5种气候生态型割手密F1和F2杂种的耐旱性评价
2017-11-28田春艳桃联安俞华先董立华经艳芬边芯郎荣斌周清明安汝东孙有芳杨李和
田春艳,桃联安,俞华先,董立华,经艳芬,边芯,郎荣斌,周清明,安汝东,孙有芳,杨李和
5种气候生态型割手密F1和F2杂种的耐旱性评价
田春艳,桃联安,俞华先,董立华,经艳芬,边芯,郎荣斌,周清明,安汝东,孙有芳,杨李和
(云南省农业科学院甘蔗研究所瑞丽育种站,云南瑞丽678600)
干旱是甘蔗生产中重要的逆境因子,严重影响甘蔗产量和品质。甘蔗野生资源割手密具有抗逆性强、宿根性强、适应性广等特点,研究其后代耐旱性,筛选耐旱种质,为培育新型抗旱亲本、实现甘蔗新品种突破提供理论依据。选取5种生态类型割手密与云瑞系列创新亲本、国外种以及国内种的杂交F1、F2代材料共36份,采用温室人工控水胁迫的方法,于甘蔗拔节初期设置正常供水(作对照)、轻度胁迫和重度胁迫3个处理。分别测定3个处理条件下甘蔗叶片质膜透性(plasma membrane permeability,PMP)、丙二醛(malondialdehyde,MDA)、叶绿素(chlorophyll,CHL)、可溶性蛋白(soluble protein,Pr)、脯氨酸(proline,Pro)含量和超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)酶活性7个生理生化指标,利用模糊隶属函数法、聚类分析和灰色关联度分析,综合评价割手密后代在不同程度水分胁迫下的耐旱性表现。水分胁迫下甘蔗叶片的PMP、MDA含量、Pro含量、SOD和POD酶活性升高,CHL含量和Pr含量降低,变化幅度因甘蔗基因型及水分胁迫程度差异而不同;隶属函数分析结果表明,不同程度水分胁迫下,割手密后代的耐旱能力不同,F1代耐旱性较强,综合抗旱能力超过双亲的个体比例为82%,远高于F2代;聚类分析表明,所有材料在轻度水分胁迫下可分为4大类群,其中,Ⅰ和Ⅱ类耐旱性最好,Ⅳ类次之,Ⅲ类耐旱性最差且全为F2代材料,F1代均分布于耐旱性较好的Ⅰ、Ⅱ、Ⅳ类群。重度胁迫下可分为3大类群,Ⅰ、Ⅲ类耐旱性较强,Ⅱ类耐旱性最差,F1代(除云割F108-473外)均分布在耐旱性较强的Ⅰ、Ⅲ类群;灰色关联度分析表明,不同程度水分胁迫下,各指标与耐旱性间的关联度不同。轻度胁迫下,关联度依次为POD>SOD>PMP>MDA>Pr>CHL>Pro;重度胁迫下关联度依次为SOD>Pr>CHL>POD>PMP>MDA>Pro。36份割手密F1和F2杂种材料中,云割F108-536、云割F211-179、云割F211-8、云割F108-474、云割F108-398、云割F108-254、云割F211-261、云割F211-151、云割F108-538、云割F108-391、云割F211-188、云割F211-37、云割F107-150、云割F211-258和云割F211-90耐旱性较强,为强耐旱种质,可在甘蔗抗旱亲本培育及抗旱育种中加以重点利用;割手密耐旱特性在不同世代中的遗传表现不同,F1代耐旱性强于F2代;割手密后代耐旱性强弱与其父本割手密的气候生态类型、地理位置及海拔无明显相关性;PMP、MDA含量、CHL含量、SOD和POD酶活性5个生理生化指标与甘蔗耐旱性关联度较高,是耐旱性评价的优良指标。
割手密;水分胁迫;生理生化指标;耐旱性;甘蔗
0 引言
【研究意义】甘蔗是热带和亚热带地区的重要经济作物,也是中国最重要的糖料作物,蔗糖产量占中国食糖总产量的92%以上[1]。中国蔗区主要分布在广西、云南和广东等地,其中旱地植蔗面积达85%以上。作为全国第二大甘蔗主产区的云南,甘蔗多种植在旱坡地,生长条件恶劣,且自然干旱经常发生,水分供给问题严重。此外,现有主栽品种随着种植时间延长会出现种性退化,产量和品质下降等问题,急需扩宽杂交亲本野生种血缘,实现亲本抗逆性突破,提高新育成品种的生活力、宿根性和适应性[2]。割手密(L.)为复杂多倍体或非整倍性植物,是甘蔗属及其近缘属植物中最有研究价值和育种价值的野生种之一[3]。具有抗逆性强、宿根性好、适应性广等特点,并且割手密资源数量庞大,生态变异类型多,遗传多样性丰富。研究割手密血缘后代对水分胁迫的响应,筛选抗旱种质,对实现抗旱甘蔗亲本创新及培育突破性甘蔗新品种尤为重要。【前人研究进展】干旱是作物生产中重要的逆境因子,严重影响作物的生长速率和发育[4]。干旱胁迫下,植物细胞遭到破坏,体内氧自由基种类增多,严重损害植物细胞、蛋白质和核酸结构,直接导致作物减产、品质下降等严重后果[5-6]。罗俊等[7-8]先后研究了叶绿体荧光参数及活性氧代谢、MDA含量和膜透性指标与甘蔗品种耐旱性及甘蔗苗期抗旱性的关系。结果表明,荧光参数的受抑程度与品种的耐旱性相对强弱密切相关,荧光参数的动态变化可作为简便易行的抗旱性强弱诊断指标在甘蔗抗旱育种上应用。Zhao等[9]研究水分亏缺下甘蔗叶片含水量、绿叶面积以及光合作用等方面的变化,说明土壤水分亏缺对甘蔗早期的生长、分蘖均有很大的影响;SILVA等[10]、BASNAYAKE等[11]采用干旱胁迫与灌溉的处理方式对甘蔗的抗旱性进行评价,结果表明干旱条件下,甘蔗减产明显,且不同基因型间差异显著。许文花等[12]、罗明珠等[13]测定了干旱胁迫下割手密叶片的细胞质膜透性、脯氨酸、丙二醛和可溶性糖含量、根系活力等一系列生理生化指标,并利用模糊隶属函数法、方差分析等对不同割手密无性系、不同抗旱性甘蔗品种进行抗旱性评价,表明不同基因型割手密抗旱性差异显著,并认为叶片脯氨酸含量和质膜透性可作为甘蔗抗旱性评价指标。以上生理生化指标体系皆从水分亏缺引起的植物体内系列生理生化反应出发,从不同代谢途径及角度对植物耐旱性进行分析评价,为植物抗旱性强弱评价体系提供了多种途径及评价指标。并且经研究证明这些指标在其他植物或作物的抗旱性鉴定上同样可靠、具有重要的研究、利用价值[14-16]。在分析方法上,桃联安等[17]、边芯等[18]分别利用抗旱隶属函数法、主成分分析法分析了云南割手密血缘F1、F2材料,筛选出抗旱性强的材料,为甘蔗抗旱育种提供亲本材料和选择依据。高三基等[19]利用相关、主成分、聚类和判别分析方法,研究了有关生理生化指标与甘蔗抗旱性的关系,结果表明,丙二醛、质膜透性等是甘蔗抗旱性综合鉴定的可靠指标,可在甘蔗抗旱性育种上加以应用。【本研究切入点】在甘蔗育种实践中,野生资源因具有强抗逆性、强宿根性以及适应性广等优点越来越受到重视。而在甘蔗新品种选育的漫长过程中,后期对品系的选择主要依据产量和糖分,对抗旱性的选择效率极低[20-22]。云南地理环境独特,气候类型多样,具有丰富的野生割手密资源,但其利用效率极低,仅有少数得到应用或利用成功[23-25]。严重阻碍了含割手密血缘的创新亲本种质在甘蔗育种中的应用。【拟解决的关键问题】本研究以甘蔗与5种气候生态型割手密杂交所获得的F1、F2后代及双亲为材料,测定其水分胁迫下的系列生理生化指标,旨在分析割手密后代对不同程度水分胁迫的响应及其耐旱性,筛选耐旱种质,为充分利用云南割手密培育耐旱亲本及进行甘蔗抗旱育种提供依据和指导。
1 材料与方法
1.1 试验材料
5种气候生态类型的割手密与云瑞系列创新亲本(含割手密血缘)、国外种以及国内种的杂交F1、F2代材料,其中,云割F1代材料12份,为6个组合的后代,云割F2代23份均为云瑞09-525×云割F108-254的杂交后代,所有供试材料共36份,F1代材料及其割手密父本信息详见电子附表1。
1.2 试验设计与生理生化指标测定
试验在云南省农业科学院甘蔗研究所瑞丽育种站进行,干旱胁迫在温室大棚内进行。试验材料于2015年3月6日进行单芽桶栽种植,每个材料种植9桶,每桶5个芽,正常浇水管理使其正常生长。于2015年9月22日(拔节初期)开始把材料分成3组,每组3桶。第1组继续正常供水,作为对照;第2、3组停止供水,进行水分胁迫处理,其中第2组作轻度水分胁迫,第3组作重度水分胁迫。1周后,处理组部分材料开始呈轻微萎蔫状态,梢部叶片略有枯黄、卷曲等轻度缺水症状,此时于早晨8:00开始取第1组、第2组材料,进行各抗旱性生理生化指标的测定;第3组继续胁迫,待胁迫15 d后,第3组较第1组(对照组)三分之一以上材料出现永久萎蔫、叶片枯黄卷曲、少部分甚至枯死等严重缺水症状时进行重度胁迫抗旱指标的测定,同时再次测定对照组材料。
参照谭常[26]的方法测定质膜透性(PMP),参照李忠光等[27]方法测定丙二醛(MDA)、脯氨酸(Pro)、可溶性蛋白(Pr)含量以及过氧化物酶(SOD)、超氧化物歧化酶(POD)活性,采用SPAD叶绿素仪测定叶绿素(CHL)含量。其中,PMP、MDA、Pr 和Pro的测定用+1叶,SOD和POD酶活性的测定用心叶,CHL含量的测定连续测定了+1—+4叶。取样时从每桶中随机选择1株,共3叶,测定时将所取的3叶叶片去主脉,剪碎混合后进行称量和后续研磨测定。
1.3 统计分析
采用Microsoft excel、DPS ver 16.05进行数据的整理和统计分析。根据模糊隶属函数法,参照张保青等[28]方法计算轻、重度水分胁迫下各生理生化指标的隶属函数值,并分别计算其轻度胁迫和重度胁迫隶属度,表示其在轻度水分胁迫、重度水分胁迫下的耐旱能力,最后计算其综合隶属度(即,轻度胁迫和重度胁迫隶属度的平均值),综合分析、评价参试材料在水分胁迫下的耐旱性。
隶属函数值计算公式如下:
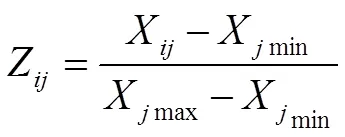
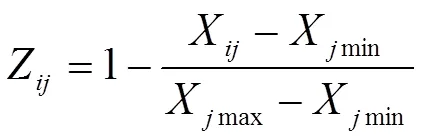
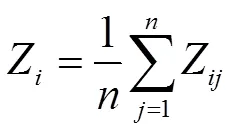
式中,Z表示基因型指标的抗旱隶属函数值,X表示基因型指标的测定值,Xmin、Xmax分别表示基因型中所测定指标的最小值和最大值。计算各指标隶属函数值时,如果某一指标与抗旱性呈正相关,用(1)式计算;若与抗旱性呈负相关,则用(2)式进行反转;(3)式表示基因型各指标的抗旱隶属函数平均值(即,抗旱隶属度),为指标数。
采用卡方距离(chi-square measure)、离差平方和法(sums of Squared Deviations)对7个生理生化指标数据进行系统聚类分析。
2 结果
2.1 割手密后代对水分胁迫的生理响应
水分胁迫下,甘蔗叶片各生理生化指标的变化幅度(表1),即各指标在水分胁迫处理下较正常供水(对照)的增减变化情况,表示植物体内各生理生化指对水分亏缺的生理响应及响应灵敏度,反映植物对水分亏缺是否敏感。水分胁迫下,甘蔗叶片的PMP、MDA含量、Pro含量、SOD酶活性和POD酶活性升高,CHL含量和Pr含量降低,变化幅度因甘蔗基因型及水分胁迫差异而不同。SOD和POD酶活性的变幅为轻度胁迫大于重度胁迫,其余5个指标的变幅皆为重度胁迫大于轻度胁迫。方差分析表明,轻度胁迫与重度胁迫间(=0.5142>0.05)差异不显著。重度水分胁迫下,除处理与对照的CHL(=0.6371>0.05)和Pr含量(=0.0654>0.05)差异不显著外,其余5个指标均达到显著水平(<0.05),其中,PMP(=0.0093<0.01)达极显著水平。
重度水分胁迫下,云割F211-253、云割F211-74、云割F211-43、云割F211-169、云割F211-122、云割F211-19、云割F211-229、云割F211-254、云割F211-162、云割F208-392、云割F211-18、云割F208-473首先表现严重缺水症状,部分材料枯死,其生理生化指标测定为PMP(100.4%—292.8%)和MDA(47.55%— 331.0%)增幅较大,叶绿素、可溶性蛋白质降解严重,同时SOD酶活性(云割F211-74增幅最小,仅为4.4%)、POD酶活性(云割F211-169,4.6%)增幅较小,未能有效缓解干旱给膜系统带来的伤害,渗透调节物质脯氨酸含量(云割F211-253,增幅仅为6.5%)也较低,这些材料耐旱能力较弱,与植株缺水症状表现一致,其中以F2代材料(约占83%)居多。
2.2 隶属函数分析
隶属函数结果(表2)表明,参试材料在轻度、重度水分胁迫下的隶属度及排序不同。部分材料如云割F108-543、云割F211-165、云割F211-192、云割F108-392、云割F211-229、云割F211-122、云割F211-43、云割F211-253在轻度水分胁迫下隶属值较大,表现较好的耐旱性,但在重度水分胁迫下隶属值较小,耐旱性很差;云割F211-179、云割F108-474、云割F108-538、云割F211-248和云割F211-56则正好相反。但重度胁迫下,胁迫时间较长,植物响应水分亏缺更充分,生理生化指标变化明显,测定结果与植物对水分亏缺引起的严重缺水症状表现一致,耐旱性评价结果更可靠。且方差分析表明,轻度胁迫与重度胁迫间差异不显著,故表2最终仅以重度胁迫下耐旱隶属值排序表示供试材料耐旱性。
参试材料中,割手密F1代较F2代表现更强的耐旱性。在F2代材料(23份)中,耐旱能力超过双亲(云割F108-254、云瑞09-525)的有云割F211-179、云割F211-8、云割F211-261、云割F211-151、云割F211-188、云割F211-37、云割F211-258、云割F211-90、云割F211-248、云割F211-56和云割F211-202,共11份,约占总数的48%;F1代材料(11份,除云割F108-254)中,耐旱能力超过上述亲本的有云割F108-536、云割F108-474、云割F108-398、云割F108-391、云割F108-543、云割F107-150、云割F107-152和云割F108-538共8份,约73%,所占比例远高于F2,这些材料皆具有较强的耐旱性。其中耐旱能力前15位依次为云割F108-536、云割F211-179、云割F211-8、云割F108-474、云割F108-398、云割F108-254、云割F211-261、云割F211-151、云割F108-538、云割F108-391、云割F211-188、云割F211-37、云割F107-150、云割F211-258和云割F211-90,可在甘蔗育种中优先利用。
根据隶属函数分析结果,5种气候生态型割手密后代F1耐旱性比较分析结果(表3)表明,耐旱性与其父本割手密的气候生态类型、地理位置以及海拔高度之间并无直接相关性。本研究中耐旱性最强的为低海拔、北热带半湿润半干燥型生态型的云割F108-536,与其具有相同生态型的云割F108-538和云割F108-543耐旱性排名分别为第5和第9,其耐旱性与父本割手密的气候生态类型、地理位置以及海拔高度之间无明显相关性。
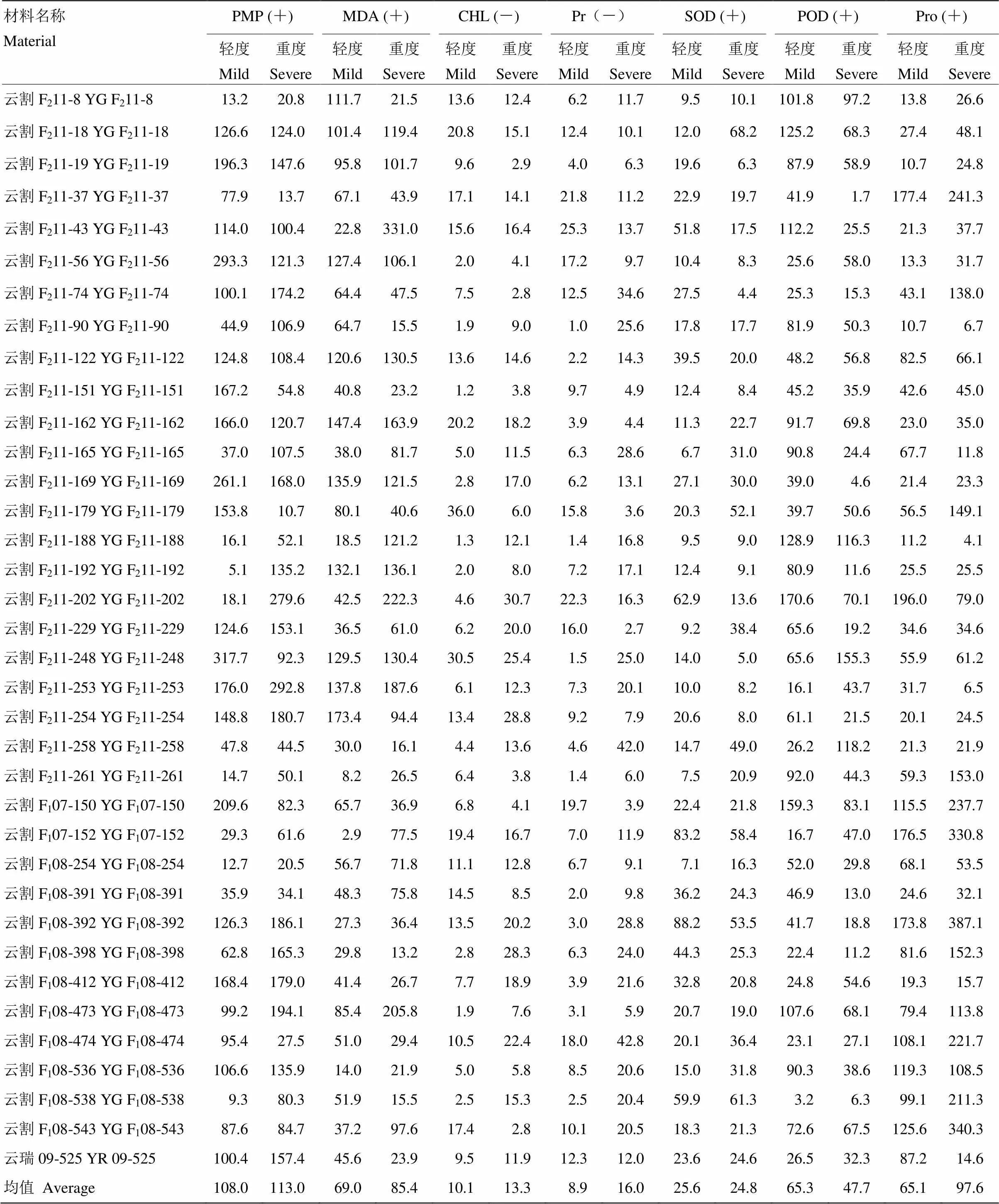
表1 水分胁迫下甘蔗叶片各生理生化指标的变化幅度
表内数值为研究材料在水分胁迫下各生理指标的变化幅度(%)=(水分胁迫下各生理指标测定值-正常供水的各生理指标测定值)÷ 正常供水的各生理指标测定值×100%;(+):表示增加;(-):表示减少
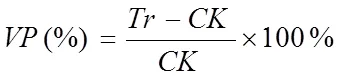
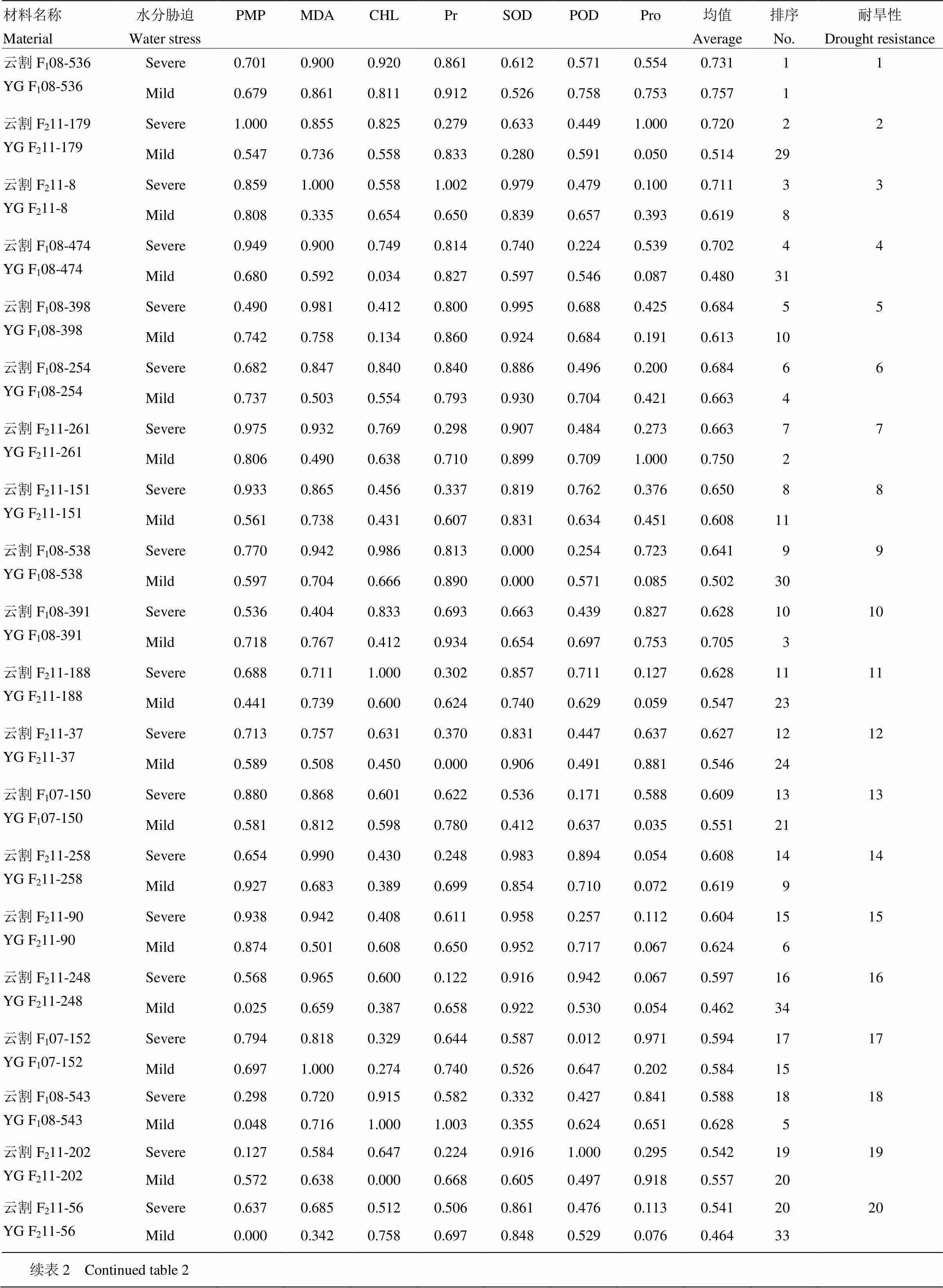
表2 生理生化指标隶属函数值及材料耐旱性
2.3 聚类分析
割手密后代7个生理生化指标的聚类分析(图1)表明,所有参试材料(共36份)在轻度水分胁迫下可分为4大类群(图1-A,Ⅰ、Ⅱ、Ⅲ、Ⅳ),其中Ⅰ类有9份,Ⅱ类10份、Ⅲ类8份、Ⅳ类8份;在重度胁迫下可分为3大类群(图1-B,Ⅰ、Ⅱ、Ⅲ),Ⅰ类有8份,Ⅱ类16份、Ⅲ类12份,此分类结果与隶属函数分析方法所得结果基本一致,仅有极少数材料存在偏差。轻度胁迫(图1-A)下,Ⅰ和Ⅱ类材料耐旱性最好,Ⅳ类次之,Ⅲ类耐旱性最差,且Ⅲ类全为割手密F2代材料,所有F1代均分布于耐旱性较好的Ⅰ、Ⅱ、Ⅳ类群中。但Ⅰ类中的云割F211-188;Ⅱ类中的云割F211-37、云割F108-474、云割F108-538;Ⅳ类中的云割F211-74、云割F211-179、云割F108-412,这些材料在隶属函数分析中耐旱性较差,与生理生化指标的聚类结果不一致。重度胁迫(图1-B)下,Ⅰ、Ⅲ类的耐旱性较强,Ⅱ类耐旱性最差,割手密F1代材料(除云割F108-473外)均分布在耐旱性较强的Ⅰ、Ⅲ类群里,说明割手密F1代较F2代具有更强的耐旱能力。但Ⅲ类的云割F211-74和云割F208-392在重度胁迫的隶属函数分析中耐旱性较差,Ⅱ类的云割F211-188、云割F211-248和云割F211-202耐旱性却相对较强,这些个别材料的耐旱性鉴定结果在2种评价方法中存在差异。
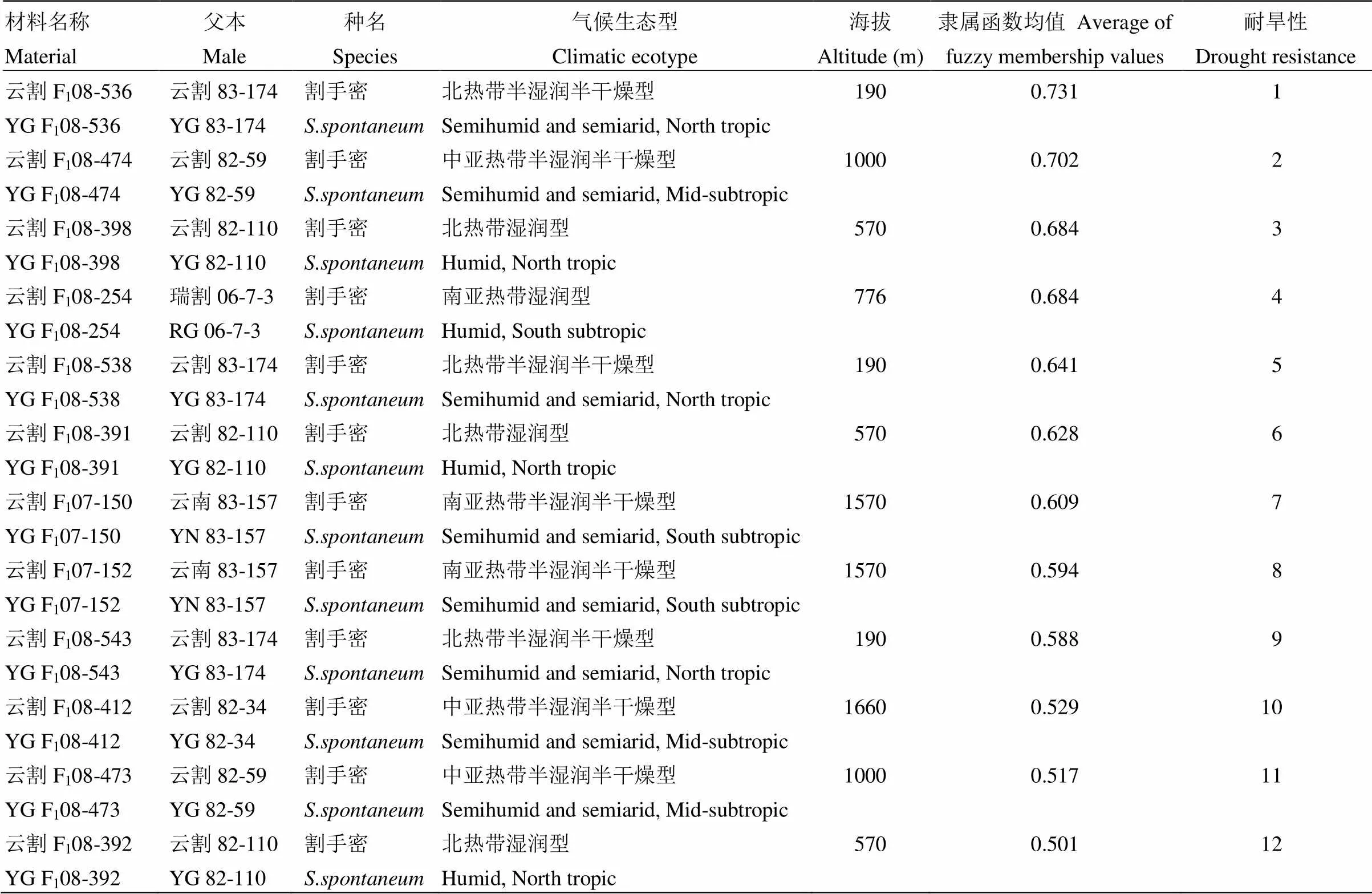
表3 5种气候生态型父本割手密F1材料耐旱性
对于父本割手密生态型与其F1材料的聚类分析显示,相同生态型父本割手密的F1材料并未完全聚在同一类群,而是分散在各个类群,同样表明后代耐旱性与父本割手密的气候生态类型、地理位置以及海拔高度之间无直接关系,与隶属函数分析结果一致。
2.4 灰色关联度分析
以模糊隶属函数分析法所得的耐旱性隶属度为母序列,7个生理生化指标变化幅度为子序列进行灰色关联度分析,结果表明,不同程度水分胁迫下,各指标与耐旱性的关联度不一致,即在轻度、重度水分胁迫下,各指标对水分亏缺的敏感程度及植株的耐旱机制不同。轻度水分胁迫下,各指标与甘蔗耐旱性间的关联密切程度依次为POD>SOD>PMP>MDA>Pr>CHL>Pro,关联系数分别为0.4323、0.4180、0.3971、0.3840、0.3795、0.3725和0.3721;重度胁迫下,其关联度依次为SOD>Pr>CHL>POD>PMP>MDA>Pro,关联系数分别为0.4766、0.4684、0.4562、0.4150、0.3939、0.3761和0.3413。脯氨酸在轻度、重度水分胁迫下皆与甘蔗耐旱性关联度最低。
3 讨论
3.1 割手密后代对水分胁迫的生理生化响应及耐旱性
水分亏缺时,甘蔗叶片失水,细胞膜脂的过氧化作用加剧,细胞质膜透性增大,MDA大量积累。同时,水分胁迫下,甘蔗叶片内叶绿素和可溶性蛋白质降解,体内的渗透调节物质脯氨酸等的积累,增加了细胞质浓度,降低渗透势,保持水分和各生理活动正常进行,缓解细胞脱水伤害;其植物体内的氧自由基含量及种类也会大量增多,此时SOD和POD酶活性增强,能清除氧活性自由基,延缓细胞衰老,有助于植株抵御干旱伤害,表现耐旱性。但不同程度水分胁迫下,各指标对水分亏缺的响应敏感度及不同基因型甘蔗在不同胁迫程度下的抗旱机制可能不同。其适应和抵御干旱的途径不是单一的,而是多种多样的[29]。

A:轻度水分胁迫下耐旱性聚类图;B:重度水分胁迫下耐旱性聚类图;分枝上数值为耐旱性排名
本研究中云割F108-474和云割F108-538在轻度胁迫下耐旱性较差,而在重度水分胁迫下却表现出很强耐旱性;云割F211-165、云割F211-253、云割F108-392在轻度胁迫下耐旱性较强,而在重度水分胁迫下却表现不耐旱或耐旱性极弱。这与白向历等[28]对玉米的抗旱性研究结果一致,即干旱胁迫程度不同,抗旱评价结果也会有偏差。当然这并不能排除可能是试验误差的影响,这在其他作物的抗旱性评价中也得到了研究证实[30-31]。总之,重度胁迫下,胁迫时间充足,植物对水分亏缺的生理响应充分,植物体内生理生化反应也渐渐趋于稳定,测定值的耐旱评价结果与所观察到的植物因水分亏缺引起的缺水症状表型结果一致,能客观反映植物耐旱性,结果更可靠。
本研究基于隶属函数分析和聚类分析,针对来源于5种不同生态型父本割手密的F1材料耐旱性评价结果表明,割手密后代耐旱性强弱与其父本割手密的气候生态类型、地理位置以及海拔高度之间并无直接相关性,这与本单位前期对其他创新种质抗旱性相关性状的主成分分析研究的结果一致[18]。这有可能是所选的父本割手密生态型差异较小、代表性不强和数量有限导致的,两者之间是否存在明确的关联性还有待进一步研究探讨。朱建荣等[32]在云南不同生态型甘蔗细茎野生种育种潜力分析中指出,不同生态型割手密(父本)对其杂交后代主要经济性状的改良主要表现为产量性状的变化,不同生态型割手密无性系的蔗茎产量、蔗糖分遗传潜力表现不尽相同,应有目的地选择利用多种类型的无性系创新种质,培育新型亲本,丰富甘蔗育种遗传基础。
3.2 割手密不同世代间耐旱性差异
割手密为复杂的异源多倍体植物,遗传背景复杂,杂交后代分离广泛,基因型间差异大。且耐旱性为多基因控制的数量性状,因此,准确评价甘蔗材料耐旱性非常困难。对亲本及其后代抗旱性的评价结果只能表明其作为亲本产生高抗旱性后代的一种趋势和可能性[2]。另外甘蔗在生长过程中受自身生长特性、群体对个体的影响及其与环境的互作等众多因素的影响。本研究针对7个抗旱指标的模糊隶属函数分析及聚类分析结果共同表明,割手密血缘后代中,F1代的抗旱性明显强于F2代,如在模糊隶属函数分析中,抗旱能力超过双亲的,F1代约占82%,F2代仅占48%,其中抗旱性最强的材料就是F1代的云割F108-536。此现象可能是由于随着杂交世代的增加,含抗旱性基因的割手密染色体不断减少,耐旱性逐渐减弱甚至消失。现有研究结果表明,异源多倍体植物中基因表达的变化主要包括亲本基因的沉默(silencing)、基因的激活(activation)以及部分同源基因(homoeologous gene)表达水平的变化[33-36]。因此,对于甘蔗这样的异源多倍体植物,准确分析评价其抗旱性显得更加复杂、困难,不仅要求在进行抗旱性评价时既要选用多指标,又要利用多种评价方法相互验证,必要时还需结合分子技术进行深入的分析研究,提高准确性,为抗旱育种提供正确的指导及可靠的依据。
3.3 甘蔗耐旱性评价指标及体系的选择
植物耐旱性是多基因控制的数量性状,同时受基因型差异和个体与环境互作的影响很大。目前对植物耐旱性的研究常采用多指标评价体系,从形态指标[37]、生理生化指标[13,29]等方面利用模糊隶属函数法[38]、主成分分析法[18]、聚类分析[23]、逐步判别分析法[2]、灰色关联度分析法[39]、相关分析[40]等进行综合评价。
本研究在前人对耐旱性研究的基础上,选取7个生理生化指标,其结果证明PMP、MDA、SOD酶活性和POD酶活性这4个指标与甘蔗耐旱性紧密相关,对干旱胁迫反应灵敏,在胁迫下均表现为含量或活性增加,且在胁迫初期大量增加,随着胁迫程度加大,增量变缓。且水分胁迫的程度不同,指标与耐旱性隶属度的关联度也不完全相同,这说明各指标对水分亏缺的灵敏度是有差异的。同时,本研究采用了模糊隶属函数分析、聚类分析和灰色关联度分析3种方法,对割手密F1、F2代在水分胁迫下的耐旱性进行了评价、分类聚类、同时分析了7个生理生化指标与耐旱性间的关联密切程度,得到了相对理想的结果,表明所选生理生化指标可靠、分析方法间形成相互验证,结果更加可靠。不少研究皆表明,PMP、MDA、CHL、SOD酶活性和POD酶活性等是可靠的抗逆评价指标,而对于脯氨酸的可靠性则仍存在较大的争议[41-43],本研究灰色关联度分析结果同样显示脯氨酸含量与抗旱性之间关联系数最小,关联度最低,这与前人研究结果一致。
4 结论
水分胁迫下,36份5种气候生态型割手密F1和F2杂种材料中耐旱性较强的有云割F108-536、云割F211-179、云割F211-8、云割F108-474、云割F108-398、云割F108-254、云割F211-261、云割F211-151、云割F108-538、云割F108-391、云割F211-188、云割F211-37、云割F107-150、云割F211-258、云割F211-90,为强耐旱种质,可在甘蔗抗旱亲本培育及抗旱育种中加以重点利用;割手密耐旱特性在不同世代中的遗传表现不同,F1代耐旱性强于F2代;割手密后代耐旱性强弱与其父本割手密的气候生态类型、地理位置及海拔无明显相关性;7个生理生化指标中,PMP、MDA含量、CHL含量、SOD和POD酶活性与甘蔗耐旱性关联度较高,是耐旱性评价的优良指标。
[1] LIU J Y, BASNAYAKE J, JACKSON P A, CHEN X K, ZHAO J, ZHAO P F, YANG L H, BAI Y D, XIA H M, ZAN F G, QIN W, YANG K, YAO L, ZHAO L P, ZHU J R, LAKSHMANAN P, ZHAO X D, FAN Y H. Growth and yield of sugarcane genotypes are strongly correlated across irrigated and rainfed environments., 2016, 196: 418-425.
[2] 陈义强, 邓祖湖, 郭春芳, 陈如凯, 张木清. 甘蔗常用亲本及其衍生品种的抗旱性评价. 中国农业科学, 2007, 40(6): 1108-1117.
CHEN Y Q, DENG Z H, GUO C F, CHEN R K, ZHANG M Q. Drought resistant evaluations of commonly used parents and their derived varieties ., 2007, 40(6): 1108-1117. (in Chinese)
[3] MING R, MOORE P H, WU K K, D’Hont A, GLASZMANN J C, TEW T L, MIRKOV T E, SILVA J D, JIFON J, RAI M, SCHNELL R J, BRUMBLEY S M, LAKSHMANAN P, COMSTOCK J C, PATERSON A H. Sugarcane improvement through breeding and biotechnology.,2006, 27: 115-118.
[4] BEGCY K, MARIANO E D, GENTILE A, LEMBKE C G, ZINGARETTI S M, SOUZA G M, MENOSSI M. A novel stress-induced sugarcane gene confers tolerance to drought, salt and oxidative stress in transgenic tobacco plants., 2012, 7(9): 1-14.
[5] MIRSHAD P P, PUTHUR J T. Drought tolerance of bioenergy grassL. Enhanced by arbuscular mycorrhizae., 2017, 3: 1-8.
[6] GRAÇA J P D, RODRIGUES F A, FARIAS J R B, DEOLIVEIRA M C N, HOFFMANN-CAMPO C B, ZINGARETTI S M. Physiological parameters in sugarcane cultivars submitted to water deficit., 2010, 22(3): 187-197.
[7] 罗俊, 林彦铨, 张木清, 吕建林, 陈如凯, 洪伟雄. 利用甘蔗叶绿体荧光参数及活性氧代谢等指标评价甘蔗品种的耐旱性. 甘蔗, 1999, 6(3): 14-22.
LUO J, LIN Y Q, ZHANG M Q, LüJ L, CHEN R K, HONG W X. Assessment of drought resistance of sugarcane varieties through chlorophyll a fluorescence induction kinetics parameters of leaf chloroplast and active oxygen metabolism., 1999, 6(3): 14-22. (in chinese)
[8] 罗俊, 张木清, 林彦铨, 张华, 陈如凯. 甘蔗苗期叶绿素荧光参数与抗旱性关系研究. 中国农业科学, 2004, 37(11): 1718-1721.
LUO J, ZHANG M Q, LIN Y Q, ZHANG H, CHEN R K. Studies on the relationship of chlorophyll fluorescence characters and drought tolerance in seedling of sugarcane under water stress., 2004, 37(11): 1718-1721. (in chinese)
[9] ZHAO D L, GLAZ B, COMSTOCK J C. Sugarcane response to water-deficit stress during early growth on organic and sand soils., 2010, 5(3): 403-414.
[10] SILVA M D A, JIFON J L, SILVA J A G D, SANTOS C M D, SHARMA V. Relationships between physiological traits and productivity of sugarcane in response to water deficit., 2012, 152(1): 104-118.
[11] BASNAYAKE J, JACKSON P A, INMAN-BAMBER N G, LAKSHMANAN P. Sugarcane for water-limited environments. Genetic variation in cane yield and sugar content in response to water stress., 2012, 63(16): 6023-6033.
[12] 许文花, 杨清辉. 甘蔗割手密无性系抗旱性鉴定. 亚热带农业研究, 2005, 1(1): 22-26.
XU W H, YANG Q H. Drought resistance ofL. clones., 2005, 1(1): 22-26. (in Chinese)
[13] 罗明珠, 刘子凡, 梁计南, 魏延明. 甘蔗抗旱性与叶片某些生理生化性状的关系. 亚热带农业研究, 2005, 1(1): 14-16.
LUO M Z, LIU Z F, LIANG J N, WEI Y M. The relationship between drought resistance of sugarcane and some physiological biochemical properties of leaves., 2005, 1(1): 14-16. (in Chinese)
[15] EIBASHITI T, HAMAMC H, O¨KTEM H A, YUCEL M. Biochemical analysis of trehalose and its metabolizing enzymes in wheat under abiotic stress conditions., 2005, 169: 47-54.
[16] HURA T, GRZESIAK S, HURA K, THIEMT E, TOKARZ K, WEDZONY M. Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter Triticale: accumulation of ferulic acid correlates with drought tolerance.,2007, 100(4): 767-775.
[17] 桃联安, 杨李和, 经艳芬, 段慧芬, 董立华, 安汝东, 周清明, 朱建荣. 云南割手密血缘F2代抗旱性隶属函数法综合评价. 西南农业学报, 2011, 24(5): 1676-1680.
TAO L A, YANG L H, JING Y F, DUAN H F, DONG L H, AN R D, ZHOU Q M, ZHU J R .Comprehensive evaluation of drought resistance for descendant F2ofin Yunnan by subjection function.,2011, 24(5): 1676-1680. (in Chinese)
[18] 边芯, 董立华, 孙有芳, 桃联安, 朱建荣, 周清明, 杨李和, 安汝东, 郎荣斌, 俞华先, 冯蔚, 经艳芬. 云南割手密及其血缘F1代材料抗旱相关性状的主成分分析. 干旱地区农业研究, 2014, 32(3): 56-61.
BING X, DONG L H, SUN Y F, TAO L A, ZHU J R, ZHOU Q M, YANG L H, AN R D, LANG R B, YU H X, FENg W, JING Y F. Principal component analysis of drought resistance related traits ofL. and its F1hybrids., 2014, 32(3): 56-61. (in Chinese)
[19] 高三基, 罗俊, 陈如凯, 张木清, 潘大仁. 甘蔗品种抗旱性光合生理指标及其综合评价. 作物学报, 2002, 28(1): 94-98.
GAO S J, LUO J, CHEN R K, ZHANG M Q, PAN D R. Photosynthetic physiology indexes of the drought resistance of sugarcane and its comprehensive evaluation.,2002, 28(1): 94-98. (in Chinese)
[20] JACKSON P A, MCRAE T A. Gains from selection of broadly adapted and specifically adapted sugarcane families., 1998, 59(3): 151-162.
根据本项目的经验,随着大体积混凝土施工应用越来越普遍,大体积混凝土的施工质量不仅仅会关联到国家经济的发展,还关系到人民群众的生命财产安全,所以大体积混凝土施工需要严格按照规矩来施工,改善施工的工艺,尽最大的力量来减少大体积混凝土施工裂纹/裂缝,以此提高大体积混凝土的施工质量。
[21] STRINGER J K, CULLIS B R, THOMPSON R. Joint modeling of spatial variability and within-row inter plot competition to increase the efficiency of plant improvement., 2011, 16(2): 269-281.
[22] 赵培方, 赵俊, 刘家勇, 昝逢刚, 夏红明, JACKSON P A, Basnayake J, Inman-Bamber N G, 杨昆, 赵丽萍, 覃伟, 陈学宽, 赵兴东, 范源洪. 干旱胁迫对甘蔗4个生理指标遗传变异的影响. 中国农业科学, 2017, 50(1): 28-37.
ZHAO P F, ZHAO J, LIU J Y, ZAN F G, XIA H M, JACKSON P A, BASNAYAKE J, INMAN-BAMBER N G, YANG K, ZHAO L P, QIN W, CHEN X K, ZHAO X D, FAN Y H. Genetic variation of four physiological indexes as impacted by water stress in sugarcane.,2017, 50(1): 28-37. (in Chinese)
[23] 经艳芬, 边芯, 桃联安, 董立华, 周清明, 朱建荣, 安汝东, 杨李和, 郎荣斌, 俞华先, 冯蔚. 云南割手密血缘F1创新种质的因子和聚类分析. 植物遗传资源学报, 2014, 15(1): 177-181.
JING Y F, BIAN X, TAO L A, DONG L H, ZHOU Q M, ZHU J R, AN R D, YANG L H, LANG R B, YU H X, FENG W. Factor and cluster analysis of Yunnan innovated germplasm materials F1of., 2014, 15(1): 177-181. (in Chinese)
[24] 桃联安, 安汝东, 杨李和, 经艳芬, 董立华, 周清明, 朱建荣, 边芯. 甘蔗属割手密种(Saccharum spontaneum)‘云南82-114’的F2(BC1)代同异法综合评价及同一度分类. 热带亚热带植物学报, 2014, 22(6): 590-600.
TAO L A, AN R D, YANG L H, JING Y F, DONG L H, ZHOU Q M, ZHU J R, BIAN X. Integrated evaluating and classifying of the similarity-difference analysis for the generation F2(BC1) from‘Yunnan 82-114’., 2014, 22(6): 590-600. (in Chinese)
[25] 张琼, 齐永文, 张垂明, 陈勇生, 邓海华. 我国大陆甘蔗骨干亲本亲缘关系分析. 广东农业科学, 2009(10): 44-48.
ZHANG Q, QI Y W, ZHANG C M, CHEN Y S, DENG H H. Pedigree analysis of genetic relationship among core parents of sugarcane in Mainland China., 2009(10):44-48. (in Chinese)
[26] 谭常. 植物生理学实验手册. 上海: 上海科技文化出版社, 1985.
TAN C.. Shanghai: Shanghai Scientific Technology and Culture Press, 1985. (in Chinese)
[27] 李忠光, 龚明. 植物生理学综合性和设计性实验教程. 武汉: 华中科技大学出版社, 2014: 52-144.
LI Z G, GONG M.. Wuhan: Huazhong university of science technology press, 2014: 52-144. (in Chinese)
[28] 张保青, 杨丽涛, 李杨瑞. 自然条件下甘蔗品种抗寒生理生化特性的比较. 作物学报, 2011, 37(3): 496-505.
ZHANG B Q, YANG L T, LI Y R. Comparison of physiological and biochemical characteristics related to cold resistance in Sugarcane under field conditions., 2011, 37(3): 496-505. (in Chinese)
[29] 高三基, 罗俊, 张华, 陈如凯, 林彦铨. 甘蔗抗旱性生理生化鉴定指标. 应用生态学报, 2006, 17(6): 1051-1054.
GAO S J, LUO J, ZHANG H, CHEN R K, LIN Y Q. Physiological and biochemical indexes of drought resistance of sugarcane (spp.)., 2006, 17(6): 1051-1054. (in Chinese)
[30] 白向历, 齐华, 刘明, 张振平. 玉米抗旱性与生理生化指标关系的研究. 玉米科学, 2007, 15(5): 79-83.
BAI X L, QI H, LIU M, ZHANG Z P. Study on the relationship between drought resistance and physiological index of., 2007, 15(5): 79-83. (in Chinese)
[31] 汪德水. 旱地农田水肥协同效应与耦合模式. 北京: 气象出版社, 1999. WANG D S.. Beijing: Meteorology Press, 1999. (in Chinese)
[32] 朱建荣, 桃联安, 董立华, 周清明, 安汝东, 杨李和, 段惠芬, 刘洪博, 经艳芬. 云南不同生态型甘蔗细茎野生种育种潜力分析. 南方农业学报, 2011, 42(9): 1035-1040.
ZHU J R, TAO L A, DONG L H, ZHOU Q M, AN R D, YANG L H, DUAN H F, LIU H B, JING Y F. Breeding potential ofL. collected from several ecological areas of Yunnan., 2011, 42(9): 1035-1040. (in Chinese)
[33] 庄勇, 陈龙正, 杨寅桂, 娄群峰, 陈劲枫. 植物异源多倍体进化中基因表达的变化. 植物学通报, 2006, 23(2): 207-214.
ZHUANG Y, CHEN L Z, YANG Y G, LOU Q F, CHEN J F. Changes in gene expression in evolution of plant allopolyploids., 2006, 23(2): 207-214. (in Chinese)
[34] ADAMS K L, WENDEL J F. Exploring the genomic mysteries of polyploidy in cotton.,2004, 82(4): 573-581.
[35] HE P, FRIEBE B R, GILL B S, ZHOU J M. Allopolyploidy alters gene expression in the highly stable hexaploid wheat., 2003, 52(2): 401-414.
[36] KASHKUSH K, FELDMAN M, LEVY A A. Gene loss, silencing and activation in newly synthesized wheat allopolyploid., 2002, 160: 1651-1659.
[37] 程建峰, 潘晓云, 刘宜柏, 戴廷波, 曹卫星. 水稻抗旱性鉴定的形态指标. 生态学报, 2005, 25(11): 3117-3125.
CHENG J F, PAN X Y, LIU Y B, DAI T B, CAO W X. Morphological indexes of drought resistance identification in rice., 2005, 25(11): 3117-3125. (in Chinese)
[38] 钮福祥, 华希新, 郭小丁, 邹景禹, 李洪民, 丁成伟. 甘薯品种抗旱性生理指标及其综合评价初探. 作物学报, 1996, 22(4): 392-398.
NIU F X, HUA X X, GUO X D, ZOU J Y, Li H M, DING C W. Studies on several physiological indexes of the drought resistance of sweet potato and comprehensive evaluation., 1996, 22(4): 392-398. (in Chinese)
[39] 白亚东, 赵俊, 杨桂林, 赵兴东, 卢顺才, 李秀梅, 杨丽华, 张燕艳. 10个云蔗系列甘蔗品种(系)灰色关联度多维综合评估. 中国糖料, 2016, 38(6): 27-30.
BAI Y D, ZHAO J, YANG G L, ZHAO X D, LU S C, LI X M, YANG L H, ZHANG Y Y. Integrated evaluation of 10 CYZ sugarcane varieties by gray correlation degree., 2016, 38(6): 27-30. (in Chinese)
[40] 赵俊, 白亚东, 赵兴东, 杨丽华, 卢顺才, 李秀梅, 马云飞, 李梅会, 王少河, 范源洪. 灌溉与干旱条件下甘蔗工农艺性状的相关性分析与品种抗旱性评价. 湖南农业大学学报(自然科学版), 2016, 42(6): 579-586.
ZHAO J, BAI Y D, ZHAO X D, YANG L H, LU S C, LI X M, MA Y F, LI M H, WANG S H, FAN Y H. Drought resistance evaluation and correlation analysis of agronomic and quality traits of sugarcane under irrigated and rain-fed conditions., 2016, 42(6): 579-586. (in Chinese)
[41] 夏红明, 赵培方, 吴才文, 刘家勇. 甘蔗品种抗旱性综合评价. 亚热带农业研究. 2014, 10(1):7-11.
XIA H M, ZHAO P F, WU C W, LIU J Y. Comprehensive evaluation on the drought resistance of sugarcane., 2014, 10(1): 7-11. (in Chinese)
[42] 卢会文, 杨清辉, 肖关丽, 赵正红, 黄淋华. 云南省主栽甘蔗品种抗旱性鉴定. 亚热带农业研究, 2012, 8(1): 8-12.
LU H W, YANG Q H, XIAO G L, ZHAO Z H, HUANG L H. Identification of drought resistance in dominant sugarcane varieties in Yunnan., 2012, 8(1): 8-12. (in Chinese)
[43] 兰靖, 陈永, 陆玉朵, 韩世健, 裴铁雄, 陆国盈, 李贤宇, 周洁琼. 6个甘蔗品种(系)的抗旱性差异评价. 西南农业学报, 2014, 274(4): 1374-1381.
LAN J, CHEN Y, LU Y D, HAN S J, PEI T X, LU G Y, LI X Y, ZHOU J Q. Comprehensive evaluation on drought resistance difference of six sugarcane varieties.,2014, 27(4): 1374-1381. (in Chinese)
(责任编辑 李莉)
Drought Resistance Evolution of F1andF2Hybrids from Five Climatic EcotypesL.
TIAN ChunYan, TAO LianAn, YU HuaXian, DONG LiHua, JING YanFen, BIAN Xin, LANG RongBin, ZHOU QingMing, AN RuDong, SUN YouFang, YANG LiHe
(Ruili Breeding Station, Sugarcane Research Institute, Yunnan Academy of Agricultural Sciences, Ruili 678600, Yunnan)
Drought is an important abiotic factor which has a severe impact on yield and qualities of sugarcane production.L., a wild resource of sugarcane, possesses a series of characters such as strong stress resistance, excellent perennial root and wide adaptability. Therefore, study on the drought resistance of its progenies, screening of drought resistant germplasms could provide a basis for drought resistance parent selection and realize breakthrough in breeding new sugarcane cultivars.In this study, thirty-six F1andF2individuals from five ecotypeswithYunrui innovation parents, introduced species and domestic species were bucket-planted in the greenhouse used as materials. The materials were treated by artificial water stress including normal water supply (as control), mild water stress and severe water stress at the early elongation stage of sugarcane. Subsequently, seven physiological and biochemical indexes including plasma membrane permeability (PMP), contents of malondialdehyde (MDA), chlorophyll (CHL), soluble protein (Pr) and proline (Pro) content and activities of superoxide dismutase (SOD) and peroxidase (POD) in sugarcane leaves were measured. Eventually, the drought resistance of F1andF2generations was evaluated comprehensively using fuzzy membership function method, cluster and grey relational analysis.Under water stress, the PMP, contents of MDA and proline, and the activities of SOD and POD raised, while the contents of CHL and PRO declined with different extents depending on sugarcane genotypes and stress degree. The results of fuzzy membership function analysis indicated that the drought resistance of.progeny varied was different under different water stress degrees, and the drought resistance of F1generation was stronger than that of F2. The comprehensive drought resistance of individuals exceeded the parental up to 82% in F1,which was much higher than F2. The tested materials were classified into four groups under mild water stress by cluster analysis, and the drought resistance of groups Ⅰ and Ⅱ was the best, followed by group Ⅳ, and the worst was group Ⅲ. Moreover, the individuals of group Ⅲ were all from F2, and all the F1individuals distributed in groupsⅠ, Ⅱ and Ⅳ, respectively, which had strong drought resistance. Simultaneously, they could be divided into three groups under severe stress, and the F1individualsexcepted YGF108-473 were all distributed in groups Ⅰ and Ⅲ, which showed stronger drought resistance than group Ⅱ. The grey relation analysis revealed that the correlation between physiological indexes and membership value was different because of water stress degree. Under mild stress, the correlation degree was in the order of POD>SOD>PMP>MDA>Pr>CHL>Pro. But, the correlation degree under severe stress was in the order of SOD>Pr>CHL>POD>PMP>MDA>Pro.In all thirty-six tested materials, YGF108-536, YGF211-261, YG F108-391, YGF108-254, YGF211-90, YGF211-8, YGF211-258, YGF108-398, YGF108-543, YGF211-151 and YGF211-53 were exhibited strong resistance under water stress. These materials could be utilized emphatically in parent selection and drought resistance breeding. Furthermore, genetic performance of drought resistance had significant difference between F1and F2hybrids of, and the drought resistance of F1wasstronger than that of F2.In addition, the drought resistance ofhybrids was no obvious correlation with paternal climate-type, geographic position and altitude. Based on the results of this study, PMP, contents of MDA and CHL, and activities of SOD and POD were good physiological and biochemical indicators with close correlation to drought resistance of sugarcane.
L.; water stress; physiological and biochemical indexes; drought resistance; sugarcane
2017-04-24;
国家自然科学基金(31360358)、云南省重点基金(2015FA034)、云南省科技惠民计划(2014RA059)、云南省科技创新人才计划(2014HCO15)、云南省甘蔗遗传改良重点实验室开放基金(2015DG015-05、2015DG015-09)
接受日期:2017-06-02
联系方式:田春艳,Tel:0692-4144319;E-mail:tianchy89@126.com。通信作者经艳芬,E-mail:rljyf@126.com
