不同强度一次性运动对大鼠睾周白色脂肪FNDC5、UCP-1表达量的影响
2017-11-29徐瑞严翊谢敏豪
徐瑞严翊谢敏豪
1北京体育大学运动人体科学学院(北京 100084)
2国家体育总局运动医学研究所(北京 100061)
研究发现,具有促进白色脂肪棕色化作用的鸢尾素(Irisin)是通过剪切骨骼肌中细胞外跨膜受体Ⅲ型纤连蛋白域蛋白5(Fibronectin type III domain-containing 5 protein,FNDC5)的结构域所得,并以二聚体的形式表现。运动能够诱导肌肉中过氧化物酶体增殖物激活受体γ辅激活因子-lα(peroxisome proliferator-activated receptor gamma coactivator1 alpha,PGC-lα)的表达后进而激活FNDC5的表达,从而促进生成Irisin。FNDC5是Irisin的前体,因此,在研究中一般使用FNDC5来代表Irisin[1]。
当白色脂肪组织受到某些生理刺激(如寒冷暴露等)时,会出现棕色脂肪表性特征,即白色脂肪棕色化(Browning)[2,3],使机体耗能增加。在脂肪组织中,尤其是棕色脂肪组织中,主要是通过线粒体中的解偶联蛋白1(uncoupling protein 1,UCP-1)来发挥产热和耗能的作用,故UCP-1被公认为是脂肪组织中产热和耗能的标志性蛋白,可以作为检验脂肪组织中产热和耗能是否活跃的指标,是代表棕色脂肪的重要基因。
已有研究证明,运动可以促进“Browning”。但现有研究多集中于长时间的有氧运动干预,缺乏不同运动强度干预效果的对比研究。因此,为探究运动促进白色脂肪棕色化的有效负荷强度及运动效果的持续时间,本研究观察了一次性中等强度运动和高强度间歇运动后大鼠白色脂肪组织FNDC5和UCP-1的变化规律。
1 材料与方法
1.1 实验动物
8周龄SPF级雄性SD大鼠30只购自于北京市维通利华动物实验中心,许可证编号:SYXK(京)2011-0034。随机分为对照组(C组,n=6)和运动组(E组,n=24),其中E组大鼠根据运动强度分为中等强度运动组(EM组,n=12)和高强度间歇运动组(EH组,n=12),大鼠组间体重无显著性差异,具体情况见表1。再根据一次性运动后的取材时间分成中等强度运动后即刻组(EM0组,n=6)和高强度间歇运动后即刻组(EH0组,n=6)、中等强度运动后6h组(EM6组,n=6)和高强度间歇运动后6h组(EH6组,n=6)。大鼠分笼饲养,室温20℃~24℃,相对湿度50%~70%,每日12小时光照/12小时熄灯,国家标准啮齿类动物常规饲料喂养,大鼠自由饮水、饮食。

表1 各组大鼠体重情况
1.2 运动方案
C组大鼠安静饲养,E组大鼠经3天适应性训练后,随机分为中等强度运动(EM)组和高强度间歇运动(EH)组(大鼠跑台 DSPT-208,杭州段式制造)。运动模型参照本实验室前期对大鼠最大摄氧量的测试结果[4,5]以及Bedford的模型建立[6]。具体为SD大鼠以60%~70%VO2max强度(对应跑台速度为15 m/min)运动60 min为中等强度运动;90%VO2max强度(对应跑台速度约为35 m/min)为高强度运动,高强度间歇模型为大鼠以35 m/min速度持续跑6 min,间歇时期休息5 min,以“高强度运动—间歇”模式重复3次[6,7]。
1.3 取材
EM组和EH组分别在运动结束后的即刻、6小时用2%戊巴比妥钠(0.25 ml/100 g体重)腹腔注射麻醉,C组不加以任何运动干预,在运动组第1次取材时一起取材,每小组样本量均为6只。待大鼠麻醉后腹主动脉取血,并迅速剥离大鼠睾周白色脂肪。所有白色脂肪样本均于冰上迅速切成小块,用编好号的锡纸包好置于液氮中速冻,待取材完成后统一转移至-80℃冰箱中保存。
1.4 白色脂肪组织的UCP-1和FNDC5 mRNA的相对含量的测定
1.4.1 总RNA提取和浓度测定
根据SG高纯总RNA提取试剂盒,#R1002(Sino-Gene公司)说明书要求在液氮中研磨白色脂肪组织样品,提取白色脂肪组织总RNA。用紫外分光光度计测总RNA在260 nm和280 nm处吸光度值并计算浓度。用琼脂糖凝胶检测总RNA的纯度(电泳槽JY-SPFT,电泳仪为JY300C,北京君意东方电泳设备有限公司凝胶成像系统JY04S-3C)。
1.4.2 RT-qPCR反应
根据DNaseI处理试剂(DNaseI,RNase-free,EN0521,Fermentas公司)反转录试剂盒(Thermo First cDNA Synthesis Kit,#Q1010,SinoGene公司)说明书操作逆转录过程获取cDNA。通过查询NCBI的Primer-BLAST,检索出UCP-1和FNDC5 mRNA的引物序列,所有引物由北京信诺金达生物科技有限公司合成。RT-PCR反应体系为:7.5 μl 2×SG Green qPCR Mix,0.25 μl上游引物(10 μM),0.25 μl下游引物(10 μM),1 μl cDNA,加 6 μl RNase-free ddH2O 至 15 μl。采用△△CT法(比较阈值法)表示目的基因的相对表达[8]。引物信息参见表2,PCR反映条件参见表3。

表2 引物序列
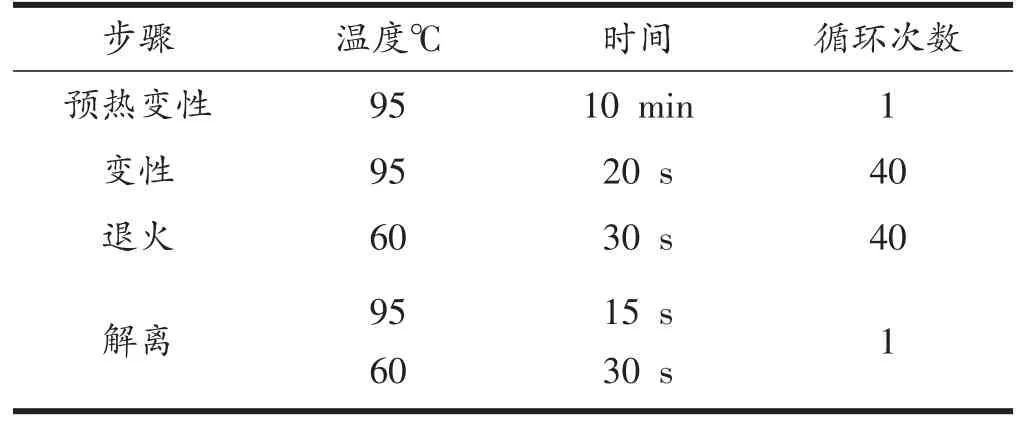
表3 RT-qPCR反应程序设置
1.5 白色脂肪组织的UCP-1和FNDC5蛋白的相对含量的测定
1.5.1 总蛋白提取及浓度检测
使用RIPA提取各组织总蛋白,用BCA蛋白浓度测定试剂盒测定蛋白的浓度,再用生理盐水统一调整至1.5 μg/μl浓度的上样量。
1.5.2 Western Blot法测试蛋白含量
12%聚丙烯酰胺凝胶电泳后将蛋白转至NC膜,根据Marker位置剪下目的条带和内参GAPDH条带,用1×TBST洗净后,封闭至5%BSA溶液中,室温孵育1 h以封闭膜上的非特异结合。孵育一抗稀释液(FNDC5,UCP-1,GAPDH浓度均1∶1000,Abcam公司)中,室温下轻摇1 h,之后4℃过夜,次日再次置于室温中孵育1 h,用TBST洗涤3次,每次用摇床摇10 min。接着孵育二抗,(FNDC5二抗为羊抗兔浓度为1:10000,UCP-1二抗为羊抗兔浓度1:5000,GAPDH二抗为羊抗鼠浓度为1:5000,Abcam公司)孵育1 h后用TBST洗涤3次,每次用摇床摇10 min。按1∶1比例将适量的ECL发光液混合后滴加到膜上于暗室曝光,根据荧光强度,选择合适的曝光时间,胶片经显影液显影后放置到定影液中,用Quantity one软件分析扫描后的图像,读取条带光密度曲线下的面积,结果用目的蛋白/内参蛋白值表示。
1.6 统计学分析
所有数据均以平均数±标准差表示。数据的统计分析使用SPSS21.0软件包。多组间数据比较分析用单因素方差分析(ANOVA),以P<0.05表示有显著性差异,P<0.01表示极具显著性差异。
2 结果
2.1 一次性运动对大鼠睾周白色脂肪FNDC5和UCP-1 mRNA表达量的影响
与C组相比,EM组大鼠睾周白色脂肪FNDC5 mRNA的相对表达量在运动后即刻和运动后6小时有上升趋势,但无显著性差异(P>0.05);与C组相比较,EH组大鼠的FNDC5 mRNA相对表达量在运动后即刻显著上升(P<0.01),与EH0组相比较,在6小时后显著下降(P<0.05)。运动后即刻,EM组的FNDC5 mRNA相对表达量显著低于EH组(P<0.05),而运动后6小时,EM组的FNDC5 mRNA相对表达量显著高于EH组(P<0.05)。见图1。
与C组相比较,EM组和EH组大鼠睾周白色脂肪的UCP-1 mRNA的表达量均在在运动后即刻显著降低(P<0.05),6小时后出现显著上升(P<0.05)。见图2。

图1 不同强度运动后睾周白色脂肪组织FNDC5 mRNA相对表达量的变化
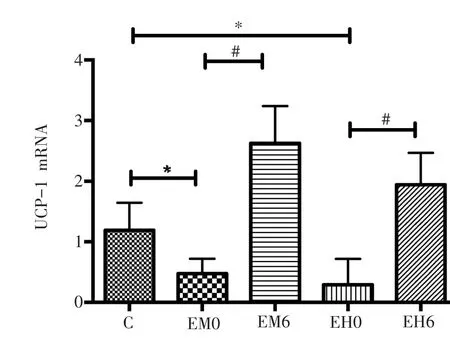
图2 不同强度运动后睾周白色脂肪组织UCP-1 mRNA相对表达量的变化
2.2 一次性运动对大鼠睾周白色脂肪FNDC5和UCP-1蛋白表达量的影响
一次性中等强度运动后,与C组相比较,EM组大鼠睾周白色脂肪FNDC5蛋白表达量持续升高(P<0.05);EH组大鼠睾周白色脂肪FNDC5蛋白相对表达量有先升高在降低的趋势,但无显著性差异(P>0.05)。运动后6小时,EH组的FNDC5蛋白相对表达量显著低于EM组(P<0.01)。
与C组相比较,EM 组大鼠睾周白色脂肪UCP-1蛋白相对表达量呈有上升趋势,但无显著性差异(P>0.05);EH组大鼠睾周白色脂肪UCP-1蛋白相对表达量在运动后即刻显著升高(P<0.05),到运动后6小时有降低的趋势,但与EH0组相比较无显著性差异(P>0.05)。
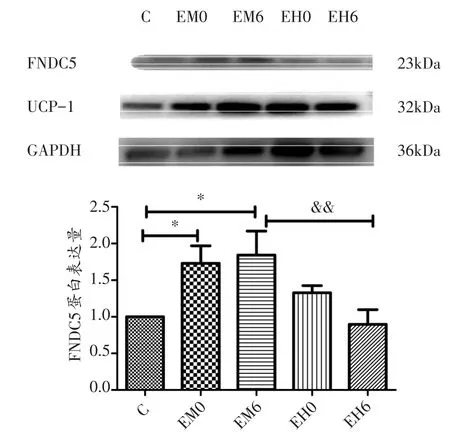
图3 不同强度运动后睾周白色脂肪组织FNDC5蛋白相对表达量的变化

图4 不同强度运动后睾周白色脂肪组织UCP-1蛋白相对表达量的变化
3 讨论
本研究观察到,一次性中等强度运动和高强度间歇运动均可引起大鼠睾周白色脂肪FNDC5、UCP-1 mRNA和蛋白表达量的变化,但在作用持续时间上具有较大的差异。一次性中等强度运动后大鼠睾周白色脂肪FNDC5 mRNA表达量有持续增加的趋势,但无显著性差异;FNDC5蛋白表达量在运动后持续增加。而一次性高强度间歇训练后即刻大鼠睾周白色脂肪FNDC5 mRNA表达量较运动前就有非常显著的升高,但在6小时后显著减低,甚至低于运动前水平;FNDC5蛋白表达量有相似的变化趋势,但无显著差异。一次性中等强度运动和高强度间歇运动均可使睾周白色脂肪UCP1 mRNA的表达量在运动后即刻显著降低,但在6小时后又显著增加。一次性中等强度运动使睾周白色脂肪UCP-1蛋白表达量有增加的趋势,但无显著差异;而一次高强度间歇运动后即刻UCP-1蛋白表达量显著增加。
3.1 一次性运动对大鼠睾周白色脂肪FNDC5表达量的影响
本研究中,一次性运动能够诱导E组大鼠FNDC5的表达量上升。Irisin和其前体物质FNDC5在脂肪组织中是反映白色脂肪出现棕色化的标志性因子,因此可以通过Irisin和FNDC5来反映脂类代谢和能量消耗。说明本研究中一次性运动能够诱导白色脂肪棕色化,促进大鼠睾周白色脂肪能量消耗增加。Jedrychowski等研究显示人体中存在Irisin,运动能够刺激其表达水平升高[9]。而与正常人相比较,肥胖和II型糖尿病人身上FNDC5蛋白表达量和血清Irisin表达量显著下降[10-13]。有研究显示,运动作为激活PGC-1α/FNDC5途径的因素之一,能够促进FNDC5/Irisin的分泌和释放[14-16],由FNDC5蛋白剪切而来的Irisin受到PGC-1α的调节,在脂肪组织中能够提高脂肪代谢,增加能量消耗[17]。受 r-Irisin(recombinant Iirisin)干预(3500 mg/kg/week)的肥胖小鼠能够刺激其白色脂肪组织出现“棕色化”,同时使UCP-1表达量显著升高(>25倍),并且体重减轻[18]。除了脂肪组织,在肌肉中Irisin和肌生成抑制蛋白呈负相关[19],在敲除肌生成抑制蛋白基因表达的小鼠骨骼肌中,Irisin和其前体物质FNDC5的表达量极高,因此可以通过Irisin和FNDC5观察肌肉质量[20]。利用r-Irisin干预的小鼠后发现FNDC5细胞表达数量显著升高,提示增加Irisin会导致其前体FNDC5的增加,即Irisin和FNDC5的表达有一致性[21,22]。
本研究结果显示,中等强度运动刺激后FNDC5蛋白表达量在运动后6小时内持续升高。有研究显示,以65%VO2max强度进行10周训练会使健康成年男性血清Irisin水平增加两倍[23]。以中等强度进行1小时自行车运动后,人体中循环Irisin水平增加了3倍[24,25]。在本研究中,尤其是EH组,在一次性运动后即刻FNDC5 mRNA显著上升,并在6小时后显著下降。说明在一次性高强度间歇运动后即刻FNDC5变化较明显,提示FNDC5 mRNA对高强度间歇运动的刺激更加敏感,在运动后即刻出现一过性的升高。一次性低强度间歇运动(LIE)和高强度间歇运动(HIE)后6小时HIE组的血清Irisin浓度上升18%,在相同的能量消耗下高强度间歇运动组的血清Irisin水平显著高于低强度间歇运动组[24]。外源性注射Irisin的干预剂量会对白色脂肪棕色化的效果产生影响,低剂量(100 mg/kg/week)的r-Irisin干预不能够使小鼠出现“棕色化”[26],但能够加强骨骼中FNDC5 mRNA的表达。2015年Qiu等[27]通过meta分析发现耐力型运动对Irisin的浓度并没有显著的影响,得出这一结论的可能原因是作者没有充分考虑运动后测试Irisin浓度的时间点。Irisin分子存在降解率高、寿命较短的特点[28]。
3.2 一次性运动对大鼠睾周白色脂肪UCP-1表达量的影响
本研究结果显示,一次性运动能够使大鼠睾周白色脂肪组织UCP-1表达量升高,提示一次性运动能够刺激白色脂肪组织产热水平增加。UCP-1是位于线粒体内膜产热的效应物质,主要在“棕色化”的白色脂肪组织或棕色脂肪中通过线粒体来发挥产热和耗能的作用[29,30]。在白色脂肪组织中,诱导其出现棕色化的途径有很多,其中包括激活代表棕色脂肪的重要基因——UCP-1[31],UCP-1作为脂肪组织中产热和耗能的标志性蛋白,可以作为检验脂肪组织中产热和耗能是否活跃的指标。许多研究显示,刺激白色脂肪组织中UCP-1的表达量升高和促进白色脂肪“棕色化”,在动物模型中能够逆转遗传性肥胖[32,33]。
在本研究中,我们观察到在一次性运动后即刻UCP-1 mRNA表达量均显著下降,运动后6小时显著上升,提示运动能够诱导白色脂肪组织产热水平增加。运动能够诱导高脂饮食小鼠白色脂肪组织中线粒体酶和棕色脂肪中UCP-1蛋白含量增加。在白色脂肪细胞中,Irisin通过p38MAPK途径激活UCP-1的表达[34],伴随白色脂肪组织UCP-1表达的增加和棕色脂肪组织中UCP-1活性增强,从而激活棕色脂肪组织中线粒体的产热作用,促进脂肪的燃烧。本研究通过RT-qPCR方法测定组织中UCP-1 mRNA表达量发现睾周白色脂肪组织中UCP-1受到运动刺激所产生的反映具有时效性,其发挥效应的时间在运动后6小时。有研究显示,在培养的脂肪细胞中添加FNDC5会导致多腔室脂滴中的UCP-1阳性脂肪细胞显著性增加,同时UCP-1蛋白表达增加[35]。以40%VO2max强度运动后,皮下白色脂肪组织中Irisin蛋白表达增加,从而促进白色脂肪组织向棕色脂肪组织转变和提高棕色脂肪样因子UCP-1蛋白表达增加[36]。说明脂肪组织中白色脂肪棕色化水平的增加会伴随着UCP-1的表达量的增加,FNDC5和UCP-1呈正相关。在本研究中,一次性运动能够诱导UCP-1蛋白水平增加,尤其是在一次性高强度间歇运动刺激后即刻,UCP-1蛋白表达水平出现显著性上升,并在运动后6小时内持续发挥效能,产热作用持续升高。说明一次性运动能够诱导白色脂肪产热能力增加。
在本研究中,FNDC5和UCP-1受不同强度运动刺激的影响后变化的波动情况略有差异,一次性运动后UCP-1 mRNA的变化情况并未同FNDC5 mRNA的变化情况一致。在本研究中,FNDC5需要一次性的大强度运动刺激后即刻才会诱导其显著性增加,而UCP-1 mRNA的表达水平并没有在一次性运动后马上升高,而是在运动后即刻显著性降低,在6小时后又会出现显著性升高,即运动后6小时产热水平增加。从蛋白表达角度看,一次性运动能够诱导FNDC5和UCP-1蛋白表达量上升,即一过性的运动能够使白色脂肪组织出现“棕色化”,组织中产热上升。在本研究中,EM6组FNDC5的表达量显著高于EH6组,提示FNDC5蛋白对中等强度运动的刺激较敏感,而UCP-1对高强度间歇的运动刺激更为敏感。
在脂肪组织中,运动诱导白色脂肪棕色化并不止存在PGC-1α—FNDC5—UCP-1这样一条通路。机体中还有其他因子和途径参与白色脂肪中棕色脂肪细胞的增殖和分泌。白色脂肪棕色化过程涉及许多转录因子的表达,其中包括PPAR家族[37],PRD1-BF-1-RIZ1同源蛋白如PRDM16[38]等,BMP-7的升高也能够刺激UCP-1表达量上升[39],肌生成抑制蛋白(Mtn)也能通过miR-34a调控FNDC5,参与调控白色脂肪细胞棕色化[40]。激活TRPV1诱导SIRT-1表达增加,也能够刺激脂肪组织产热增加[41],参与脂肪的产热。所有PPAR的亚型(α,β,γ)都与UCP-1的转录有关[42,43],激活PPARγ能够诱导UCP-1表达增加[44],但是也有研究显示,在棕色脂肪细胞的分化过程中,当PPARγ基因表达没有出现上升、甚至下降的情况下,UCP-1的表达量也有可能出现上升,提示UCP-1的基因表达与其上游因子不止存在简单效应[45]。在这一过程中,激活PPAR-α激动剂,肾上腺激素受体、甲状腺激素受体,Irisin和FGF21等均有可能诱导脂肪组织中出现“棕色化”现象[37]。
除了蛋白和基因对组织中产热功能的影响外,外周的甲状腺激素和瘦素等激素也会对机体的产热功能产生影响[46]。运动过程中和运动后机体的激素水平会发生改变。运动可以促进大脑释放去甲肾上腺素,心脏(心肌)释放利钠肽,也可以促进骨骼肌释放PGC-lα,PGC-lα能够促进血液中 Irisin、氨基异丁酸(BABAβ)和成纤维细胞生长因子21(FGF21)等生成。机体交感神经系统通过去甲肾上腺素中枢调节生热作用,其作用于肾上腺素能受体,也可能导致以急性和/或长期方式激活产热[37]。因此,在一次性运动后虽然能够诱导大鼠睾周白色脂肪组织产热耗能增加,但是有可能不是完全依赖FNDC5—UCP-1激活白色脂肪组织产热水平上升。
4 结论
(1)一次性运动能够提高大鼠睾周白色脂肪FNDC5和UCP-1蛋白表达水平,促进白色脂肪棕色化,增加脂肪组织产热。
(2)一次性运动后FNDC5mRNA和UCP-1mRNA的变化具有时效性,其中UCP-1mRNA在运动后6小时出现显著升高,而FNDC5mRNA在高强度间歇运动后即刻变化最明显。
