喀斯特高原山地区水源涵养林土壤及凋落物的生态化学计量特征
2017-11-16秦仕忆喻阳华邢容容
秦仕忆,喻阳华,邢容容,王 璐
(1.贵州师范大学 地理与环境科学学院,贵阳 550025;2.贵州师范大学 喀斯特研究院,贵阳 550001;3.贵州大学 资源与环境工程学院,贵阳 550025)
喀斯特高原山地区水源涵养林土壤及凋落物的生态化学计量特征
秦仕忆1,喻阳华2,邢容容3,王 璐1
(1.贵州师范大学 地理与环境科学学院,贵阳 550025;2.贵州师范大学 喀斯特研究院,贵阳 550001;3.贵州大学 资源与环境工程学院,贵阳 550025)
探明石漠化区水源涵养林凋落物与土壤化学计量特征有助于深入了解养分状况。以喀斯特高原山地区云南松(Pinusyunnanensis)+光皮桦(Betulaluminifera)+银白杨(Populusalba)林、栓皮栎(Quercusvariabilis)+川榛(Corylusheterophylla)林、银白杨(Populusalba)+光皮桦(Betulaluminifera)林、杜鹃(Rhododendronsimsii)+银白杨(Populusalba)林、光皮桦(Betulaluminifera)林为对象,研究凋落物与土壤OC,TN,TP化学计量特征及内在关联。结果表明:1)表层土壤OC,TN,TP含量分别是25.02,1.56,0.33mg/g;凋落物OC,TN,TP含量分别为407.09,16.07,2.06mg/g。2)土壤C∶N为15.13,C∶P为78.40,N∶P为5.17;凋落物C∶N为25.67,C∶P为212.87,N∶P为8.38;C∶N,C∶P,N∶P均呈凋落物层>土壤层。3)凋落物层、土壤层C∶P与N∶P均呈极显著正相关,土壤C∶N与凋落物C∶N,N∶P分别呈极显著正、显著负相关。4)该区土壤N,P养分亏缺,凋落物呈低C高N高P格局,土壤固持养分能力不强。
水源涵养林;凋落物;土壤;生态化学计量;喀斯特高原山地区
0 引言
生态化学计量学是一门分析多重化学元素质量平衡与生态过程间相互影响的科学,为研究退化生态系统植物内稳性与N,P等限制提供了有效手段[1]。C,N,P作为植物生长的必需元素,对植物的生长发育、养分循环以及个体功能的运行具有重要作用[2-3]。凋落物分解过程中养分的归还使得土壤养分供应量具有明显的时空变异[4-5],增加了二者间关系的复杂性。而在喀斯特强烈发育的地区,凋落物养分的归还量是贫瘠土壤养分的重要来源[6],使得对森林群落凋落物、土壤的生态化学计量特征研究具有十分重要的理论和现实意义。对喀斯特区土壤水分和养分水平研究表明:土层厚度、土壤有机质含量、土壤颗粒组成及0.03~0.3mm孔隙度是影响土壤水分库容的主要因素[7];推行自然恢复,减少人为干扰可有效提高喀斯特退化生态系统土壤生产力,增加土壤有机碳积累[8]。在生态化学计量学研究方面:潘复静等[9]分析了喀斯特峰丛洼地4个不同演替阶段植被生态化学计量特征在不同坡位间的差异,发现在上坡位,成熟林群落的凋落物有利于积累养分;曾昭霞等[10]对喀斯特原生林和次生林植物-凋落物-土壤的研究结果表明,喀斯特森林凋落物与植物均呈现低C∶N和高N∶P的生态化学计量特征;张亚冰等[11]应用生态化学计量学的方法对贵州月亮山5种森林类型土壤进行研究,发现不同森林类型的C,N,P含量及其化学计量特征存在显著差异。以上学者的研究,初步揭示了喀斯特森林凋落物、土壤养分含量及生态化学计量。水源涵养林在拦蓄降水、调节径流和净化水质等方面的重要功能对喀斯特高原山地区十分重要[12],但是关于此地区水源涵养林生态化学计量特征方面的研究却鲜有报到。本文以喀斯特高原山地区5种典型水源涵养林为研究对象,对不同植物群落凋落物、土壤的C,N,P含量及生态学计量特征予以研究,试图回答不同水源涵养林的养分变化及其相互关系,以期完善喀斯特退化生态系统生态化学计量学的研究。
1 材料与方法
1.1研究区概况
毕节撒拉溪喀斯特高原山地潜在-轻度石漠化综合治理示范区位于贵州省毕节市西南部(27°11′36″~27°16′51″N,105°02′01″~105°08′09″E),喀斯特面积占74.25%,海拔1 600~1 950m,属长江流域乌江水系六冲河上游支流区。土壤以黄壤为主,有部分黄棕壤和石灰土;植被以亚热带针、阔叶林和落叶阔叶林为主;年均气温12.9℃,≥10℃积温4 109℃,年均日照1 261h,无霜期258d,年相对湿度81.69%,年均降雨量984.40mm,降水主要集中在5—9月,占年降雨总量的57.6%~87.3%[13]。
1.2样品采集
水源涵养林以云南松(Pinusyunnanensis)、光皮桦(Betulaluminifera)、银白杨(Populusalba)、栓皮栎(Quercusvariabilis)、川榛(Corylusheterophylla)、杜鹃(Rhododendronsimsii)为优势树种。本文选取云南松+光皮桦+银白杨林(1号)、栓皮栎+川榛林(2号)、银白杨+光皮桦林(3号)、杜鹃+银白杨林(4号)、光皮桦林(5号)等5种水源涵养林为研究对象(表1)。于2016年7月中旬,在不同水源涵养林内分别设置20m×20m样地,采用蛇形布点法设置5~7个取样点,用尼龙网袋收集凋落物组成混合样品,同时采集0~20cm表层土壤制成混合样品。样品带回实验室后,凋落物置于恒温干燥箱中,65℃烘至恒质量,研细并充分混匀备用;土壤剔除可见石砾、根系及动植物残体,自然风干后研磨,依次过2.00,1.00,0.250,0.150mm筛备用。
1.3样品分析
用重铬酸钾-外加热法测定凋落物与土壤的有机碳。用凯氏定氮法测定土壤全氮;用高氯酸-硫酸消煮,钼锑抗比色法测定土壤全磷。用硫酸-过氧化氢消煮,奈氏比色法测定凋落物全氮;用硫酸-过氧化氢消煮,钼锑抗比色法测定凋落物全磷[14]。
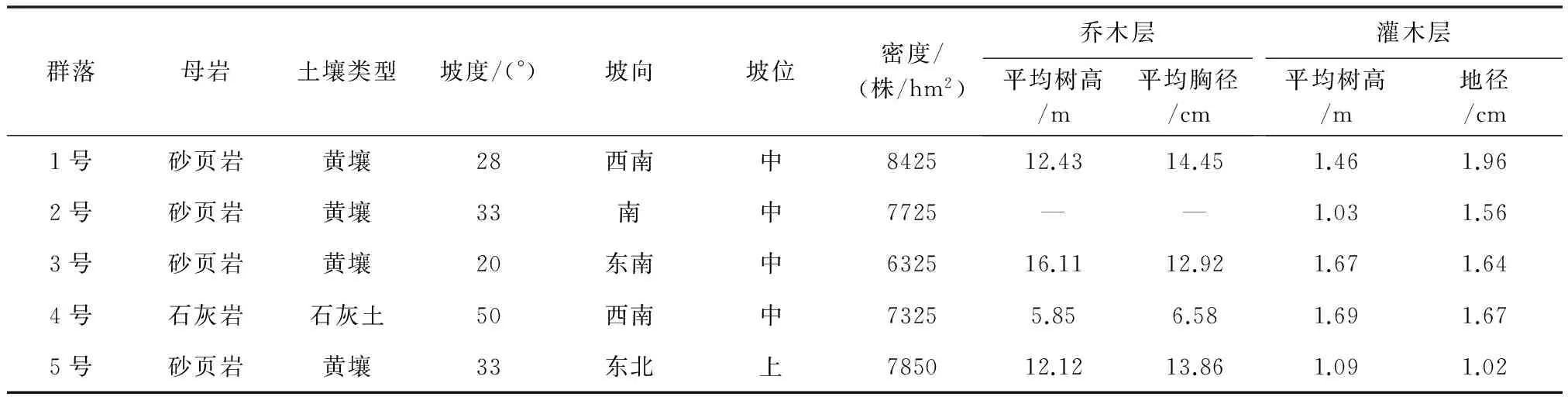
表1 植物群落基本概况
注:“—”表示没有出现。
1.4数据处理与分析
所有实验数据在Excel 2010和SPSS 20统计软件中进行统计、整理。采用单因素方差分析(one-way ANOVA)的最小显著差异(LSD)法对凋落物和土壤有机碳(OC)、全氮(TN)、全磷(TP)及其化学计量比进行差异显著性检验,采用Pearson相关系数法进行指标之间的相关性分析。
2 结果与分析
2.1不同群落土壤与凋落物OC,TN,TP含量
从表2可知,各水源涵养林OC,TN,TP含量变化规律较为相似。土壤与凋落物的OC,TN,TP含量均为OC>TN>TP,表明土壤与凋落物连续体间具有明显的关联性,且凋落物OC,TN,TP高于土壤,这与植物体为维持自身正常代谢,吸收土壤中的大部分营养,并以凋落物的形式逐步返还给土壤的实际相符合[6]。不同群落土壤:OC与TN均为4号最大(55.74,3.20mg/g)、3号最小(9.24,0.70mg/g),各群落间差异显著(P<0.05);TP为2号(0.63mg/g)显著高于其他群落,是其他群落的1~4倍,1号最低仅为0.16mg/g。不同群落凋落物:1)OC为5号最高(430.45mg/g)、2号最低(389.61mg/g),TN为1号最高(18.02mg/g)、2号最低(13.49mg/g),表明5号凋落物分解速率相对较慢,2号凋落物分解速率相对较快;2)TP差异不显著(P>0.05)。
2.2不同群落土壤与凋落物C-N-P生态化学计量特征
不同群落土壤的生态化学计量比如表3所示:C∶N为4号最大(17.44),与2号、5号差异不显著(P>0.05),与1号、3号呈显著差异(P<0.05),1号群落最小(12.09);C∶P,N∶P均为4号最高(分别为129.45,7.42)、2号最低(分别为32.44,2.03),4号在这两个指标上均约为2号的4倍,这与4号具有较高的OC与TN有关,不同群落间均呈显著差异(P<0.05)。如表3所示,不同群落凋落物C∶N表现为2号最大(28.91)、1号最小(22.22),但群落间差异不显著(P>0.05);C∶P差异较小,5个群落之间无显著差异(P>0.05);N∶P表现为1号最高,为12.01,与3,4,5号差异不显著(P>0.05),与2号差异显著(P<0.05)。
5种群落中:土壤的C∶N波动范围为12.09~17.44(均值15.13),C∶P为32.44~129.45(均值78.40),N∶P为2.03~7.42(均值5.17);凋落物的C∶N在22.22~28.91之间(均值25.67),C∶P为183.19~268.98(均值212.87),N∶P变幅为6.82~12.01(均值8.38)。凋落物的C∶N是土壤C∶N的1.70倍,C∶P是土壤的2.72倍,N∶P是土壤的1.62倍,表明群落C∶N,C∶P,N∶P均为凋落物层>土壤层。
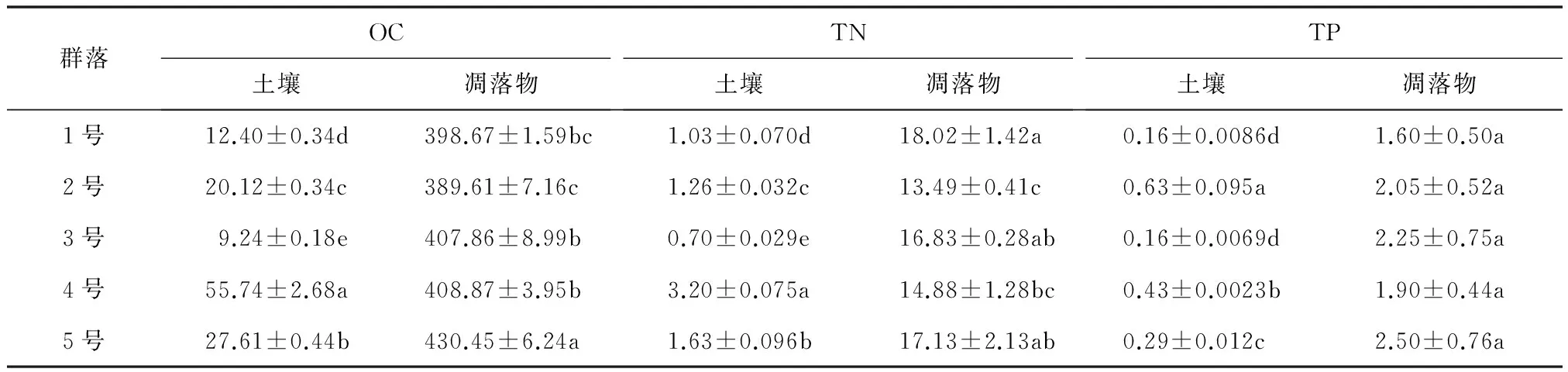
表2 群落凋落物与土壤的OC,TN,TP含量 mg/g
注:表中数据为平均值±标准差,同列不同小写字母表示差异显著(P<0.05)。
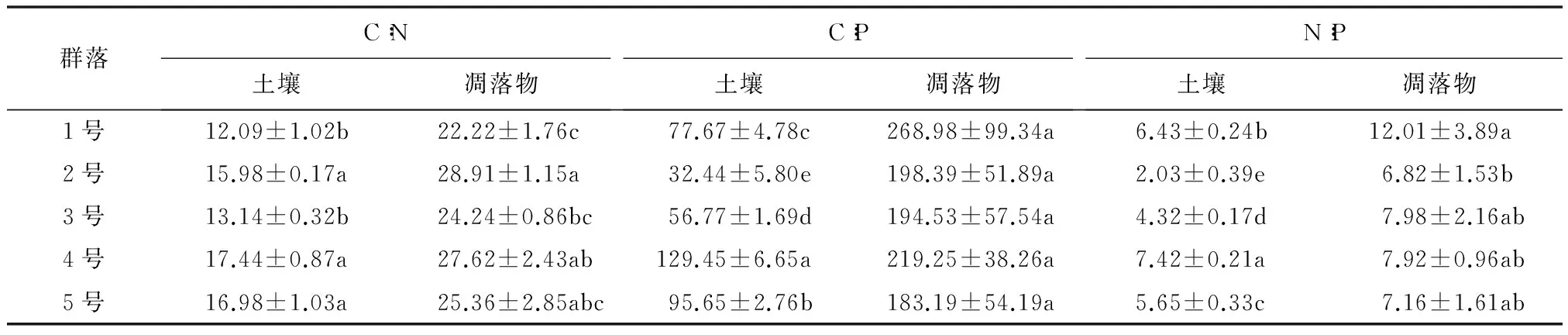
表3 群落凋落物与土壤的生态化学计量特征
注:表中数据为平均值±标准差,同列不同小写字母表示差异显著(P<0.05)。
2.3土壤与凋落物OC,TN,TP含量及其化学计量比的相关性
由表4可知:1)土壤的OC与TN,C∶N,C∶P呈极显著正相关(P<0.01),相关系数分别达到了0.99,0.77,0.79;与N∶P呈显著正相关(P<0.05),相关系数为0.53。土壤的TN与C∶N,C∶P呈极显著正相关(P<0.01,相关系数分别为0.71,0.81);与N∶P呈显著正相关(P<0.05,相关系数为0.58)。土壤的TP与C∶N呈显著正相关(P<0.05),相关系数为0.63。土壤的C∶P与N∶P呈极显著正相关(P<0.01),相关系数是0.91。2)凋落物的TN与C∶N呈极显著负相关(P<0.01),相关系数为-0.96;凋落物的TP与C∶P,N∶P呈极显著负相关(P<0.01),相关系数分别为-0.91,-0.79;凋落物的C∶P与N∶P呈极显著正相关(P<0.01),相关系数为0.93。3)土壤的TP与凋落物的TN呈极显著负相关(P<0.01),系数为-0.78;与凋落物C∶N呈极显著正相关(P<0.01),系数是0.78。土壤的C∶N与凋落物的C∶N呈极显著正相关(P<0.01),相关系数达到0.65;与凋落物N∶P呈显著负相关(P<0.05),相关系数是-0.55。土壤的C∶P与凋落物的OC的相关系数达到了0.52,为显著正相关(P<0.05)。
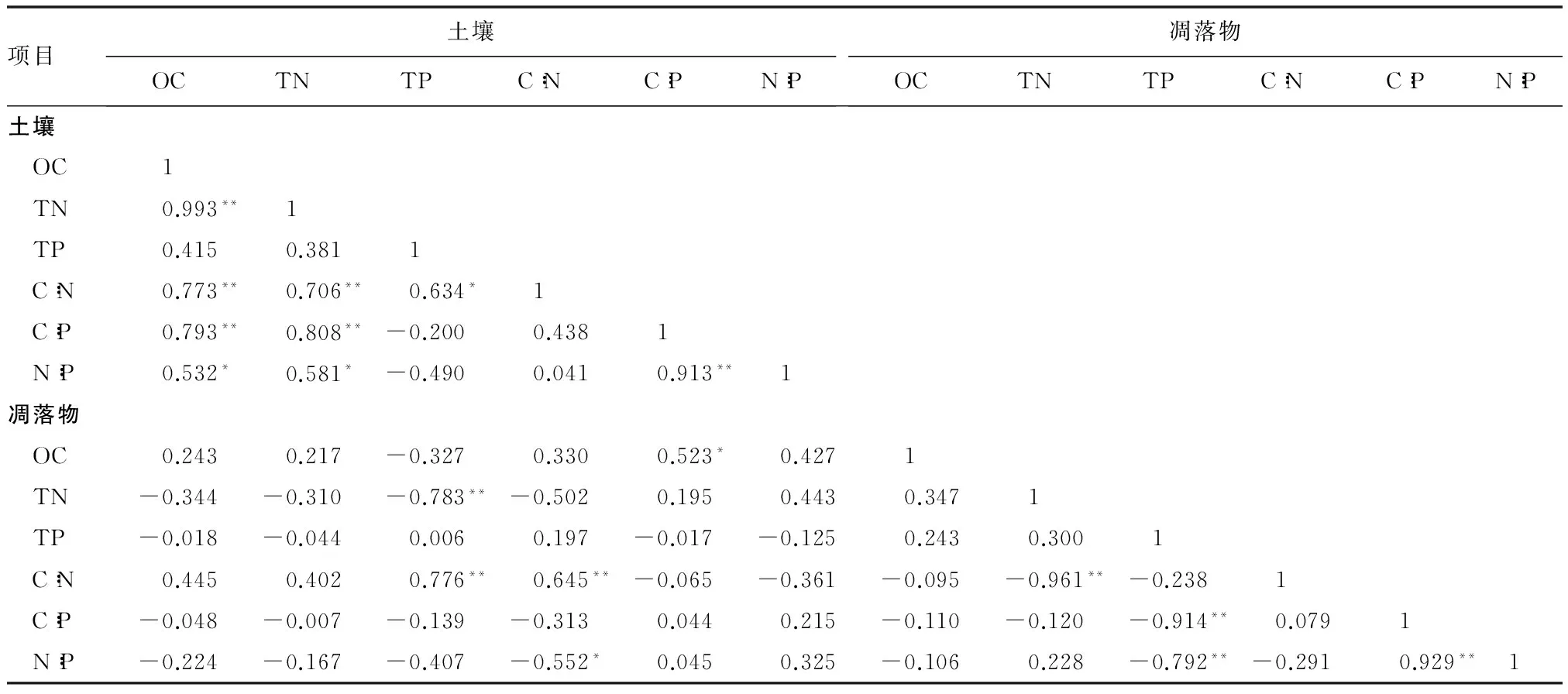
表4 凋落物与土壤OC,TN,TP含量及生态化学计量比的相关性
注:“*”表示在0.05水平(双侧)上显著相关;“**”表示在0.01水平(双侧)上显著相关。
3 讨论
3.1群落土壤的生态化学计量特征
从表5可以看出,研究区0~20cm土壤层OC含量为9.24~55.74mg/g,平均值为25.02mg/g。土壤C,N等养分主要来源于凋落物的分解,这致使养分首先在土壤表层聚集。在随水或其他介质向下层逐渐扩散的过程中,养分浓度存在明显的下降趋势[15]。研究区0~20cm土壤层OC均值高于赣中地区4种林分0~10cm土壤层OC含量[16],与全国土壤0~10cm土壤层OC含量[17]接近,表明示范区0~20cm土壤层OC含量处于较高水平。而土壤OC的形成和分解受植被类型、温度、降水、成土母质、土壤性质等因子的影响[18],该区土壤OC含量水平与喀斯特高原山地区高寒干旱的气候环境有一定关系。同时,研究区土壤OC含量低于广西环江木论喀斯特保护区2种原生林分0~20cm土壤层OC平均含量[19],这可能与喀斯特峰丛洼地复杂的生境,较高的空间异质性有关[20];研究区0~20cm土壤层TN变化范围是0.70~3.20mg/g,平均值为1.56mg/g,低于广西环江木论喀斯特保护区2种原生林分0~20cm土壤层TN含量[19];研究区0~20cm土壤层TP的变化幅度为0.16~0.63mg/g,均值是0.33mg/g。土壤磷(P)素的来源较为固定,主要是通过漫长的岩石风化,所以在土壤中的垂直分布中呈“圆柱体”状,差异不大[15],使得不同深度的研究区之间存在可比性。研究区TP含量虽然高于广西喀斯特地区0~20cm土壤TP含量[19]和赣中地区4种林分0~10cm土壤TP的研究成果[16],但仅达全国0~10cm土壤层TP平均水平[17]的42.86%,说明研究区0~20cm土壤层的TP含量较低。这可能与研究区地表流失和地下漏失严重有关。
贵州喀斯特高原山地区水源涵养林0~20cm土壤层C∶P,N∶P均值分别是78.40和5.17,仅为广西环江喀斯特地区[19]0~20cm土壤层C∶P,N∶P值的37.48%和21.72%,同时也低于赣中地区[16]、全国0~10cm土壤层平均水平[17]和全球0~10cm森林土壤层[21]。可见,研究区土壤C∶P,N∶P值均较低。C∶P是指示P有效性高低的一个指标[22],较低的C∶P说明研究区土壤P具有较高的有效性。同时,有研究提出,在土壤N,P含量不太高或太低的前提下,可将N∶P<10和N∶P>20作为评价植被生产力受到N或者P限制的指标[23]。研究区土壤N∶P均值为5.17<10,表明该区域的林分生产力主要是受N的限制。综合分析表明,森林土壤N,P处于亏缺状态,因此在群落建植过程中应注重养分质量的保护,可采取促进养分循环和自肥措施。尤其是促进主要限制养分N的循环,是研究区水源涵养林恢复的重要途径之一。
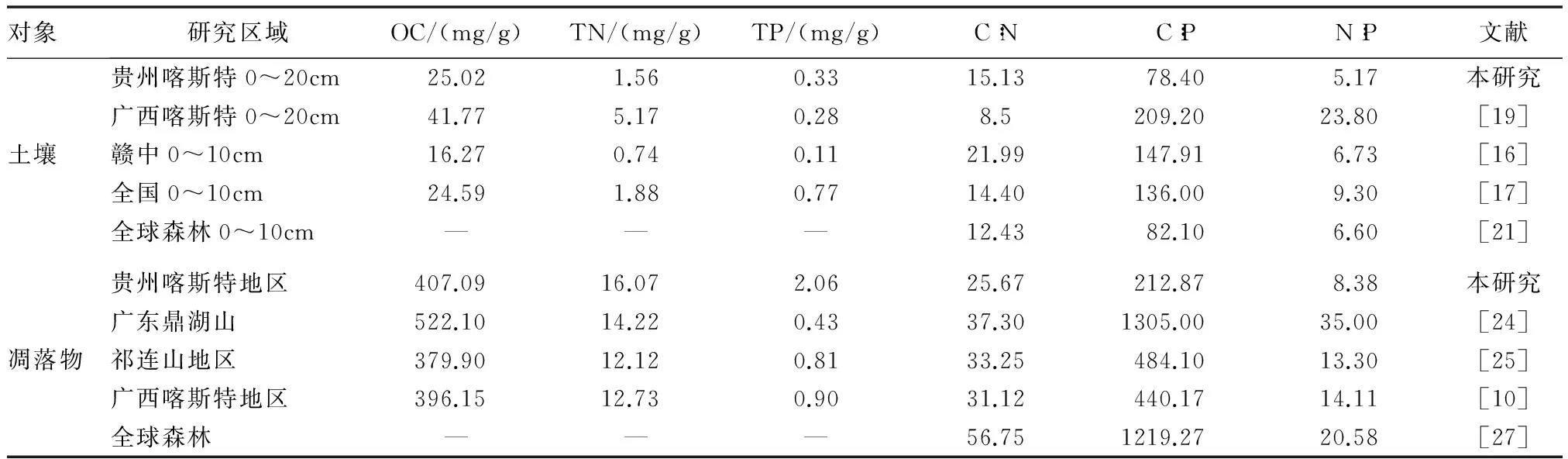
表5 贵州喀斯特高寒山地与其他区域凋落物 土壤的C,N,P含量及计量特征
注:表中所示凋落物、土壤元素含量直接来自文献或是各文献所研究对象中元素含量的平均值。
3.2群落凋落物的生态化学计量特征
由表5可知,5种群落凋落物OC含量389.61~430.45mg/g(均值为407.09mg/g),TN为13.49~18.02mg/g(平均值是16.07mg/g),TP为1.60~2.50mg/g(平均值为2.06mg/g),相对于广东鼎湖山亚热带常绿阔叶林[24],其OC含量低于后者,而TN,TP含量高于后者。研究区TN,TP含量也高于祁连山高寒半干旱山地青海云杉[25]、广西环江喀斯特地区森林[10]和全球水平[26](10.90,0.90),表明研究区凋落物OC含量处于偏低水平,TN,TP含量较高。研究区喀斯特森林凋落物呈现出低C高N高P格局,这可能与凋落物的分解程度、植物体对养分的重吸收程度以及研究区阔叶落叶树种较多有关。
凋落物C∶N,C∶P和N∶P的均值依次为25.67,212.87,8.38,均低于广东鼎湖山亚热带常绿阔叶林[24]、祁连山青海云杉林[25]、广西环江喀斯特森林[10]以及全球森林[27]的研究结果。表明研究区C∶N,C∶P和N∶P均较低,原因与凋落物呈低C高N高P格局有关。有研究认为,凋落物较低的C∶N具有较高的分解速率[22],这符合落叶阔叶树种凋落物的分解规律,但凋落物较快的分解速率不利于养分的储存[28]。研究区凋落物TN,TP较高,但土壤TN,TP较低,说明土壤蓄养水分能力较弱,可能与植被退化有一定关联。因此,采取封山育林、基于生物多样性的石漠化综合治理等措施,对于保持研究区土壤养分质量尤为必要。
3.3群落土壤与凋落物生态化学计量学的相关性
C作为构成植物体干物质的最主要元素[29],N,P作为生物体蛋白质和遗传物质的基本组成元素,均对植物的各种功能影响深刻[30]。土壤C,N,P是植物生长的重要元素,土壤养分供养状况与植物光合作用、矿质代谢等生态过程密切相关[31]。而凋落物是养分回归土壤的主要途径,是森林系统生物地球化学流的一个重要组成部分,已有研究表明森林生长所需70%~90%的养分来自凋落物的降解[32]。本研究中,土壤和凋落物的C∶P与N∶P均呈极显著正相关关系(系数分别为0.91,0.93),土壤C∶N与凋落物C∶N之间存在极显著正相关关系(相关系数是0.65),土壤C∶N与凋落物N∶P呈显著负相关(相关系数为-0.55),这与赵维俊等[25]对祁连山青海云杉林的研究结论一致,说明土壤C,N主要来源于凋落物,凋落物保留了大量的P养分,但是归还至土壤中的P较少,这与土壤P主要来源于土壤母质的事实相符合[15]。
研究区水源涵养林0~20cm土壤层的C∶N∶P质量比均值为78.22∶5.17∶1,与全球0~10cm森林土壤层C∶N∶P质量比(82.04∶6.60∶1)[21]接近;凋落物C∶N∶P质量比均值是215.11∶8.38∶1,低于全球森林凋落物水平(1 167.92∶20.58∶1)[27]。这与潘复静[9]等对另一喀斯特地区植被群落凋落物的研究结论一致。同时,5种水源涵养林凋落物C∶N∶P质量比分别是266.86∶12.01∶1,197.17∶6.82∶1,193.44∶7.98∶1,218.75∶7.92∶1和181.58∶7.16∶1,群落间较为接近。由于植物在长期的进化过程中逐渐发育了较强的生理生化调节作用以适应环境因子的波动[33],植物体C,N,P含量均会随环境不断变化[10];而相似的环境因子可能是群落间C∶N∶P比值在一定程度上趋于规律性的原因。
4 结论
本研究仅对不同水源涵养林的土壤、凋落物养分变化和化学计量特征进行了初步分析,对于全面评价森林生态系统的养分状况,还需要结合叶片、根系等构件中养分含量及其生态化学计量关系开展进一步的研究,即将不同尺度、不同生活型植物、不同植物群落的养分循环有机联系起来,以揭示养分在植被-凋落物-土壤之间的生态化学计量特征及其相互关系,指示养分限制状况。
本研究中,5种水源涵养林土壤N,P均处于亏缺状态,凋落物呈现低C高N高P格局。土壤固持养分能力不强,应对水源涵养林采取保育和恢复措施。
[1] 贾国梅,何立,程虎,等.三峡库区不同植被土壤微生物量碳氮磷生态化学计量特征[J].水土保持研究,2016,23(4):23-27.
[2] Vitousek P M,Porder S,Houlton B Z,et al.Terrestrial phosphorus limitation:mechanisms,implications,and nitrogen-phosphorus interactions[J].Ecological Applications,2010,20(1):5-15.
[3] 王维奇,徐玲琳,曾从盛,等.河口湿地植物活体枯落物土壤的碳氮磷生态化学计量特征[J].生态学报,2011,31(23):7119-7124.
[4] Han Wenxuan,Fang Jingyun,Reich P B,et al.Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate,soil and plant functional type in China[J].Ecology Letters,2011,14(8):788-796.
[5] Frouz J,Kalcík J,Velichová V.Factors causing spatial heterogeneity in soil properties,plant cover,and soil fauna in non-reclaimed post-mining site[J].Ecological Engineering,2011,37(11):1910-1913.
[6] 左巍,贺康宁,田赟,等.青海高寒区不同林分类型凋落物养分状况及化学计量特征[J].生态学杂志,2016,35(9):2271-2278.
[7] 李孝良,陈效民,周炼川,等.西南喀斯特石漠化过程对土壤水分特性的影响[J].水土保持学报,2008,22(5):198-203.
[8] 魏亚伟,苏以荣,陈香碧,等.人为干扰对桂西北喀斯特生态系统土壤有机碳、氮、磷和微生物量剖面分布的影响[J].水土保持学报,2010,24(3):164-169.
[9] 潘复静,张伟,王克林,等.典型喀斯特峰丛洼地植被群落凋落物C∶N∶P生态化学计量特征[J].生态学报,2011,31(2):0335-0343.
[10] 曾昭霞,王克林,刘孝利,等.桂西北喀斯特森林植物-凋落物-土壤生态化学计量特征[J].植物生态学报,2015,39(7):682-693.
[11] 张亚冰,吕文强,易武英,等.贵州月亮山5种森林类型土壤生态化学计量特征研究[J].热带亚热带植物学报,2016,24(6):617-625.
[12] 赵洋毅,王玉杰,王云琦,等.渝北水源区水源涵养林构建模式对土壤渗透性的影响[J].生态学报,2010,30(15):4162-4172.
[13] 池永宽.石漠化治理中农草林草空间优化配置技术与示范[D].贵阳:贵州师范大学,2015.
[14] 鲍士旦.土壤农化分析[M].第3版.北京:中国农业出版社,2008:74-271.
[15] 刘兴诏,周国逸,张德强,等.南亚热带森林不同演替阶段植物与土壤的N、P的化学计量特征[J].植物生态学报,2010,34(1):64-71.
[16] 杜满义,范少辉,刘广路,等.土地利用方式转变对赣中地区土壤活性有机碳的影响[J].应用生态学报,2013,24(10):2897-2904.
[17] Tian Hanqin,Chen Guangsheng,Zhang Chi,et al.Pattern and variation of C∶N∶P ratios in China’s soils:a synthesis of observational data[J].Biogeochemistry,2010,98:139-151.
[18] 白小芳,徐福利,王渭玲,等.华北落叶松人工林土壤碳氮磷生态化学计量特征[J].中国水土保持科学,2015,13(6):68-75.
[19] 俞月凤,彭晚霞,宋同清,等.喀斯特峰丛洼地不同森林类型植物和土壤C、N、P化学计量特征[J].应用生态学报,2014,25(4):947-954.
[20] 宋同清,彭晚霞,曾馥平,等.喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征[J].生物多样性,2010,18(4):355-364.
[21] Cleveland C C,Liptzin D.C∶N∶P stoichiometry in soil:is there a “Redfield ratio” for the microbial biomass?[J].Biogeochemistry,2007,85:235-252.
[22] 王绍强,于贵瑞.生态系统碳氮磷元素的生态化学计量学特征[J].生态学报,2008,28(8):2938-2947.
[23] Güsewell S.N∶P ratios in terrestrial plants:variation and functional significance[J].New Phytologist,2004,164:243-266.
[24] 王晶苑,王绍强,李纫兰,等.中国四种森林类型主要优势植物的C∶N∶P化学计量学特征[J].植物生态学报,2011,35(6):587-595.
[25] 赵维俊,刘贤德,金铭,等.祁连山青海云杉林叶片-枯落物-土壤的碳氮磷生态化学计量特征[J].土壤学报,2016,53(2):477-489.
[26] Kang Hongzhang,Xin Zaijun,Breg B,et al.Global pattern of leaf litter nitrogen and phosphorus in woody plants[J].Annals of Forest Science,2010,67,811.
[27] McGroddy M,Daufresne T,Hedin L O.Scaling of C∶N∶P stoichiometry in forests worldwide:implications of terrestrial redfield-type ratios[J].Ecology,2004,85(9):2390-2401.
[28] 杨佳佳,张向茹,马露莎,等.黄土高原刺槐林不同组分生态化学计量关系研究[J].土壤学报,2014,51(1):133-142.
[29] 项文化,黄志宏,闫文德,等.森林生态系统碳氮循环功能耦合研究综述[J].2006,26(7):2365-2372.
[30] 平川,王传宽,全先奎.环境变化对兴安落叶松氮磷化学计量特征的影响[J].生态学报,2014,34(8):1965-1974.
[31] 宾振钧,王静静,张文鹏,等.氮肥添加对青藏高原高寒草甸6个群落优势化学计量学特征的影响[J].植物生态学报,2014,38(3):213-237.
[32] 赵其国,王明珠,何园球.我国热带亚热带森林凋落物及其对土壤的影响[J].土壤,1991(1):8-15.
[33] 曾德慧,陈广生.生态化学计量学:复杂生命系统奥秘的探索[J].植物生态学报,2005,29(6):1007-1019.
StoichiometricCharacteristicsofLitterandSoilsinWaterConservationForestsofKarstPlateauMountainousRegion
QIN Shiyi1,YU Yanghua2,XING Rongrong3,WANG Lu1
(1.SchoolofGeographyandEnvironmentalScience,GuizhouNormalUniversity,Guiyang550025,China;2.SchoolofKarstScience,GuizhouNormalUniversity,Guiyang550001,China;3.SchoolofResourcesandEnvironmentalEngineering,GuizhouUniversity,Guiyang550025,China)
It is helpful to understand the nutrient status by ascertaining stoichiometric characteristics of the litter and soil of water conservation forests in the rocky desertification area.In this paper,the forests ofPinusyunnanensis,BetulaluminiferaandPopulusalba,QuercusvariabilisandCorylusheterophylla,PopulusalbaandBetulaluminifera,RhododendronsimsiiandPopulusalbaandBetulaluminiferain the karst plateau mountainous were used to study the organic C,TN,TP stoichiometry and intrinsic association of litter and soil.The results show that soil organic C,TN and TP in Karst communities occurred primarily in the top 0~20 cm soil layer,at 25.02 mg/g C,1.56 mg/g N and 0.33 mg/g P,with ratios of 15.13 for C∶N,78.40 for C∶P and 5.17 for N∶P.The litter organic C,TN,TP content were estimated at 407.09 mg/g,16.07 mg/g,2.06 mg/g,and the corresponding C∶N,C∶P and N∶P were 25.67,212.87 and 8.38,respectively.In general,litter layer had higher C∶N,C∶P and N∶P than soil layer.There is a significant positive correlation between C∶P and N∶P in litter layer or soil layer,the correlation between soil C∶N and litter C∶N was negative.Besides,there was a significant negative correlation between soil C∶N and litter N∶P.Nutrition of N and P in soil is deficient,the litter has a clear structure of low C,high N and high P.The capacity of holding nutrient for soil is not strong.
water conservation forest,litter,soil,ecological stoichiometry,karst plateau mountain region
S153.1
A
1002-6622(2017)05-0066-08
2017-05-31;
2017-08-15
贵州省科技计划项目“石漠化区抗冻耐旱型植被建植与恢复关键技术研究”(黔科合LH字〔2016〕7201号)
秦仕忆(1994-),女,重庆江津人,主要从事退化生态系统修复研究。Email:15885501304@163.com
喻阳华(1984-),男,贵州仁怀人,副教授,博士,主要从事植被适应性修复研究。
Email:yuyanghua2003@163.com
10.13466/j.cnki.lyzygl.2017.05.012
