没有达到对河蚬资源量的分布造成负面影响的程度。
3.3 河蚬管理建议
本研究中河蚬年资源量在0.82×107~4.15×107 kg,仅为1990—1991年(21.29×107 kg)[7]的4%~19%,且低于2010年(7.84×107 kg)、2011年(6.92×107 kg)和2012年(5.0×107 kg)[10],可见洪泽湖河蚬资源量总体呈降低趋势,而且还在持续。尽管调查所用工具有差别,但这主要是受过度与非法捕捞、栖息环境破坏等因素的影响。
针对这一现状,提出以下建议:一是合理降低河蚬捕捞量,已有研究表明捕捞死亡系数上升是引起河蚬总死亡系数上升的主要因子[10];二是增大河蚬的开捕规格,例如2015年6月洪泽湖网公布的捕捞耙齿间距增加至1.1 cm,有利于河蚬种质资源的保护;三是加强执法力度,建议相关部门出台政策,如严格限制捕捞生产时间、生产区域等;四是加强科学示范,进行人工增殖,如网围增殖模式的应用[21],不仅能起到增殖的效果,还能够净化水质。
参考文献:
[1]刘月英,张文珍,王跃先. 中国经济动物志:淡水软体动物[M]. 北京:科学出版社,1979:119-123.
[2]王 宇,刘东红. 河蚬功能性成分的研究现状与展望[J]. 食品与发酵工业,2010(6):122-124.
[3]刘晶晶,徐云婕,韩曜平,等. 洪泽湖野生河蚬营养成分的分析与评价[J]. 食品工业科技,2014,35(15):360-364.
[4]李大命,张彤晴,唐晟凯,等. 洪泽湖野生河蚬(Corbicula fluminea)线粒体COⅠ基因序列的遗传多样性分析[J]. 渔业科学进展,2015,36(5):81-86.
[5]丁怀宇,姜虎成,冯建彬,等. 河蚬微卫星引物筛选及洪泽湖野生群体遗传结构分析[J]. 水产学报,2011,35(11):1624-1632.
[6]刘 敏,熊邦喜. 河蚬的生态习性及其对重金属的富集作用[J]. 安徽农业科学,2008,36(1):221-224.
[7]袁永浒,王兴元,陈安来,等. 洪泽湖螺蚬资源调查報告[J]. 水产养殖,1994(6):15-16.
[8]严维辉,潘元潮,郝 忱,等. 洪泽湖底栖生物调查报告[J]. 水利渔业,2007,27(3):65-66.
[9]张超文,张堂林,朱挺兵,等. 洪泽湖大型底栖动物群落结构及其与环境因子的关系[J]. 水生态学杂志,2012,33(3):28-33.
[10]毕婷婷,侯 刚,张胜宇,等. 基于壳长频率分析的洪泽湖河蚬渔获物年龄结构、生长与死亡参数估计[J]. 水生生物学报,2014,38(5):797-800.
[11]蔡 炜,蔡永久,龚志军,等. 太湖河蚬时空格局[J]. 湖泊科学,2010,22(5):714-722.
[12]李丽娜,陈振楼,许世远,等. 长江口滨岸带河蚬的时空分布特征及其指示作用[J]. 应用生态学报,2006,17(5):883-886.
[13]水质标准 总氮的测定:HJ 636—2012[S].
[14]水质标准 总磷的测定:GB/T 11893—1989[S].
[15]余 辉,张文斌,卢少勇,等. 洪泽湖表层底质营养盐的形态分布特征与评价[J]. 环境科学,2010,31(4):961-968.
[16]何华春,丁海燕,张振克,等. 淮河中下游洪泽湖湖泊沉积物粒度特征及其沉积环境意义[J]. 地理科学,2005,25(5):590-596.
[17]侯冠军,李 跃. 女山湖贝类资源调查和增殖措施的研究[J]. 水产科技情报,2002,29(2):66-70.
[18]楚恩国. 洪泽湖水文特性初探[J]. 水文,2001,21(5):56-59.
[19]刘伟龙,邓 伟,王根绪,等. 洪泽湖水生植被现状及过去50多年的变化特征研究[J]. 水生态学杂志,2009,2(6):1-8.
[20]凌去非,宋学宏,康华东,等. 昆承湖河蚬资源及水体理化因子的初步探讨[J]. 水利渔业,2001,21(1):44-46.
[21]王小林. 太湖渔业生态系统修复途径探讨[J]. 河海大学学报(自然科学版),2004,32(增刊1):147-150.endprint
