七种野生铁线莲属植物核型分析
2017-11-24王娜王奎玲刘庆华刘庆超
王娜,王奎玲,刘庆华,刘庆超
(青岛农业大学园林与林学院,山东 青岛 266109)
七种野生铁线莲属植物核型分析
王娜,王奎玲,刘庆华,刘庆超*
(青岛农业大学园林与林学院,山东 青岛 266109)
铁线莲属植物种间在形态、香味、花量、绿期及生活型等方面差异较大,遗传背景也相对复杂,因此在杂交育种前了解其核型特征意义重大。采用染色体常规压片法,对东方铁线莲、大花铁线莲、大叶铁线莲、褐毛铁线莲、棉团铁线莲、齿叶铁线莲和东北铁线莲7种野生铁线莲属植物的倍性及核型进行分析,结果表明,1) 7种野生铁线莲中除东北铁线莲为四倍体(2n=4x=32)外,另外6种均为二倍体(2n=2x=16),染色体基数为8。染色体类型有中部(m)、亚中部(sm)、亚端部(st)、端部(t)4种,其中大花铁线莲和大叶铁线莲的核型公式为2n=2x=16=5m+2st+t,褐毛铁线莲的核型公式为2n=2x=16=5m+3st,棉团铁线莲的核型公式为2n=2x=16=5m+2st(SAT)+t,齿叶铁线莲的核型公式为2n=2x=16=5m(SAT)+2st+t,东方铁线莲的核型公式为2n=2x=16=5m(SAT)+sm(SAT)+2st,东北铁线莲的核型公式为2n=4x=32=5m+2st+t。东方铁线莲、棉团铁线莲、齿叶铁线莲的短臂上带有随体。2) 核型不对称系数为62.27%~66.77%,核型类型均为2A型,表明7种野生铁线莲属植物的进化程度均较原始。其中,褐毛铁线莲和齿叶铁线莲为首次报道。
铁线莲属;染色体;核型分析
物种的核型反映了其染色体水平的整体特征,研究物种的核型有助于判断物种间的亲缘关系,揭示其遗传进化的过程和机制[6]。国内已有学者对不同地区野生铁线莲的某些种及品种做过研究报道。龚维忠等[7]对东北及北京地区的11种铁线莲进行核型分析研究。张镱锂等[8-9]先后对国内13种铁线莲进行核型分析,并探讨了某些种的亲缘关系及其分类。杨亲二等[10]对粗齿铁线莲(Clematisargentilucida)和扬子铁线莲(Clematisganpiniana)进行过核型报道。彭绿春等[11]对金毛铁线莲(Clematischrysocoma)等4种野生铁线莲属植物的核型进行了详细报道。盛璐[12-13]陆续对铁线莲属23个种进行了染色体核型分析,并对其进行分类。此外,Sheidai等[14]对银叶铁线莲(Clematisdelavayi)等3种野生铁线莲属植物进行核型分析及分类,发现不同地区的同种铁线莲核型有所不同。同时,段青等[15]也曾提到同一物种不同居群在形态和分子方面有较高的遗传多样性,居群间的遗传分化较大。相较于中国各地区丰富的野生铁线莲属植物资源,仅这些居群的染色体资料还不够,且对于有些种类如大花铁线莲(Clematispatens)[7,13]、东北铁线莲(Clematisternifloravar.mandshurica)等[8]前人的研究结果也不尽相同。
远缘杂交在高等植物的进化过程中起着重要的作用[16]。如今铁线莲属植物在园林绿化应用方面的推广价值很高,野生种或品种虽在适应性等其他特性上占有优势,但这种优势往往不能集中。因此,通过远缘杂交等途径集合铁线莲属中各种的优良特性,培育出花量覆盖率大、单花大、花香、绿期长的新品种,并利用雄蕊瓣化的特性改良单瓣花以得到具有优质观赏价值的重瓣花。但为避免前人研究的不确定性对今后的实验带来困扰,为积累铁线莲属植物在染色体层面的基本特征状况,本研究选择7种在北方适应性较强又均存在各自优良品质的铁线莲属植物,如花香浓郁的东北铁线莲、大花型的大花铁线莲、钟状花型的褐毛铁线莲(Clematisfusca)、花量覆盖率大的齿叶铁线莲(Clematisserratifolia)、半灌木型的大叶铁线莲(Clematisheracleifolia)以及雄蕊瓣化较多的棉团铁线莲(Clematishexapetala)等作为种质资源创新的材料,并对其染色体核型进行分析,为后续的杂交育种提供可靠的细胞学依据,同时也为其遗传变异、系统分类、品种特性以及栽培利用等方面奠定科学基础。
1 材料与方法
1.1试验材料
7种野生铁线莲属植物材料(表1)自野外采集后,于2014年12月统一栽植于青岛农业大学试验地内。除东方铁线莲(Clematisorientalis)是以种子萌发的幼嫩根尖为材料外,其他皆于2016年4月取盆栽植株新长出的幼嫩侧根的根尖为材料。
1.2试验方法
采用染色体常规压片法[17],于早10:00-11:00切取新长出长度为1~2 cm的根尖,蒸馏水清洗后,在4 ℃条件下以0.1%的秋水仙素溶液处理4 h,再经蒸馏水清洗后用卡诺固定液4 ℃冰箱中固定20 h,之后以95%、85%各20 min的乙醇梯度浸泡后转70%的乙醇中置于4 ℃的冰箱内保存备用。染色体制片时,经酸解(1 mol/L HCl,60 ℃水浴10 min)和酶解(2.5%混合酶,4 ℃下1 h)两次解离,用蒸馏水清洗干净,取根尖顶端2 mm的乳白色分生区于载玻片上,并用滤纸在一侧吸干水分,滴改良卡宝品红溶液染色10 min,盖上盖玻片,轻敲,压片,在Oympus显微镜下镜检,拍照。

表1 供试材料Table 1 Materials used for this study
1.3数据分析
从7种野生铁线莲染色体制片中分别选出30个较好的分裂相,观察并统计其染色体的数目。并从中选5个清晰且分散最好的分裂相,以Photoshop CS3抠图对比配对,Image J测其染色体的臂长,以这5个细胞各项指标的均值为各参数值计算。染色体核型分析采用李懋学等[18]的分类标准。核型对称与不对称分类参照Stebbins[19]的标准。着丝粒位置的确定参照Levan等[20]的二点四区系统标准。核型不对称系数(asymmetric karyotype coefficient, AKC)按Arano[21]的方法计算。
核型不对称系数(AKC)=(染色体长臂总长/全组染色体总长)×100%
2 结果与分析
7种铁线莲材料的染色体形态、数目及核型图见图1。核型分析的主要染色体参数列于表2和表3,并分述如下。
东方铁线莲(C.orientalis):染色体数2n=2x=16,为二倍体。核型公式为2n=16=5m(SAT)+sm(SAT)+2st。染色体相对长度为10.34%~14.71%,臂比的变化范围为1.17~6.07,平均臂比为2.31,最长与最短染色体的比值为1.42,核型不对称系数为62.27%,着丝点指数为36.55%,其核型属于“2A”型。第5、6对染色体的短臂上存在次缢痕,连有随体。
大花铁线莲(C.patens):染色体数2n=2x=16,为二倍体。核型公式为2n=16=5m+2st+t,染色体相对长度为9.38%~15.27%,臂比的变化范围为1.34~8.50,平均臂比为3.22,最长与最短染色体的比值为1.63;核型不对称系数为66.77%;着丝点指数为31.19%,其核型属于“2A”型。
东北铁线莲(C.ternifloravar.mandshurica):染色体数2n=4x=32,为四倍体。核型公式:2n=32=5m+2st+t,染色体的相对长度变化范围为8.70%~15.93%,臂比的变化范围为1.07~9.38,平均臂比为3.24,最长与最短染色体的比值为1.83,核型不对称系数为63.99%,着丝点指数为33.04%,属于“2A”型。
褐毛铁线莲(C.fusca):染色体数2n=2x=16,为二倍体。核型公式为2n=2x=16=5m+3st,染色体相对长度为9.47%~14.88%,臂比的变化范围为1.16~5.38,平均臂比为2.49,最长染色体与最短染色体的比值为1.57;核型不对称系数为63.02%,着丝点指数为34.8%,属于“2A”型。
大叶铁线莲(C.heracleifolia):染色体数2n=2x=16,为二倍体。核型公式为2n=2x=16=5m+2st+t,染色体相对长度为8.37%~14.78%,臂比的变化范围为1.17~11.67,平均臂比为3.32,最长染色体与最短染色体的比值为1.76,核型不对称系数为64.77%,着丝点指数为33.14%,属于“2A”型。
棉团铁线莲(C.hexapetala):染色体数2n=2x=16,二倍体。核型公式为2n=2x=16=5m+2st(SAT)+t,染色体相对长度为9.98%~14.77%,臂比的变化范围为1.10~7.64,平均臂比为2.87,最长染色体与最短染色体的比值为1.47;核型不对称系数为64.49%,着丝点指数为33.77%,属于“2A”型。第7对染色体的短臂上连有随体。
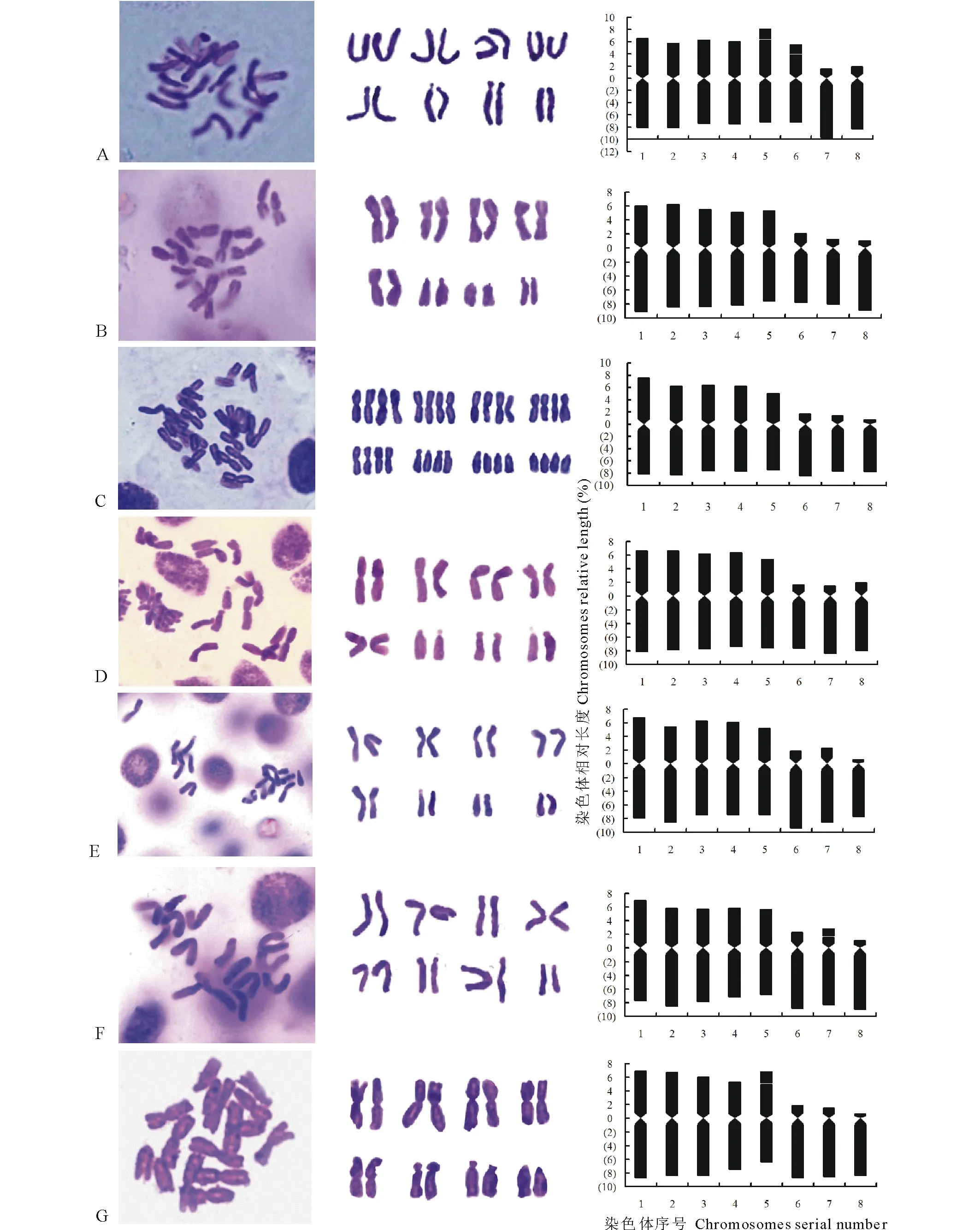
图1 7种铁线莲中期染色体形态及核型模式图Fig.1 Somatic metaphase photomicrographs and idiogram of seven species of Clematis A:东方铁线莲C. orientalis;B:大花铁线莲C. patens;C:东北铁线莲C. terniflora var. mandshurica;D:褐毛铁线莲C. fusca;E:大叶铁线莲C. heracleifolia;F:棉团铁线莲C. hexapetala;G:齿叶铁线莲C. serratifolia.
齿叶铁线莲(C.serratifolia):染色体数2n=2x=16,二倍体。核型公式为2n=2x=16=5m(SAT)+2st+t,染色体相对长度为9.14%~15.77%,臂比的变化范围为1.24~11.75,平均臂比为3.51,最长染色体与最短染色体的比值为1.73,核型不对称系数为65.41%,着丝点指数为32.3%,属于“2A”型。第5对染色体的短臂上有随体。
注:相对长度Relative length:长臂Long arm+短臂Short arm=全长Total.*:随体,随体长度不计入染色体长度;m:中部着丝粒染色体;sm:亚中部着丝粒染色体;st:亚端部着丝粒染色体;t:端部着丝粒染色体。下同。
Notes:* satellite, chromosome length exclude satellites. m: Metacentric chromosome;sm: Submetacentric chromosome;st: Subtelocentric chromosome;t: Telocentric chromosome. The same below.
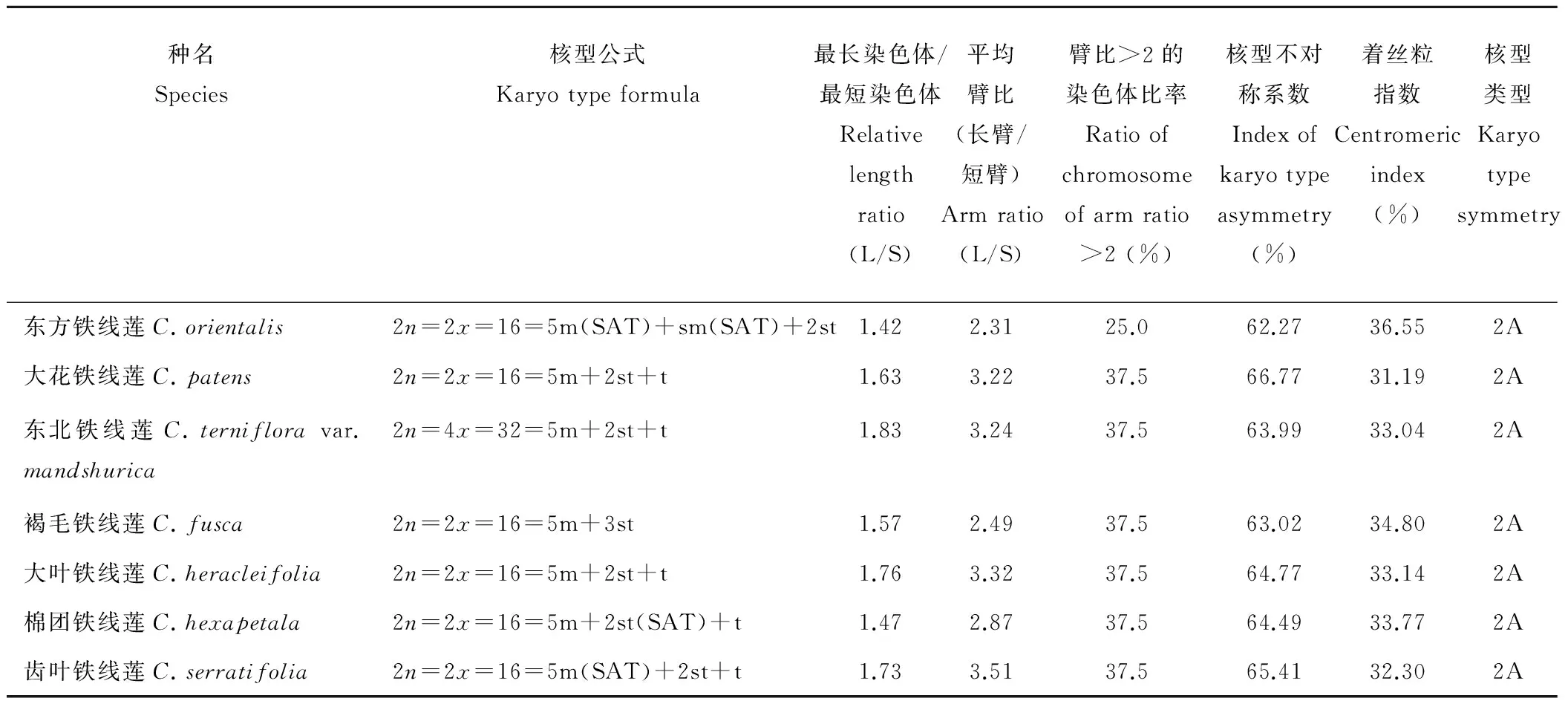
表3 7种野生铁线莲主要核型特征比较Table 3 The comparison on significant characteristics of chromosome of seven species of Clematis
3 讨论
本研究对7种野生铁线莲的核型进行了分析。首次报道了褐毛铁线莲和齿叶铁线莲的核型,均为2A型。其中褐毛铁线莲的核型公式为2n=2x=16=5m+3st,不含随体,核型不对称系数为63.02%;齿叶铁线莲的核型公式为2n=2x=16=5m(SAT)+2st+t,第5对染色体的短臂上有随体,核型不对称系数为65.41%。与Sheidai等[14]的研究相比较,本研究中东方铁线莲的核型公式及类型与其不一致,Sheidai等[14]研究的东方铁线莲取自Roodbar地区,核型为2B型。造成这种差异的原因可能是环境和地理因素的不同,周其兴等[22]和尚宝龙等[23]对核型的研究中也提到B型染色体通常具有较强的适应力和竞争力,相较于A型不对称性更强,进化程度更高。同时,从生境条件来看,伊朗北部Roodbar地区的东方铁线莲位于较高的海拔,具有较强的耐寒冷、抗紫外及其他抗逆能力,通常能够在比较恶劣的环境中生存,这在一定程度上与该植物中存在B型染色体有关,同时也表明东方铁线莲的遗传变异更为活跃。因此,中国新疆地区东方铁线莲的种植资源较为原始,而伊朗北部Roodbar地区的东方铁线莲为了适应环境而更为进化。
大花铁线莲和大叶铁线莲的核型公式与龚维忠等[7]的相悖,且未发现随体,与盛璐[13]的核型公式相同,但由于是不同来源的种质,相同的核型类型下也有着不同的对称性。李懋学等[17]认为在植物界,核型进化的基本趋势是由对称向不对称发展的,处于比较古老或原始的植物,大多具有较对称的核型,而不对称的核型则常见于衍生的或进化较高级的植物中[24]。核型不对称系数可反映不同种质的不对称性,取自南京的比取自辽宁的大叶铁线莲不对称性更低,更为原始。平均臂比与染色体长度比也可从另一角度反映不同铁线莲的不对称性,二者的值越大,核型不对称性越高[24],再综合核型不对称系数,可知取自辽宁的比取自山东的大花铁线莲不对称性更高,更为进化。东北铁线莲是圆锥铁线莲的一个变种,为四倍体(2n=4x=32),与龚维忠等[7]的研究结果相同。
研究发现东方铁线莲、棉团铁线莲和齿叶铁线莲带有随体,且均位于染色体短臂,这一结果符合李懋学等[18]研究中所提到的大部分植物的随体位于短臂的结论,其他4种无随体。但在其他学者的研究中大花和东北铁线莲也有随体存在[7]。关于随体,李真等[25]的研究中提到,由于不同居群的进化过程不同,导致其核型特征特别是随体的数量、大小、分布位置往往存在差异。因此,无论是在核型公式、核型的对称性,还是在有无随体的研究上均能证明由于种源地的不同,其核型会存在细微差别。但这种差别是否会对远缘杂交产生影响,有待于进一步的探讨。
References:
[1] Pei H M, Mao H L. Study on the germplasm resources and fancy application of wildClematisL. in south Gansu province. Chinese Wild Plant Resources, 2004, 23(6): 30-32.
裴会明, 毛浩龙. 甘肃南部野生铁线莲属植物种质资源及观赏应用. 中国野生植物资源, 2004, 23(6): 30-32.
[2] Ma D. Research on Cold Resistance ofClematisheracleifoliain Harbin[D]. Harbin: Northeast Forestry University, 2010.
马迪. 哈尔滨地区大叶铁线莲抗寒性的研究[D]. 哈尔滨: 东北林业大学, 2010.
[3] Jiang L Y. Effect of Na2CO3, NaHCO3Stress on Morphological and Physiological ofClematisheracleifolia[D]. Harbin: Northeast Forestry University, 2011.
姜丽颖. Na2CO3、NaHCO3胁迫对大叶铁线莲形态和生理的影响[D]. 哈尔滨: 东北林业大学, 2011.
[4] Yang L, Yao G Y, Xue H Y,etal. Antitum or activities of total saponins fromClematishexapetalaPall. Acta Chinese Medicine and Pharmacology, 2011, 39(2): 21-24.
杨林, 姚广玉, 薛鸿燕, 等. 棉团铁线莲总皂苷抗肿瘤作用研究. 中医药学报, 2011, 39(2): 21-24.
[5] Feng L F, Weng M Y, Tian F R,etal. Comparasion on total flavonoid content from aerial parts of 11 medicinal plants ofClematisL. Modern Chinese Medicine, 2011, 13(7): 14-18.
冯玲枫, 翁美娅, 田富饶, 等. 11种铁线莲属药用植物地上部分总黄酮含量比较研究. 中国现代中药, 2011, 13(7): 14-18.
[6] Liu H M, Zhi L, Zhao L H,etal. Karyotype analysis of four wildLiliumspecies. Journal of Plant Genetic Resources, 2010, 11(4): 469-473.
刘华敏, 智丽, 赵丽华, 等. 四种野生百合核型分析. 植物遗传资源学报, 2010, 11(4): 469-473.
[7] Gong W Z, Long Y Y, Li M X. Karyotype studies onClematisfrom Beijing China. Journal of Wuhan Botanical Research, 1985, 3(4): 371-379.
龚维忠, 龙雅宜, 李懋学. 北京地区铁线莲属植物的核型研究. 武汉植物学研究, 1985, 3(4): 371-379.
[8] Zhang Y L, He S Y. Chromosome studies on 6 species ofClematisin China. Journal of Wuhan Botanical Research, 1990, 8(2): 115-121.
张镱锂, 贺士元. 国产6种铁线莲的染色体研究. 武汉植物学研究, 1990, 8(2): 115-121.
[9] Zhang Y L. Chromosome studies on 7 species ofClematisin China. Journal of Wuhan Botanical Research, 1991, 9(2): 107-111, 201-202.
张镱锂. 7种铁线莲的染色体研究. 武汉植物学研究, 1991, 9(2): 107-111, 201-202.
[10] Yang Q E, Luo Y B, Hong D Y. A karyotypic study of species in the Ranunculaceae from Hunan. Guihaia, 1994, 14(1): 27-36, 99-101.
杨亲二, 罗毅波, 洪德元. 湖南6种毛茛科植物的核型研究. 广西植物, 1994, 14(1): 27-36, 99-101.
[11] Peng L C, Yu H J, Yu N,etal. Karyotype features of 10 species ofClematisand cluster analysis of karyotypes resemblance-near coefficient. Journal of Hunan Agricultural University: Natural Sciences, 2012, 38(6): 617-622.
彭绿春, 于恒隽, 余娜, 等. 10种铁线莲的核型特征及核型似近系数聚类分析. 湖南农业大学学报: 自然科学版, 2012, 38(6): 617-622.
[12] Sheng L. Karyotype Analysis on Sixteen Species of GenusClematis[D]. Nanjing: Nanjing Forestry University, 2011.
盛璐. 铁钱莲属16种植物的核型分析[D]. 南京: 南京林业大学, 2011.
[13] Sheng L. Karyotype Analysis and Molecular Systematics Study of GenusClematis[D]. Nanjing: Nanjing Forestry University, 2015.
盛璐. 铁线莲属植物的核型分析及分子系统学研究[D]. 南京: 南京林业大学, 2015.
[14] Sheidai M, Habibi M, Azizian D. Cytology and palynology of theClematisL. species (Ranunculaceae) in Iran. Acta Botanica Croatica, 2009, 68(1): 67-77.
[15] Duan Q, Cui G F, Wang X N,etal. Genetic diversity ofLiliumsargentiaebased on phenotypic traits and ISSR markers. Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(6): 1106-1113.
段青, 崔光芬, 王祥宁, 等. 云南泸定百合遗传多样性的表型与ISSR分析. 西北植物学报, 2013, 33(6): 1106-1113.
[16] Li Z Y, Hua Y W, Ge X H,etal. Chromosomal behaviors in plant wide hybridizations and their genetic and evolutionary implications. Hereditas(Beijing), 2005, (2): 315-324.
李再云, 华玉伟, 葛贤宏, 等. 植物远缘杂交中的染色体行为及其遗传与进化意义. 遗传, 2005, (2): 315-324.
[17] Li M X, Zhang Z P. Chromosome and Research Techniques of Crops[M]. Beijing: China Agriculture Press, 1996: 292-297.
李懋学, 张赞平. 作物染色体及其研究技术[M]. 北京: 中国农业出版社, 1996: 292-297.
[18] Li M X, Chen R Y. The standardization about the karyotype analysis. Journal of Wuhan Botanical Research, 1985, 3(4): 297-302.
李懋学, 陈瑞阳. 关于核型分析的标准化问题. 武汉植物学研究, 1985, 3(4): 297-302.
[19] Stebbins G L. Chromosomal Evolution in Higher Plants[M]. London: Edward Arnold Press, 1971: 87-89.
[20] Levan A, Fredga K, Sandberg A A. Nomenclature for centromeric position on chromosomes. Hereditas, 1964, 52(2): 201-220.
[21] Arano H. Cytological studies in subfamily Carduoideae (Compositae) of Japan. Botanical Magazine of Tokyo, 1963, 76: 32-39.
[22] Zhou Q X, Yang Y P, Zhang M L. Karyotypes of fourteen species inCaragana. Bulletin of Botanical Research, 2002, 22(4): 492-496, 518-519.
周其兴, 杨永平, 张明理. 锦鸡儿属植物14个种类的核型. 植物研究, 2002, 22(4): 492-496, 518-519.
[23] Shang B L, Meng K B, Wang Y J. Karyotype analysis of fiveSaussureaspecies. Acta Botanica Boreali-Occidentalia Sinica, 2014, 34(11): 2220-2226.
尚宝龙, 蒙奎宾, 王玉金. 风毛菊属5种植物的核型分析. 西北植物学报, 2014, 34(11): 2220-2226.
[24] Zhang F, Xing S Y, Han C J,etal. Analysis on karyotype and its evolutional trend ofGinkgobilobavar.epiphyllagermplasms resources. Acta Horticulturae Sinica, 2011, 38(12): 2245-2252.
张芳, 邢世岩, 韩晨静, 等. 叶籽银杏种质资源染色体核型分析及进化趋势. 园艺学报, 2011, 38(12): 2245-2252.
[25] Li Z, Chen S M, Chen F D,etal. Distribution of 45S rDNA sequence on chromosomes of five species inArtemisia. Acta Horticulturae Sinica, 2011, 38(2): 353-360.
李真, 陈素梅, 陈发棣, 等. 45S rDNA在蒿属5种植物染色体中的分布. 园艺学报, 2011, 38(2): 353-360.
KaryotypeanalysisofsevenwildClematisspecies
WANG Na, WANG Kui-Ling, LIU Qing-Hua, LIU Qing-Chao*
College of Landscape Architecture and Forestry, Qingdao Agricultural University, Qingdao 266109, China
There are many significant differences in the wild species ofClematis, including differences in morphology, aroma, flower number, green period and life form. These differences reflect the species’ complex genetic background. It is advantageous to study the chromosome karyotypes of the variousClematisspecies prior to any attempts at cross-breeding. In this study, traditional squashing methods were used to study the karyotypes of 7 wildClematisspecies:Clematisorientalis,C.patens,C.heracleifolia,C.fusca,C.hexapetala,C.serratifoliaandC.ternifloravar.mandshurica. The results showed that six of theClematisspecies were diploid (2n=2x=16), with the exception beingC.ternifloravar.mandshurica, which is tetraploid (2n=4x=32). The basic chromosome number was 8. The karyotypes of all the testedClematisspecies were mainly made of median, submedian, subterminal and terminal chromosomes. Their karyotype formulae are as followsC.patensandC.heracleifolia: 2n=2x=16=5m+2st+t;C.fusca: 2n=2x=16=5m+3st;C.hexapetala: 2n=2x=16=5m+2st(SAT)+t;C.serratifolia: 2n=2x=16=5m(SAT)+2st+t;C.orientalis: 2n=2x=16=5m(SAT)+sm(SAT)+2st;C.ternifloravar.mandshurica: 2n=4x=32=5m+2st+t. The karyotype formulae ofC.orientalis,C.hexapetalaandC.serratifoliahad satellites in the long arm of their chromosomes. The asymmetry index of the chromosomes ranged from 62.27% to 66.77%. The karyotypes of theseClematisspecies were 2A, showing that these seven species are relatively primitive. The karyotypes ofC.fuscaandC.serratifoliaare reported here for the first time.
Clematis; chromosome; karyotype analysis
10.11686/cyxb2017025http//cyxb.lzu.edu.cn
王娜, 王奎玲, 刘庆华, 刘庆超. 七种野生铁线莲属植物核型分析. 草业学报, 2017, 26(11): 123-130.
WANG Na, WANG Kui-Ling, LIU Qing-Hua, LIU Qing-Chao. Karyotype analysis of seven wildClematisspecies. Acta Prataculturae Sinica, 2017, 26(11): 123-130.
2017-01-18;改回日期:2017-03-29
青岛市公共领域科技支撑计划项目(12-1-3-49-nsh)资助。
王娜(1990-),女,山东德州人,在读硕士。E-mail: air_wn1990@126.com
*通信作者Corresponding author. E-mail: liuqingchao7205@126.com
毛茛科(Ranunculaceae)铁线莲属(Clematis)植物花色丰富、花期长,有较高的观赏价值[1]和较强的抗逆性[2-3],此外其药用价值也不可忽视[4-5]。铁线莲属植物广泛分布于世界各地,约有340多种,其中我国有140多种,而这些宝贵的资源是铁线莲种质创新与遗传改良的物质基础。
