节水抗旱稻旱优113号的根系生长对土壤水分亏缺的响应
2017-11-28补红英宋维周曹凑贵李萍
补红英,宋维周,曹凑贵,2,李萍,2
节水抗旱稻旱优113号的根系生长对土壤水分亏缺的响应
补红英1,宋维周1,曹凑贵1,2,李萍1,2
(1华中农业大学植物科学技术学院,武汉 430070;2主要粮食作物产业化湖北省协同创新中心(长江大学),湖北荆州 434025)
揭示土壤水分亏缺条件下节水抗旱稻根系的形态和生理指标的变化规律,阐明其节水抗旱的特异性生理基础。2015和2016年利用盆栽试验,以节水抗旱稻旱优113号(HY113)和高产水稻扬两优6号(YLY6)为试验材料,通过设置淹灌(对照)和干旱(土壤水势-38 kPa左右)处理,研究干旱对节水抗旱稻与高产水稻根系形态结构和生理指标的影响及其与地上部生物量积累的关系。与淹灌相比,干旱处理显著降低了两个水稻品种的地上、地下干物质积累量,同时显著降低了节水抗旱稻HY113的根冠比(由0.18降为0.12),高产水稻YLY6的根冠比无显著变化。干旱处理下,两个品种的水稻根系活力均显著增加,但HY113的增加幅度显著高于YLY6,乳熟期HY113在2015和2016两年的平均增幅为38.7%,而YLY6为22.8%,其中2015年乳熟期HY113的根系活力从86 µg·g-1·h-1增加至174 µg·g-1·h-1。干旱处理下,HY113和YLY6的根系吸收面积均显著减小,但HY113的减小幅度显著低于YLY6。与高产水稻YLY6相比,节水抗旱稻HY113的根数、根体积和根干重均较少,总干物质积累量较少,但其根系活力和根系有效吸收面积较大。节水抗旱稻HY113具有根量少,但根系吸收效率高的特点,其在缺水条件下能维持较高的根系活力和根系吸收面积;在遭遇水分亏缺时HY113可通过减小根冠比,使得更多的干物质留在地上部分以保证籽粒产量。
节水抗旱稻;水分亏缺;根冠比;根系特性
0 引言
【研究意义】中国是水资源极为短缺的国家,水资源危机已经成为制约经济社会可持续发展的瓶颈因素[1]。农业用水效率低下,其中水稻生产中的用水浪费严重是主要问题之一[2-3],如何提高其水分利用效率已经成为研究热点。目前,关于节水灌溉技术研究已获得较好的成果,人们进而逐渐开始挖掘植物自身的抗旱节水潜力[4]。近些年,节水抗旱稻品种已培育成功,节水抗旱稻是一种新型水稻品种,与当前水稻品种相比,不仅有高产潜力,也具有抗旱节水能力[5]。水稻根系是吸收水分的重要器官,其生长情况与活力会直接影响整个水稻的生长发育、营养状况和产量水平[6]。迄今为止对水稻根系的研究较多,但专门针对节水抗旱稻的根系形态、生理及其对干旱的特异性应对策略等方面的研究仍比较有限[7],节水抗旱稻的根系具有哪些特点以达到节水抗旱的效果尚不清楚,进一步研究水稻节水抗旱机理有重要意义。【前人研究进展】已有研究表明,节水抗旱稻可以减少约50%的水分消耗,同时与普通水稻相比,其产量不会显著减少[8],但节水抗旱稻的产量水平因不同品种而异。现有一些研究发现在节水灌溉条件下,高产水稻扬两优6号的产量比节水抗旱稻旱优113号高21.5%,而有人却发现在相同水分灌溉模式下,节水抗旱稻旱优8号的产量比高产水稻两优培九高17%,比高产水稻陵香优18高11.3%[9-10]。前人研究表明良好的根系形态可提高水稻根系代谢能力和籽粒库的生理活性,能促进水稻高产[11-13]。干旱逆境下植株强大的根系有利于植株维持较好的水分状况和生理功能。多数研究发现在干旱胁迫下,或控制灌溉时,水稻根系生物量相对增加,总根数、根长、根体积及干物质积累均增加,同化物在地上部分的分配比例会有所降低[14-17]。然而,也有人提出了“根系冗余生长”的观点[18-20],即植株的根量过大会消耗大量的同化物而对产量产生不利的影响,因为根系既是水分和养分的吸收器官,但同时又需要消耗地上部提供的光合产物用以形态建成和维持生长。并且与地上部相比,用于生产单位根干物质量所消耗的能量是地上部的2倍,干旱逆境下如果根系过大并不利于维持较高的籽粒产量。【本研究切入点】在干旱缺水时,节水抗旱稻的根系形态和生理特性如何变化,根系和地上部生物量积累如何协调以适应水分亏缺尚缺乏专门报道。本文以节水抗旱稻(HY113)和高产水稻(YLY6)为材料,通过设置干旱处理,研究干旱逆境下HY113的根系形态和生理指标的变化规律。【拟解决的关键问题】本研究旨在揭示节水抗旱稻HY113的根系生长对土壤水分亏缺的特异性响应机制,为实现水稻节水栽培提供理论依据。
1 材料与方法
1.1 试验地点、时间与材料
于2015年和2016年在湖北省武汉市华中农业大学(114°29' E,30°28' N)校内网室进行盆栽试验。试验品种为旱优113号(HY113)和扬两优6号(YLY6),其中HY113是籼型三系杂交旱稻,属于节水抗旱稻品种,由上海农业生物基因中心提供;YLY6为籼型两系杂交水稻,是湖北省大面积推广的高产水稻品种。
1.2 试验设计
使用高30 cm、直径30 cm底部密封的塑料桶,将与塑料桶相似大小的根袋套于桶内。试验土壤为黏壤土,土:沙比为3﹕1,每桶装土沙混合物12.5 kg。采用秧田水育秧,2015年5月17日播种,5月31日带土移栽,10月中旬收获;2016年5月19日播种,6月5日带土移栽,10月中旬收获。每桶定植生长大小相近的3株秧苗。土壤中全氮含量为1.23 g·kg-1,全磷为122.7 mg·kg-1,速效磷为33.82 mg·kg-1,每盆施用20 g复合肥(N﹕P2O5﹕K2O=15﹕15﹕15)作底肥,抽穗开花期每盆再追施1 g尿素。
试验设置淹灌(全生育期保持3—5 cm水层)和干旱(土壤水势-38±4 kPa)两个水分处理,每个处理12次重复。于水稻移栽返青后开始控制水分,返青期间每盆均留3—5 cm水层。水分处理开始后根据土壤水势变化进行定量补水,并记录累计灌水量(图1)。利用土壤张力计(LBT-TEN50)对土壤水分状况进行监测并记录(图2)。
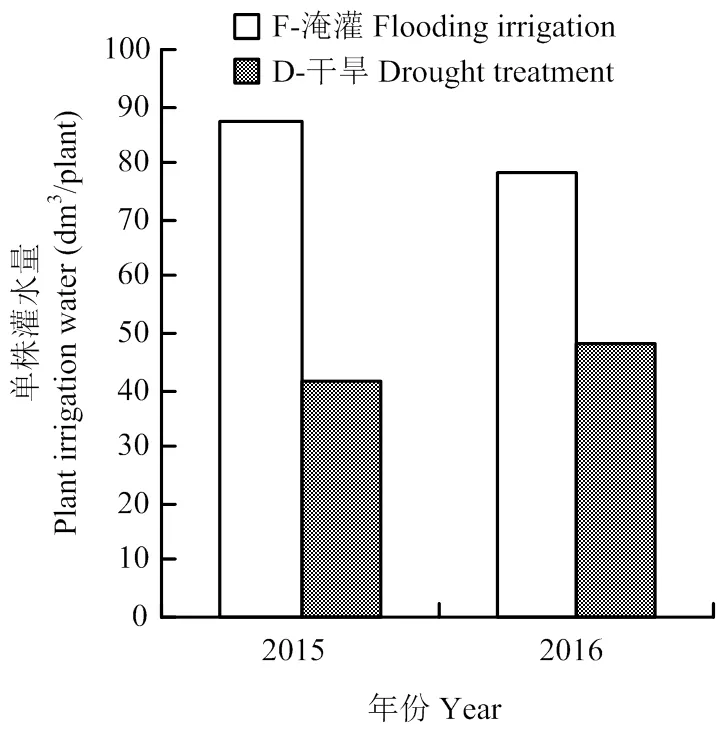
图1 2015年和2016年水稻生育期内淹灌(F)和干旱(D)的单株总灌水量
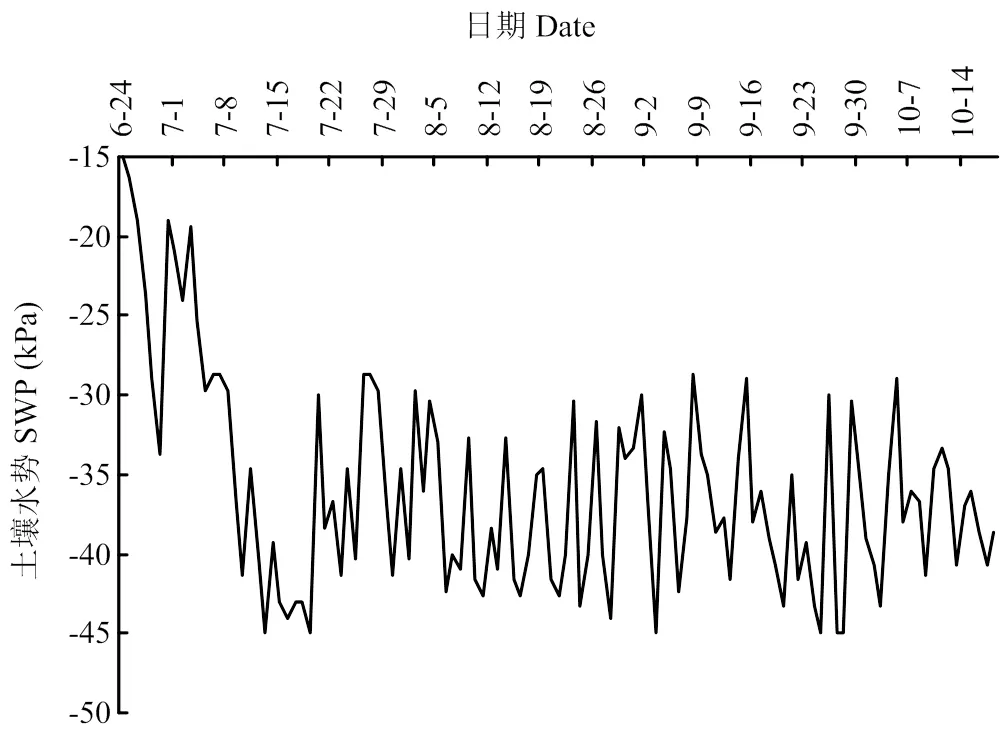
图2 2016年干旱处理的土壤水势动态变化
1.3 测定项目与方法
1.3.1 环境指标监测 分别使用TPJ-20 温湿度记录仪(浙江托普仪器有限公司)和ZC-FS太阳总辐射测试仪(北京哲成科技有限公司)自动对网室内环境温度和太阳总辐射进行全天(设定每隔30min 记录一次)记录监测。
太阳总辐射量:由北京哲成科技有限公司生产的ZC-FS太阳总辐射传感器对网室内进行全天(设定每隔30min 记录一次)自动记录监测。
1.3.2 叶片水势与气叶温差测定 叶片水势测定:晴天中午 11:00—13:00取冠层完全展开叶或剑叶3 片,放入封口塑料袋置于冰盒带回室内,采用露点水势仪(WP4C,DECAGON Inc.,USA)测定叶片水势。
气叶温差测定:采用UT300A红外测温仪,分别于分蘖盛期、孕穗期、灌浆乳熟期,在无风晴朗天气的下午13:00—15:00,在距离叶片1 m处测定植株冠层完全展开叶或剑叶叶片中部向光部分的叶温,同时记录此时网室中大气温度,测得叶温与网室中温度的差值即为气叶温差。
1.3.3 根系特性测定 根系形态指标(根直径、体积、数目等)测定:在孕穗期、乳熟期、成熟期3个时期取鲜样,根直径采用游标卡尺测定;根体积采用排水法测定;根数目人工统计所得(本研究根数目为水稻的不定根数目)。
根系活力的测定:在孕穗期、乳熟期2个时期,剪取3—5 cm长的根尖0.2—0.3 g,采用氯化三苯基四氮唑(TTC)法测定(本研究将根系还原力作为根系活力)。
根系吸收面积与活跃吸收面积:在孕穗期、乳熟期和成熟期3个时期,取完整根称重后,将其放入甲烯蓝溶液,然后用分光光度计测定浸染后的甲烯蓝溶液浓度,用来计算吸收面积。
1.3.4 干物质量测定 分别在孕穗期、乳熟期、成熟期3个时期,取测定根形态的水稻植株,分根、茎、叶(穗),在105℃烘箱杀青30 min后,于80℃恒温下烘干至恒重,用1/100电子天平称重。
1.4 统计分析
试验数据采用Excel整理,试验数据的方差分析和相关性分析采用 SPSS系统处理,主成分分析采用CANOCO(CANonical Community Ordination)系统处理。
2 结果
2.1 环境指标与方差分析
2.1.1 环境温度 由图3可知,2015年气温在7月中旬到9月中旬的日最高温度、日平均温度、日最低温度均相对平稳,日平均温度基本保持在28—32℃,气温从9月下旬开始逐渐下降。2016年水稻生育期气温波动较大,6月下旬到9月下旬日平均温度最高达41.8℃,最低仅为23.7℃。其中,在水稻孕穗期及乳熟期遭遇了接近1周的低温,对水稻的生长和产量造成了一定的影响。
2.1.2 太阳总辐射量 由图4可知,2015年日太阳总辐射量普遍高于2016年,除6月26日至7月8日和8月4日至8月11日外,随着水稻生育期的推进,两年的太阳总辐射变化趋势基本一致。2015年6月2日至10月15日期间太阳总辐射量为2055.9 MJ·m-2,高于2016年的1974.6 MJ·m-2。
2.1.3 方差分析结果 由方差分析结果可知,根直径、孕穗期叶水势、生育后期的根系吸收面积在两个品种间差异不显著,其他指标在品种间均存在显著差异。除根冠比外,所有的指标受干旱处理的影响极显著。生育后期两水稻品种的生物量(地上干重和根系干重)在2015年与2016年差异显著,这与两年的气候差异有关,地上部干重、根冠比、根系活力在几个生育期均无品种×处理(G×T)的交互作用,而根直径存在显著的G×T交互作用(表1)。
2.2 叶片水势与气叶温差对干旱胁迫的响应
由图5可知,干旱胁迫显著降低了叶水势,而且两个品种的乳熟期叶水势均显著高于孕穗期;HY113的叶水势在孕穗期和乳熟期两年平均降低54.5%和118.4%;而YLY6的叶水势在孕穗期和乳熟期分别降低19.3%和14.4%,虽然HY113在干旱处理下叶水势降低幅度大于YLY6,但除2015年的干旱处理(D-HY113<D-YLY6)外,HY113的叶水势一直显著高于YLY6,表明植株具有较好的水分状况。
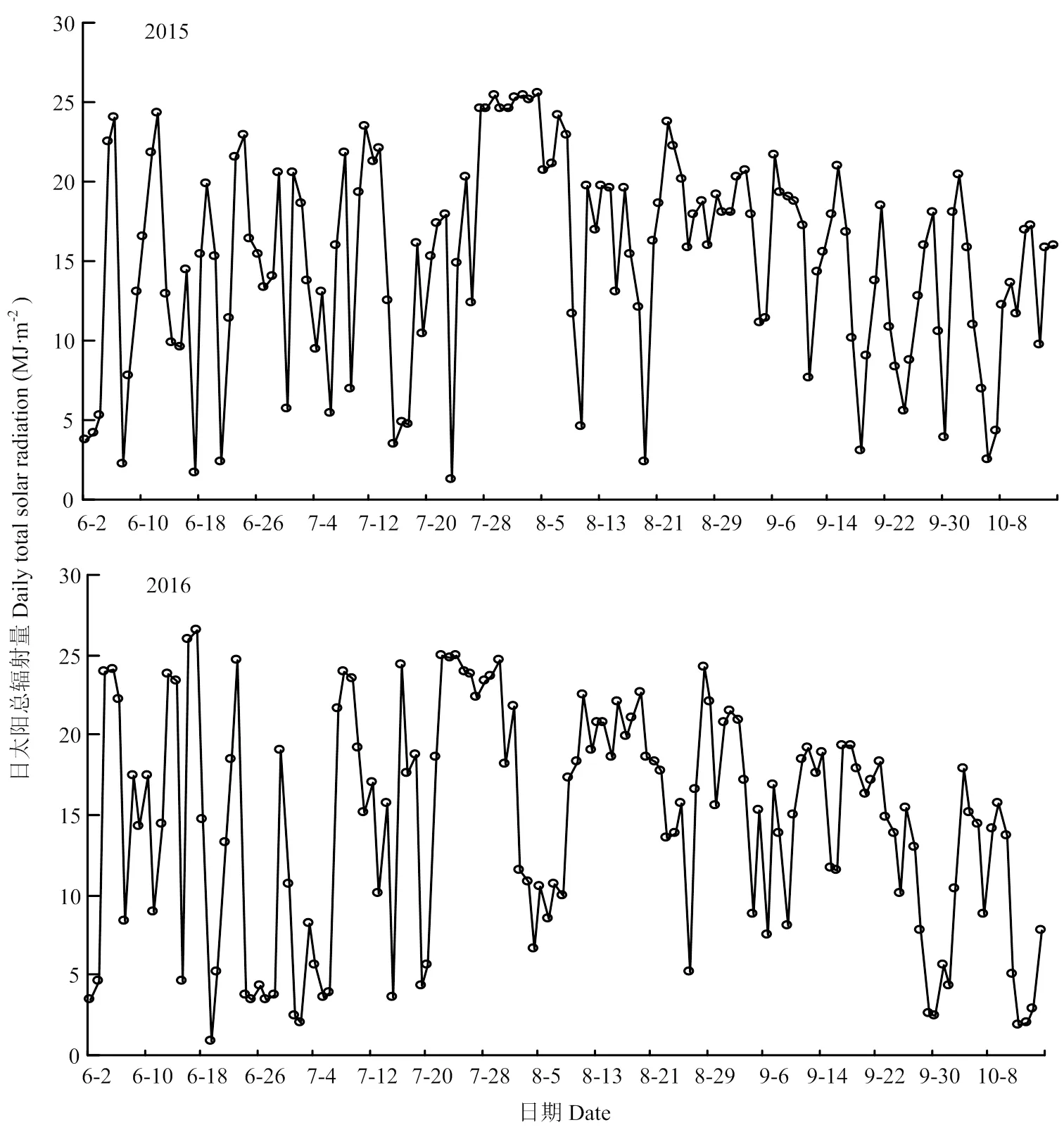
图4 2015和2016年水稻生育期(6月2日至10月15日)内每天00:00-23:59的太阳总辐射量的变化
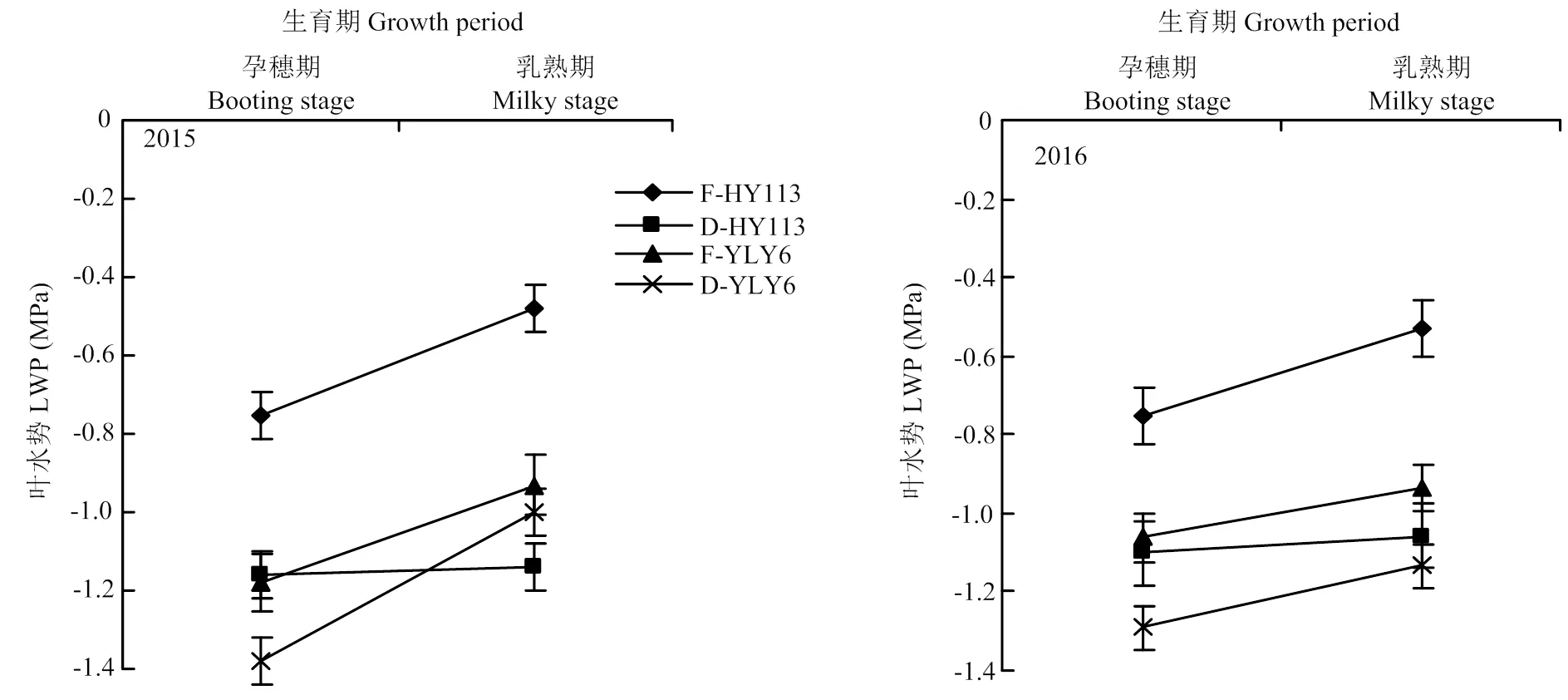
图5 淹灌(F)和干旱处理(D)下旱优113号(HY113)和扬两优6号(YLY6)在不同生育期的叶片水势
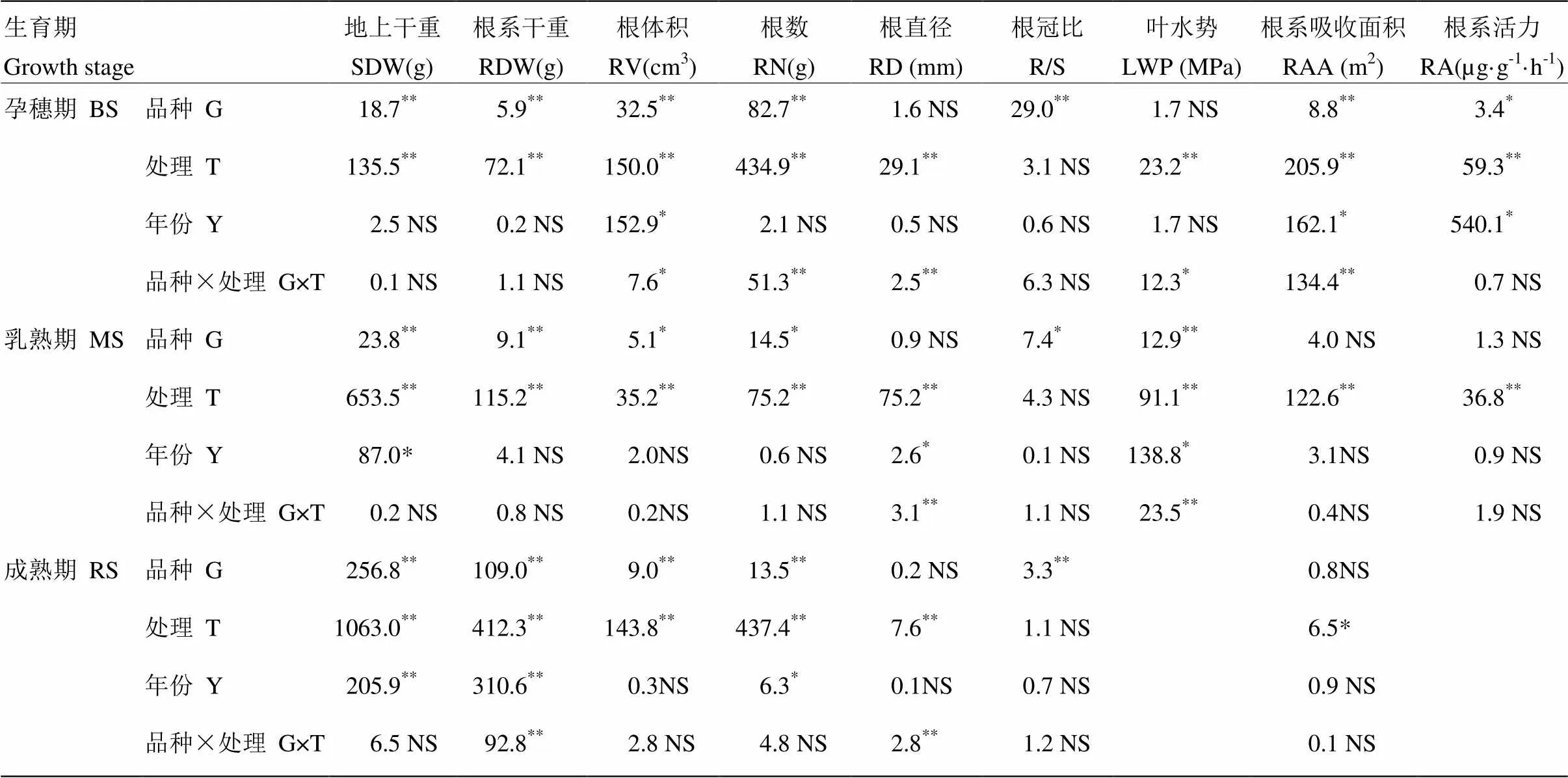
表1 不同水分处理对节水抗旱稻与高产水稻各指标方差分析结果
SDW:地上干重;RDW: 根系干重;RV:根体积;RN:根数;RD:根直径;R/S:根冠比;LWP:叶水势;RAA:根系吸收面积;RA:根系活力。BS:孕穗期;MS:乳熟期;RS:成熟期。数字为值,NS表示不显著,*和**分别代表显著性水平为0.05和0.01,*,<0.05和 **,<0.01。下同
SDW: Shoot dry weight; RDW: Root dry weight; RV: Root volume; RN: Root number; RD: Root diameter; R/S: Root-shoot ratio; LWP: Leaf water potential; RAA: Root absorption area; RA: Root activity. BS: Booting stage; MS: Milky stage; RS: Ripening stage.The number meansvalue, NS means no significant difference,* and ** are significant at 5% and 1% levels of probability, respectively. *<0.05,**,<0.01. The same as below
与淹灌相比,干旱处理导致气叶温差显著降低,干旱胁迫下HY113的气叶温差在孕穗期和乳熟期两年平均降低10.5%和20.6%;YLY6在孕穗期和乳熟期分别降低40.1%和36.0%。干旱处理下,YLY6的气叶温差平均降幅较大,表明其叶片蒸腾失水量的减少幅度更大些(图6)。
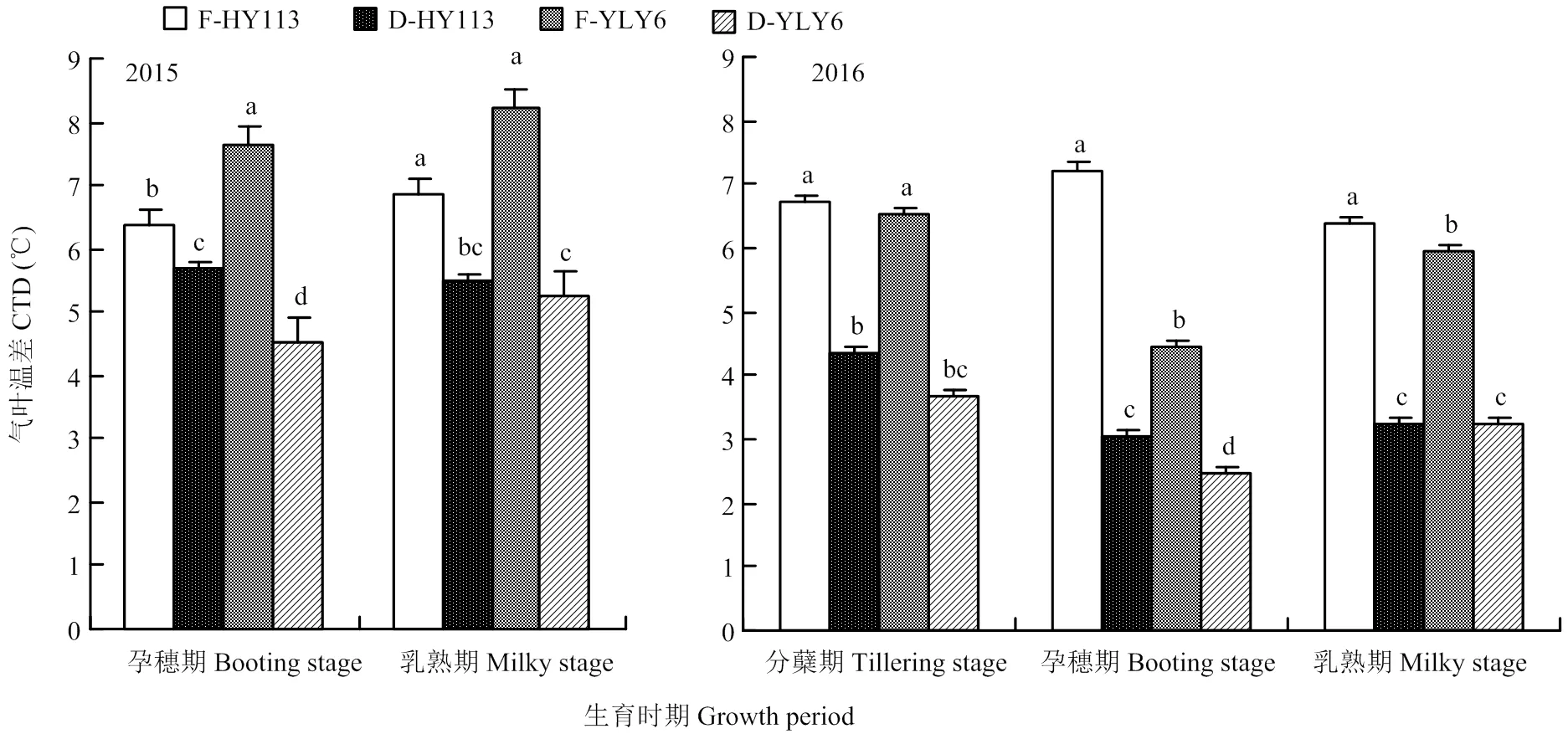
图6 淹灌(F)和干旱处理(D)下旱优113号(HY113)和扬两优6号(YLY6)在不同生育期的气叶温差
2.3 根系吸收面积与根系活力对干旱胁迫的响应
由表2可知,与淹灌相比,干旱处理导致节水抗旱稻和高产水稻的根系总吸收面积和活跃吸收面积均显著减小。淹灌条件下高产水稻YLY6的根系总吸收面积和活跃吸收面积显著大于或接近节水抗旱稻HY113;而在干旱处理下,HY113的根系吸收面积显著大于或接近YLY6。在干旱胁迫下,节水抗旱稻HY113的根系总吸收面积在孕穗期和乳熟期的两年,平均降幅分别为66.3%和51.1%,高产水稻YLY6在孕穗期和乳熟期分别降低73.7%和57.7%,根系活跃吸收面积在处理和品种间的变化基本与总吸收面积一致。表明干旱胁迫下HY113的根系吸收面积的减幅较小。
与传统淹灌相比,除2015年孕穗期的HY113外,干旱处理下水稻的根系活力均显著增强。干旱条件下HY113的根系活力在孕穗期和乳熟期两年分别增加了49.5%和66.8%,其中2015年乳熟期干旱处理下HY113根系活力的增幅最大,从86 µg·g-1·h-1增加至174 µg·g-1·h-1;而同期YLY6的根系活力分别增加了33.2%和33.7%。除2015年孕穗期外,节水抗旱稻HY113在干旱胁迫下根系活力显著高于高产水稻YLY6,表明其在干旱逆境下的根系吸水能力较强,且HY113根系具有抗旱潜力(图7)。
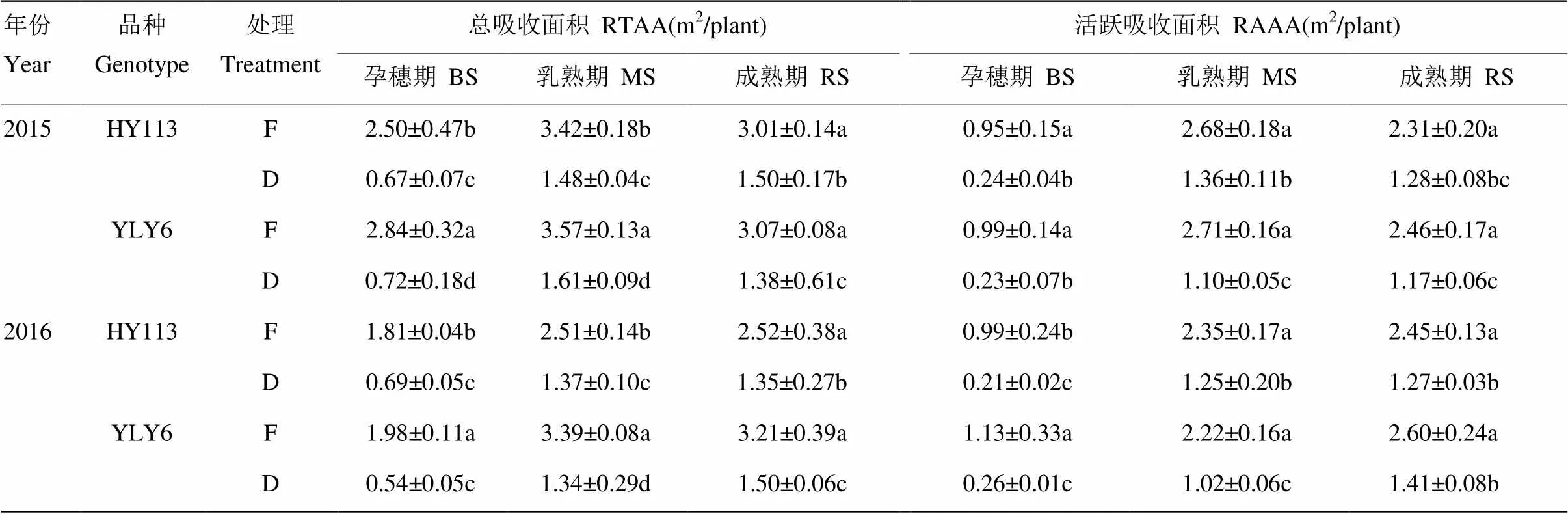
表2 淹灌(F)和干旱(D)处理下节水抗旱稻旱优113号(HY113)与高产水稻扬两优6号(YLY6)的根系吸收面积
RTAA:总吸收面积;RAAA:活跃吸收面积。表中多重比较仅限于同一取样时间,不同字母表示差异显著(<0.05)。下同
RTAA: Root total absorption area; RAAA: Root active absorption area. Multiple comparisons in the table is limited to the same sampling time, different letters mean significant difference (< 0.05).The same as below
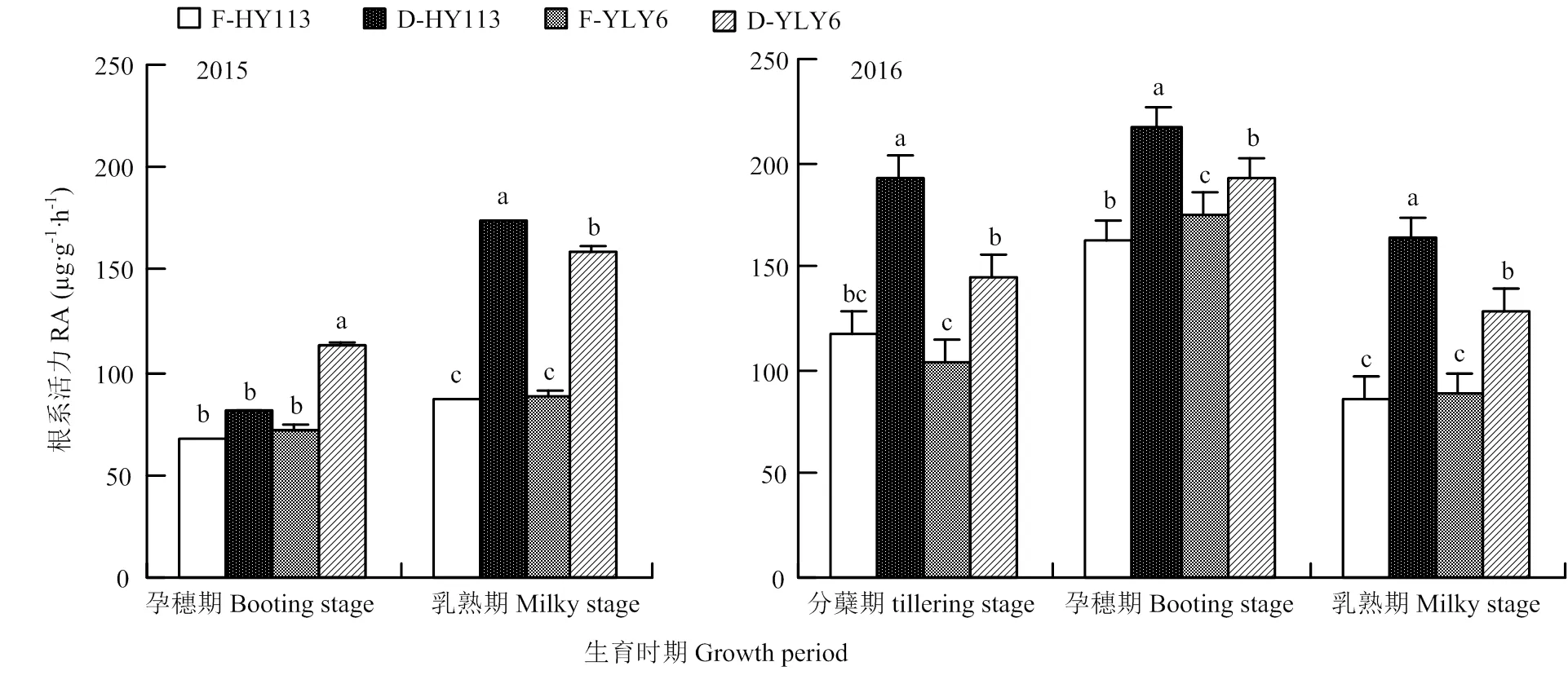
图7 淹灌(F)和干旱处理(D)下旱优113号(HY113)和扬两优6号(YLY6)在不同生育期的根系活力
2.4 根系形态对干旱胁迫的响应
如表3所示,与传统淹灌相比,干旱处理下在孕穗期两水稻品种的根直径显著减小,而除2016年乳熟期的YLY6外,干旱胁迫对水稻生育后期(乳熟期和成熟期)的根直径无显著影响;两个水稻品种的根直径无显著差异。与淹灌对照相比,干旱处理下水稻的单株根体积和单株根数均显著降低。在正常供水条件下,除2016年成熟期的单株根数外,节水抗旱稻HY113的单株根体积和单株根数均显著低于高产水稻YLY6。干旱胁迫下,除2015年乳熟期和2016年孕穗期外,HY113的单株根体积和单株根数均与YLY6无显著差异。表明节水抗旱稻HY113在水分充足时其根系较小,但在遭遇干旱胁迫时根量的减幅较小。
2.5 植株地上部、地下部的干物质积累与分配对干旱胁迫的响应
与淹灌对照相比,干旱处理下HY113与YLY6的植株地上部干重都显著降低(图8)。在孕穗期、乳熟期和成熟期,HY113的地上部干重两年平均降低了23.5%、44.7%和49.5%;YLY6同期分别降低了41.5%、44.4%和53.4%。两水稻品种间相比,淹灌条件下节水抗旱稻HY113的地上部干重显著小于或接近高产水稻YLY6;而干旱胁迫下,除2015孕穗期外,两个水稻品种间无显著差异。表明HY113遭遇干旱胁迫后地上部干物质积累的减幅较小。
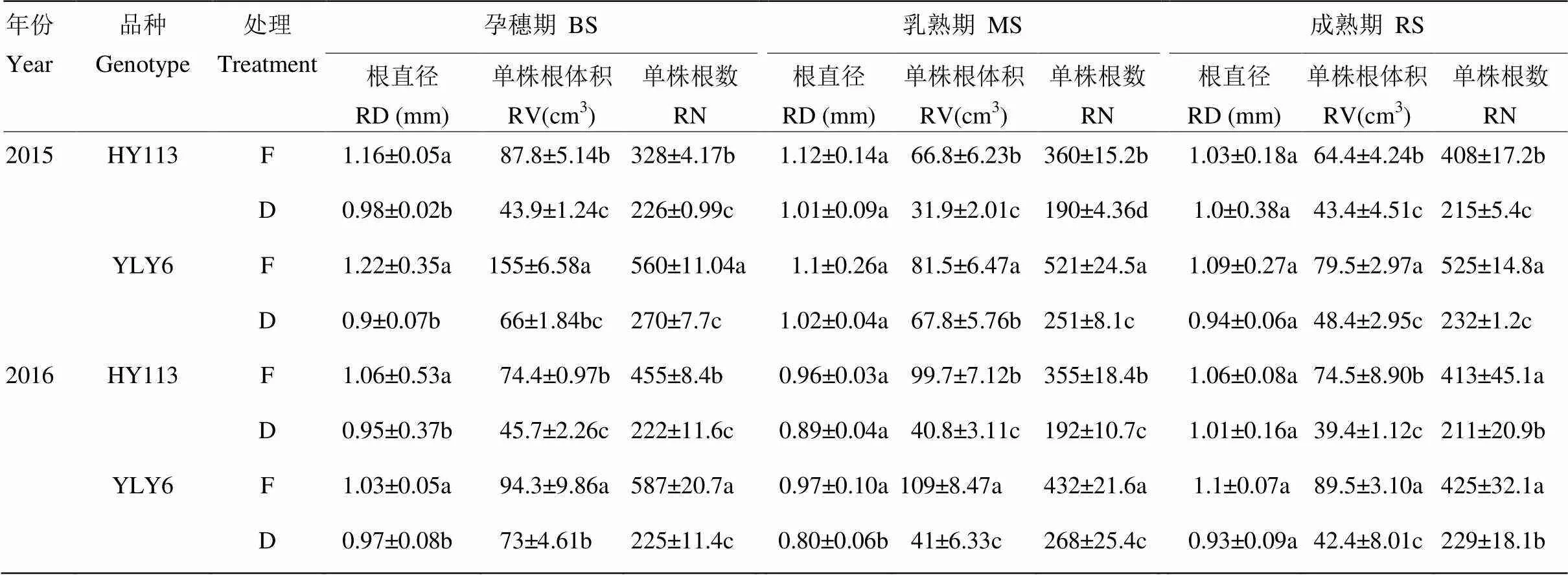
表3 淹灌(F)和干旱(D)处理下节水抗旱稻旱优113号(HY113)与高产水稻扬两优6号(YLY6)的根系形态指标
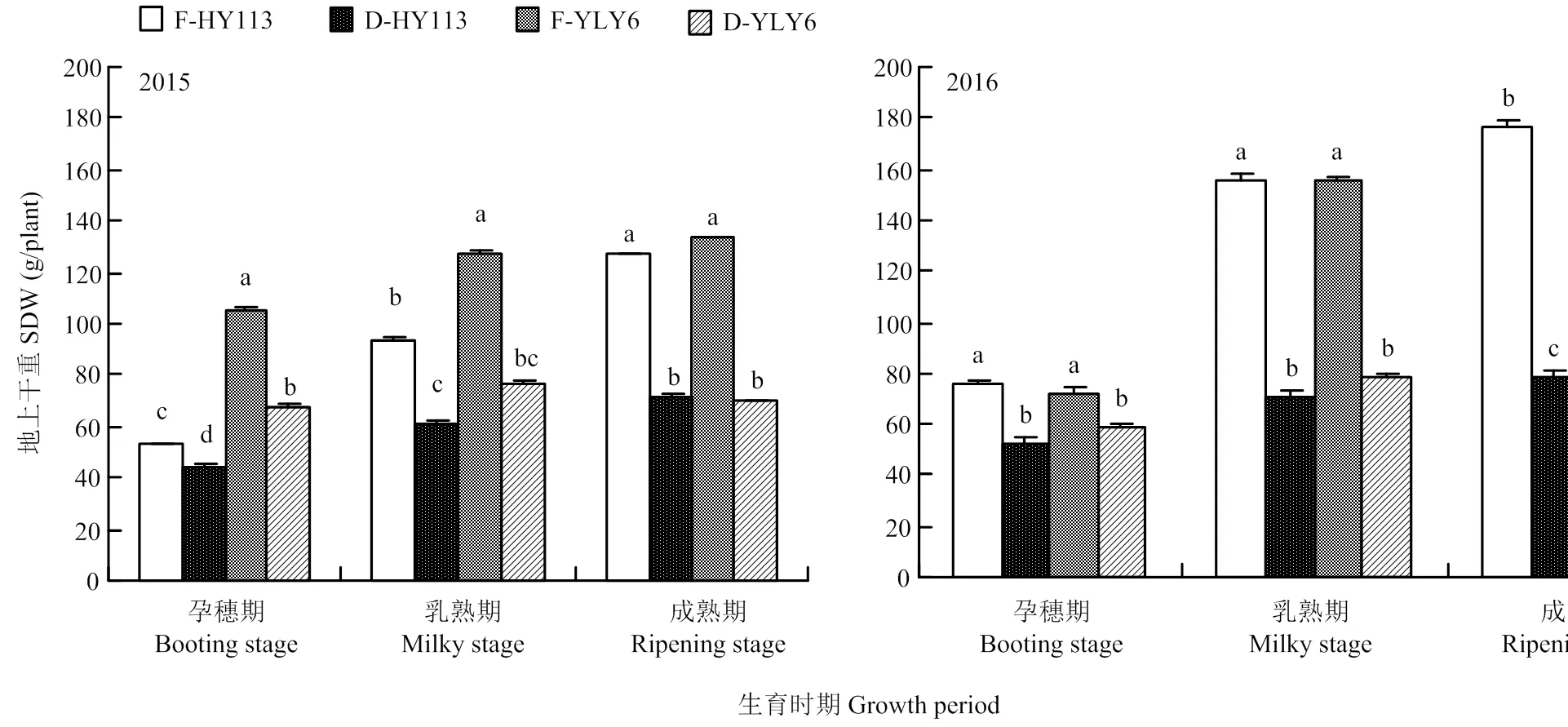
图8 淹灌(F)和干旱处理(D)下旱优113号(HY113)和扬两优6号(YLY6)在不同生育期的地上干重
如图9所示,干旱处理导致水稻的根系干重显著降低。节水抗旱稻HY113的根系干重显著低于高产水稻YLY6,表明HY113根系生物量积累比YLY6少,这与其单株根体积和根数较少有关。
干旱处理对根冠比的影响因品种而异。干旱胁迫下节水抗旱稻HY113的根冠比显著降低,比如2016年孕穗期的降幅最大,由0.18降为0.12;而高产水稻YLY6的根冠比无显著变化。表明干旱胁迫下HY113减少了同化物向根系的分配比例,将干物质较多地留在植株地上部(图10)。
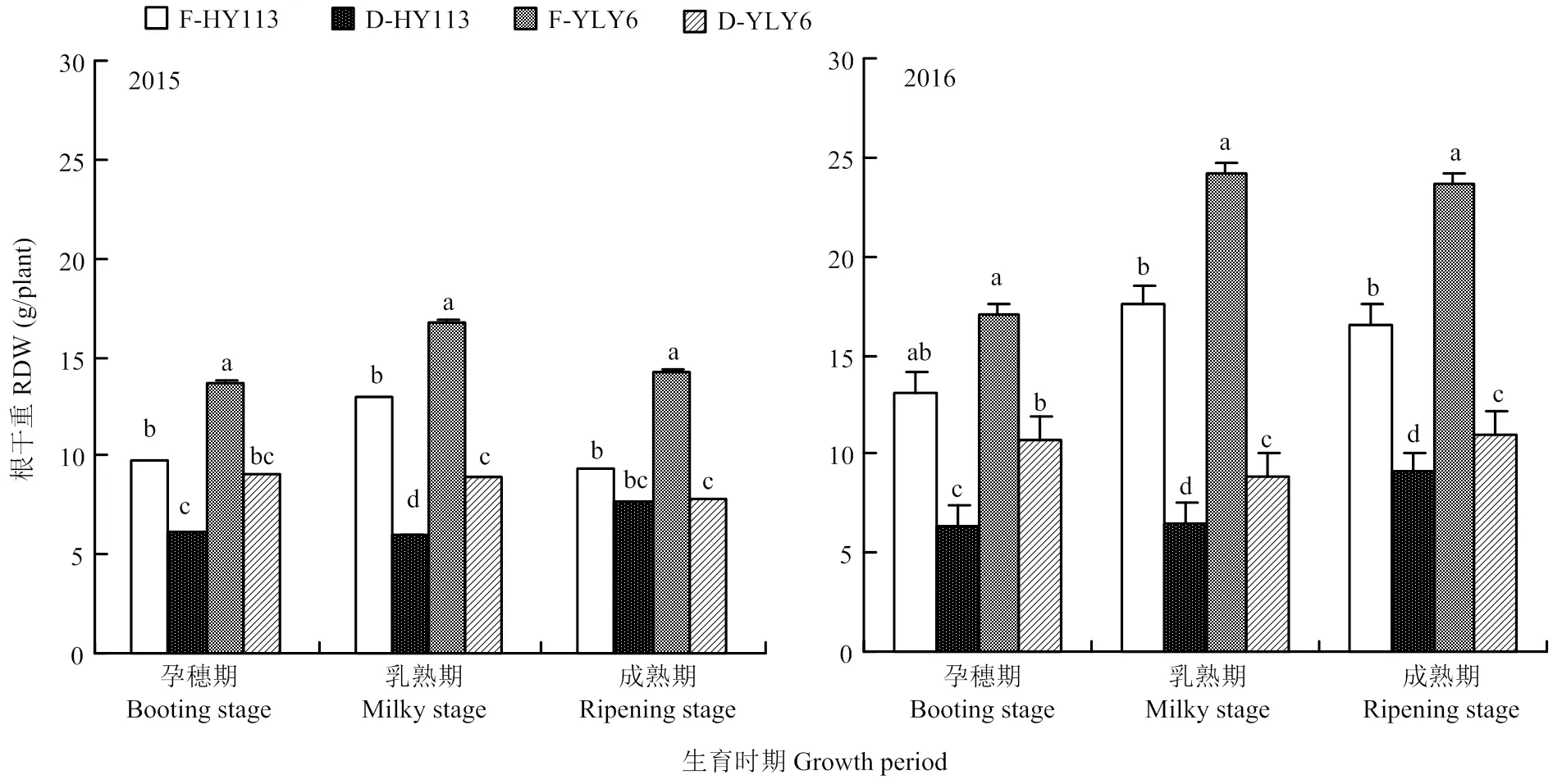
图9 淹灌(F)和干旱处理(D)下旱优113号(HY113)和扬两优6号(YLY6)在不同生育期的根系干重
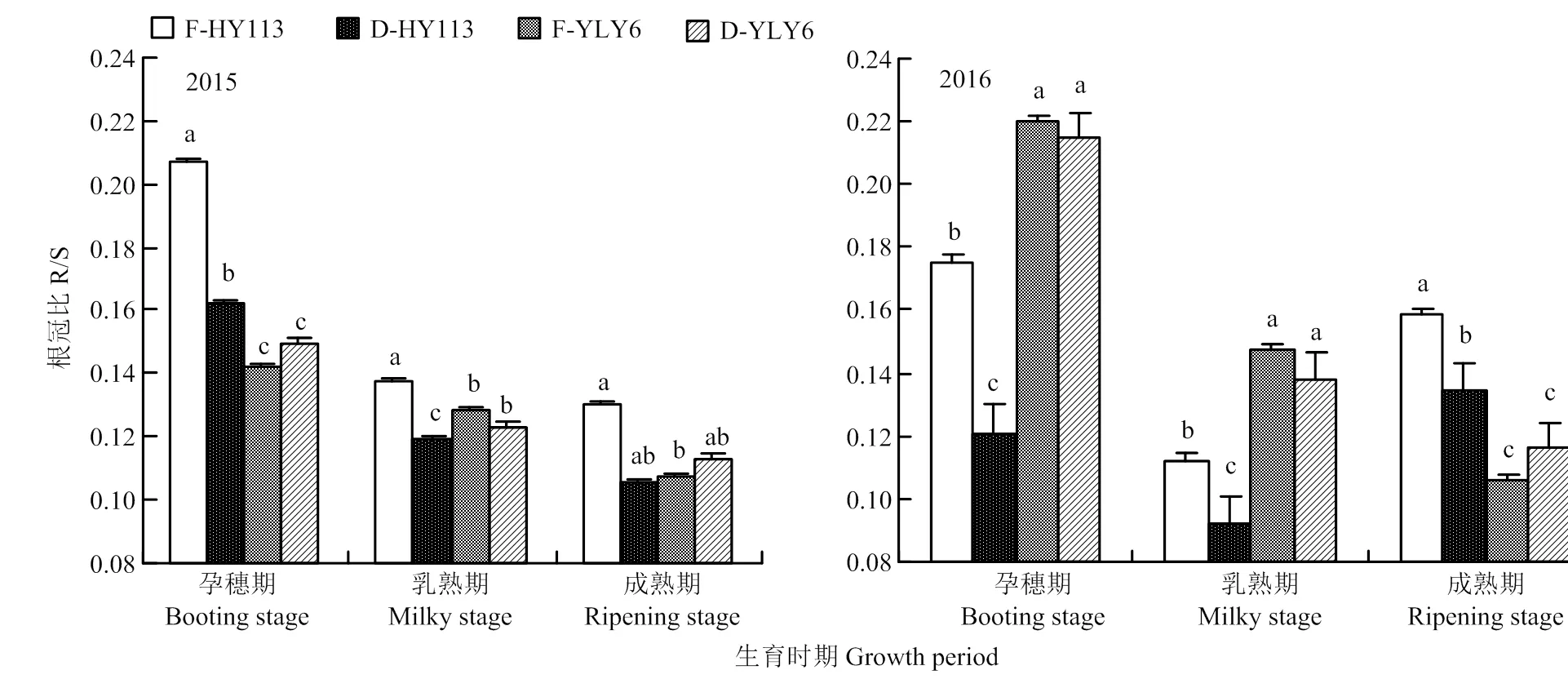
图10 淹灌(F)和干旱处理(D)下旱优113号(HY113)和扬两优6号(YLY6)在不同生育期的根冠比
2.6 各指标与成熟期生物量的相关性分析
相关性分析结果表明,水稻的根干重、地上部干重、根体积、根系活跃吸收面积与单株生物量积累呈极显著正相关关系,其中地上部干重与单株生物量积累相关性最显著(=0.99,<0.001)。而根系活力与生物量积累显著负相关(=-0.85,<0.01)。根冠比与其他指标均不相关,根系活力与其他指标均呈负相关关系(表4)。
表 4各指标与成熟期生物量的相关性分析
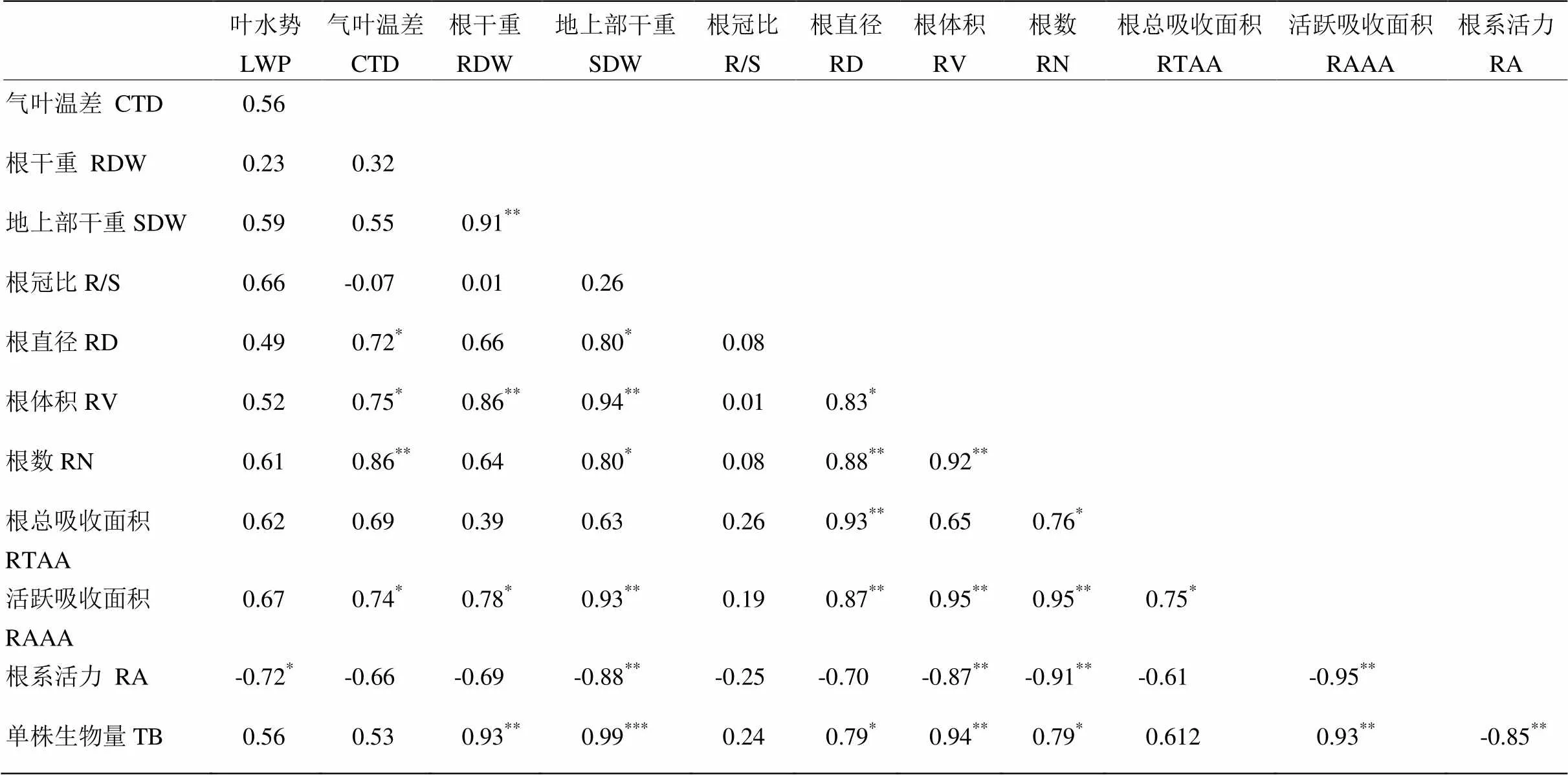
Table 4 Pearson’s correlation coefficients between target traits and rice biomass
CTD:气叶温差;TB:单株生物量。*,**和***分别代表显著性水平为0.05,0.01和0.001
CTD: Canopy temperature depression; TB: Total biomass. *, ** and *** are significant at 5%, 1% and 0.1% levels of probability, respectively
主成分分析结果表明,第一主成分可以解释59.4%的变异,第二主成分可以解释22.6%的变异(图11)。图中离散的点为试验处理,即旱优113号-淹灌(HY113-F)、旱优113号-干旱(HY113-D)、扬两优6号-淹灌(YLY6-F)、扬两优6号-干旱(YLY6-D)。由图10所示,HY113在淹灌与干旱处理间差异显著,但其显著性低于YLY6在两处理间的差异。根系与地上部的干物质积累是显著影响水稻单株生物量积累的主要因素。结果表明提高根体积和根系活跃吸收能力有助于水稻生物量的积累。
3 讨论
植物在遭遇水分亏缺时,最早受到影响的是根系,土壤水分含量下降,根吸收的水分含量也降低,直接影响到根系和地上部的生长发育[21-22]。根系总吸收面积和活跃吸收面积是反映根系对水分和养分吸收能力的重要指标,持续的水分亏缺明显降低水稻各生育期根系总吸收面积与活跃吸收面积的大小[10]。本研究也得到了相似的结果,干旱处理显著降低了水稻的根系吸收能力。此外,本研究还发现干旱胁迫下节水抗旱稻的根系总吸收面积和活跃吸收面积的降低幅度比高产水稻小,并且不同生育期均表现一致(表2)。表明干旱缺水时HY113的根系仍具有较强吸收能力,从而保证植株内水分和养分吸收和供应。
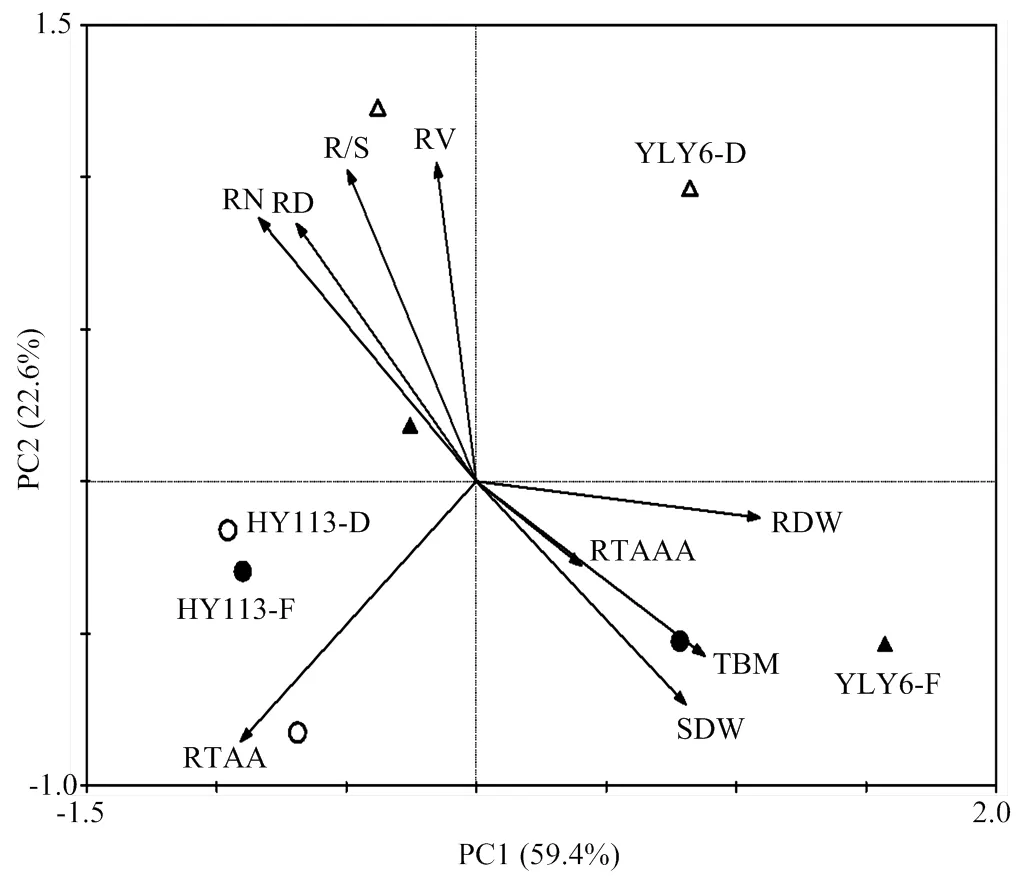
第一主成分可以解释59.4%的变异,第二主成分可以解释22.6%的变异
根系活力是反映根系的生长发育、代谢强度等状况的重要指标[23-24]。本研究发现干旱胁迫能提高水稻的根系活力,再次验证了前人的研究结果[25-26]。罗利军等[27-28]认为节水抗旱稻在水分亏缺下仍能维持较好的根系生理功能,本研究结果也表明除2015年孕穗期外,在干旱胁迫下节水抗旱稻HY113的根系活力在孕穗期和乳熟期均显著高于高产水稻YLY6(图5)。水稻根系的活力是随着生育进程而不断变化的,与根系的呼吸作用有着密切的联系,强的根系活力对后期延缓衰老,促进光合产物的形成及运输,进一步提高产量有重要作用[29-30]。
已有研究发现,水分亏缺使水稻根系的不定根数与根体积都显著降低,孕穗期根系直径显著降低[31-33]。本研究发现干旱胁迫可导致单株根数和根体积显著降低,但根直径未必有显著变化,根直径的变化可能因品种而异,也可能还受到生育期等其他因素的影响(表3)。本试验使用的两个品种的根直径无显著差异,节水抗旱稻HY113的根体积明显小于高产水稻YLY6,主要是因为HY113的根数较少。
土壤水分持续亏缺,导致水稻植株的干物质积累减少[22],本研究发现其直接原因是水分亏缺时,水稻根系活跃吸收面积减少,减缓了根系的与代谢强度,从而导致根数减少、根系干重降低;同时地上部的生长与根系对吸收的水分与养分的吸收能力密切相关,因而会进一步降低地上部的干物质积累。此外,水分亏缺使叶片水势降低,引起叶片气孔开度减少或部分关闭,导致叶片的光合作用减弱,从而引起水稻生物量积累的减少。水稻品种HY113和YLY6相比,2015年与2016年地上生物量的差异性不一致,主要可能是两年的气温与光照等气候因素不一致造成的,但在干旱胁迫下HY113各生育期的地上部生物量的降低幅度均小于YLY6,表明节水抗旱稻HY113在缺水时维持较高地上生物量的能力较强。
本研究结果表明,遭遇干旱胁迫时HY113的根系生物量显著低于YLY6,但根系活性明显高于YLY6,表明节水抗旱稻HY113的根系效率较高。与淹灌相比,HY113的根冠比在干旱胁迫下显著降低,而YLY6的根冠比基本维持不变,并且两年的试验结果基本一致。这与一些研究发现干旱胁迫后植株根冠比增大的结果不一致[34-36],植株出现不同表现应该是因为不同试验中所用水稻品种特性和实施干旱处理时间(包括处理时期和累计处理时间)不同引起的。在遭遇干旱胁迫时,HY113仍具有较高的根系活力,能促进光合产物的形成及运输以保证植株的正常生长;同时HY113通过根冠比的减小,使得光合产物较多的留在了植株地上部分,有利于维持较高的籽粒产量。
4 结论
土壤水分亏缺导致节水抗旱稻和高产水稻品种的地上、地下的干物质积累量均显著降低,其中根系干物质显著降低的主要原因是根数目和根体积的减少。节水抗旱稻HY113的单株总干物质积累量显著低于高产水稻YLY6,主要是其根系干物质积累量较少导致的。与YLY6相比,HY113的根量较少,但其HY113的根系活力与根系吸收能力显著高于YLY6,较强的根系活力能促进水分吸收、光合产物的形成及运输。并且,水分亏缺下节水抗旱稻HY113的根冠比减小,使得更多的干物质留在地上部分供给籽粒灌浆,有利于植株在干旱逆境下维持较高的籽粒产量。
[1] 罗利军, 梅捍卫, 余新桥, 刘鸿艳, 冯芳君. 节水抗旱稻及其发展策略. 科学通报, 2011, 56(11): 804-811.
LUO L J, MEI H W, YU X Q, LIU H Y, FENG F J. Water-saving and drought-resistant rice and its development strategy., 2011, 56(11): 804-811. (in Chinese)
[2] Cassman K G, Dobermann A, Waiters D T. Meeting cereal demand while protecting natural resources and improving environmental quality.2003, 28(1): 315-358.
[3] Tilman D K, Cassman K G, Matson P A. Agricultural sustainability and intensive production practices.2002, 418(8): 671-677.
[4] 茆智. 水稻节水灌溉及其对环境的影响. 中国工程科学, 2002, 4(7): 8-16.
Mao Z. Rice water-saving irrigation and its impact on environment., 2002, 4(7): 8-16. (in Chinese)
[5] 董桂春, 王余龙, 王坚刚, 单玉华, 马爱京, 杨洪, 张传胜, 蔡惠荣. 不同类型水稻品种间根系性状差异. 作物学报,2002, 28(6): 749-755.
DONG G C, WANG Y L, WANG J G,Shan Y H, MA A J, YANG H, ZHANG C S, CAI H R. Study on the differences of root traits between various types of varieties in rice(L. )., 2002, 28 (6): 749-755. (in Chinese)
[6] 杨建昌. 水稻根系形态生理与产量、品质形成及养分吸收利用的关系. 中国农业科学, 2011, 44(1): 36-46.
YANG J C. Relationships of rice root morphology and physiology with the formation of grain yield and quality and the nutrient absorption and utilization., 2011, 44(1): 36-46. (in Chinese)
[7] 刘骞, 齐宏伟. 水稻根系研究进展. 种子世界, 2009(11): 33-35.
LIU Q, QI H W. Advances in rice root research., 2009(11): 33-35. (in Chinese)
[8] LUO L J. Breeding for water-saving and drought-resistance rice (WDR) in China., 2010, 61(13): 3509-3517.
[9] CHU G , CHEN T T, WANG Z Q, YANG J C, ZHANG J H. Reprint of “Morphological and physiological traits of roots and their relationships with water productivity in water-saving and drought- resistant rice”2014, 165(6): 36-48.
[10] 张耗, 剧成欣, 陈婷婷, 曹转勤, 王志琴, 杨建昌. 节水灌溉对节水抗旱水稻品种产量的影响及生理基础. 中国农业科学, 2012, 45(23): 4782-4793.
ZHANG H, JU C X, CHEN T T, CAO Z Q, WANG Z Q, YANG J C. Effect of water-saving irrigation on the grain yield of water-saving and drought-resistance rice and its physiological bases., 2012, 45(23): 4782-4793. (in Chinese)
[11] 张耗, 黄钻华, 王静超, 王志琴, 杨建昌. 江苏中籼水稻品种演进过程中根系形态生理性状的变化及其与产量的关系. 作物学报, 2011, 37(6): 1020-1030.
ZHANG H, HUANG Z H, WANG J C, WANG Z Q, YANG J C. Changes in morphological and physiological traits of roots and their relationships with grain yield during the evolution of mid-seasonrice cultivars in Jiangsu province., 2011, 37(6): 1020-1030. (in Chinese)
[12] 褚光, 周群, 薛亚光, 颜晓元, 刘立军, 杨建昌. 栽培模式对杂交粳稻常优5号根系形态生理性状和地上部生长的影响. 作物学报, 2014, 40(7): 1245-1258.
CHU G, ZHOU Q, XUE Y G, YAN X Y, LIU L J, YANG J C. Effects of cultivation patterns on root morph-physiological traits and aboveground development ofhybrid rice cultivar changyou 5., 2014, 40(7): 1245-1258. (in Chinese)
[13] 杨彩玲, 刘立龙, CHUNG N T, 秦华东, 赵泉, 陈德威, 徐世宏, 黄敏, 江立庚. 土壤水分对免耕水稻根系生长的影响. 华中农业大学学报, 2016, 35(1): 8-16.
YANG C L, LIU L L, CHUNG N T, QIN H D, ZHAO Q, CHEN D W, XU S H, HUANG M, JIANG L G. Effects of soil moisture on root growth of no-tillage rice., 2016, 35(1): 8-16. (in Chinese)
[14] 陈达刚, 周新桥, 李丽君, 刘传光, 张旭, 陈友订. 华南主栽高产籼稻根系形态特征及其与产量构成的关系. 作物学报, 2013, 39(10): 1899-1908.
CHEN D G, ZHOU X Q, LI L J, LIU C G, ZHANG X, CHEN Y D. Relationship between root morphological characteristics and yield components of major commercialrice in South China.2013, 39(10): 1899-1908. (in Chinese)
[15] 徐国伟, 吕强, 陆大克, 王贺正, 陈明灿. 干湿交替灌溉耦合施氮对水稻根系性状及籽粒库活性的影响. 作物学报, 2016, 42(10): 1495-1505.
XU G W , LÜ Q, LU D K, WANG H Z, CHEN M C. Effect of wetting and drying alternative irrigation coupling with nitrogen application on root characteristic and grain-sink activity.2016, 42(10): 1495-1505. (in Chinese)
[16] 蔡昆争, 骆世明, 段舜山. 水稻根系在根袋处理条件下对氮养分的反应. 生态学报, 2003, 23(6): 1109-1116.
CAI K Z, LUO S M, DUAN S S. The response of the rice root system to nitrogen conditions under root confinement., 2003, 23(6): 1109-1116. (in Chinese)
[17] PENG S B, HUANG J L, ZHONG X H, YANG J C, WANG G H, ZOU Y B, ZHANG F S, ZHU Q S, Buresh R J, Witt C. Challenge and opportunity in improving fertilizer-nitrogen use efficiency of irrigated rice in China.2002, 1(7): 776-785.
[18] 胡艳丽, 毛志泉, 沈向, 张黎黎. 影响农作物根系冗余影响因子探讨. 现代农业科技, 2008(23): 13-15.
HU Y L, MAO Z Q, SHEN X, ZHANG L L. Discussion on influencing factors of crop roots redundancy., 2008(23): 13-15. (in Chinese)
[19] 蔡昆争, 骆世明, 段舜山. 水稻群体根系特征与地上部生长发育和产量的关系. 华南农业大学学报, 2005, 25(6): 1-4.
CAI K Z, LUO S M, DUAN S S. The relationship between roots system of rice and aboveground characteristics and yield.2005, 25(6): 1-4. (in Chinese)
[20] MA S C, LI F M, XU B C, HUANG Z B. Effect of lowering the root/shoot ratio by pruning roots on water use efficiency and grain yield of winter wheat., 2010, 115(2): 158-164.
[21] YANG J, ZHANG H, ZHANG J. Root morphology and physiology in relation to the yield formation of rice., 2012, 11(6): 920-926.
[22] ZHANG H, XUE Y G, WANG Z Q, YANG J C, ZHANG J H. Morphological and physiological traits of roots and their relationships with shoot growth in “super” rice., 2009, 113(1): 31-40.
[23] 蔡永萍, 杨其光, 黄义德. 水稻水作与旱作对抽穗后剑叶光合特性、衰老和根系活性的影响. 中国水稻科学, 2000, 14(4): 219-224.
CAI Y P, YANG Q G, HUANG Y D. Effects of rice cultivated under paddy and upland condition on photosynthesis and senescence of flag leaf and activity of root system after heading., 2000, 14(4): 219-224. (in Chinese)
[24] 程兆伟, 邹应斌, 刘武. 水稻根系研究进展. 作物研究, 2006, 20(s1): 504-507.
CHENG Z W, ZOU Y B, LIU W. Advances of research on rice root systems., 2006, 20(s1): 504-507. (in Chinese)
[25] 蔡昆争, 吴学祝, 骆世明, 王维. 不同生育期水分胁迫对水稻根系活力、叶片水势和保护酶活性的影响. 华南农业大学学报, 2008, 29(2): 7-10.
CAI K Z, WU X Z, LUO S M, WANG W. Effects of water stress at different growth stages on root activity, leaf water potential and protective enzymes activity in rice., 2008, 29(2): 7-10. (in Chinese)
[26] 郝树荣, 郭相平, 张展羽, 王为木. 水稻根冠功能对水分胁迫及复水的补偿响应. 农业机械学报, 2010, 41(5): 52-55.
HAO S R, GUO X P, ZHANG Z Y, WANG W M. Compensation effects of water stress and re-watering on the function of root shoot., 2010, 41(5): 52-55. (in Chinese)
[27] LUO L J, MEI H W, YU X Q, LIU H Y, FENG F J. Water-saving and drought-resistance rice and its development strategy., 2011, 56(11): 804-811.
[28] ZOU G H, LIU H Y, MEI H W, LIU G L, YU X Q, LI M S, WU J H, CHEN L, LUO L J. Screening for drought resistance of rice recombinant inbred populations in the field., 2007, 49(10): 1508-1516.
[29] 孙静文, 陈温福, 臧春明, 王彦荣, 吴淑琴. 水稻根系研究进展. 沈阳农业大学学报, 2002, 33(6): 174-180.
Sun J W, Chen W F, Zang C M, Wang Y R, Wu S Q. Advances of research on rice root systems.y, 2002, 33(6): 174-180. (in Chinese)
[30] 腾胜, 曾大力, 钱前, 国广泰史, 藤本宽, 黄大年, 朱立煌. 水稻根系活力的遗传分析. 中国水稻科学, 2002, 16(2): 119-123.
TENG S, ZENG D L, QIAN Q, GUO G T S, TENG B K, HUANG D N, ZHU L H. Genetic analysis of root activity in rice ().2002, 16(2): 119-123. (in Chinese)
[31] Kadam N N, Yin X, Bindraban P S, Struik P C, Jagadish K S V. Does morphological and anatomical plasticity during the vegetative stage make wheat more tolerant of water deficit stress than rice., 2015, 167(4): 1389-1401.
[32] Farooq M, Wahid A, Kobayashi N, Fujita D, Basra S M A. Plant drought stress: Effects, mechanisms and management.2009, 29(1): 153-188.
[33] 明东风, 袁红梅, 王玉海, 宫海军, 周伟军. 水分胁迫下硅对水稻苗期根系生理生化性状的影响. 中国农业科学, 2012, 45(12): 2510-2519.
MING D F, YUAN H M, WANG Y H, GONG H J, ZHOU W J. Effects of silicon on the physiological and biochemical characteristics of roots of rice seedlings under water stress., 2012, 45(12): 2510-2519. (in Chinese)
[34] 杨长明, 杨林章, 欧阳竹. 不同养分与水分管理对水稻植株根系形态及其活力的影响. 中国生态农业学报, 2004, 12(4): 82-85.
YANG C M, YANG L Z, OU Y Z. Effects of different nutrient and water regimes on the morphology and activity of rice root system., 2004, 12(4): 82-85. (in Chinese)
[35] YANG J C, ZHANG J H. Root morphology and physiology in relation to the yield formation of rice., 2012, 11(6): 920-926.
[36] ZHANG H, XUE Y G, WANG Z Q, YANG J C, ZHANA J H. An alternate wetting and moderate soil drying regime improves root and shoot growth in rice., 2009, 49(6): 2246-2260.
(责任编辑 杨鑫浩)
Root Growth Responses to Soil Water Deficit for a Water-Saving and Drought-Resistant Rice Genotype Hanyou113
BU HongYing1,SONG WeiZhou1, CAO CouGui1,2, LI Ping1,2
(1College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070;2Hubei Collaborative Innovation Center for Grain Industry, Yangtze University, Jingzhou 434025, Hubei)
The objective of this paper is to reveal root morphological and physiological changing characteristics of water-saving and drought-resistant rice under soil water deficit condition, and to illuminate its specific physiological basis of water-saving and drought-resistance.Pot experiments were conducted in 2015 and 2016, and one water-saving and drought-resistant rice genotype (HY113) and one high-yielding rice genotype (YLY6) were used under two water treatments, including flooding irrigation and drought treatment. The drought effects on root morphological and physiological traits of the two rice genotypes and their relationship with aboveground biomass accumulation were studied.Compared with flooding irrigation, drought stress significantly reduced the dry matter accumulation of the two rice genotypes, and the root-shoot ratio of HY113 was also decreased significantly (from 0.18 to 0.12), while the root-shoot ratio of YLY6 was not changed. Under drought stress, the root activity of the two rice genotypes both increased significantly. In 2015 and 2016, the average increase percentage of HY113 was 38.7% at milky stage, while that of YLY6 was 22.8%, and the root activity of HY113 increased from 86 μg·g-1·h-1to 174 μg·g-1·h-1at milky stage in 2015. Drought treatment also significantly reduced the root absorption area of both two genotypes, but the reduction percentage of HY113 was significantly lower than that of YLY6. Compared with high-yielding rice YLY6, HY113 had smaller root number, root volume and root dry weight, and the total dry matter accumulation was also significantly lower, but its root activity and root absorption area were greater.The water-saving and drought-tolerant rice genotype HY113 possesses less root biomass but higher root uptake efficiency, and it could reduce the root-shoot ratio under drought condition to make more dry matter stay in the aboveground part, so that the relatively high grain yield could be maintained.
water-saving and drought-resistant rice; water deficit; root-shoot ratio; root characteristics
2017-02-20;
国家重点研发计划“粮食丰产增效科技创新”专项(2017YFD0301400)
接受日期:2017-09-20
联系方式:补红英,E-mail:764992312@qq.com。通信作者李萍,E-mail:sleep1022@mail.hzau.edu.cn
