10种淫羊藿属植物核型分析及其分类学的意义
2017-11-10王霖娇田静盛茂银
王霖娇 ,田静 ,盛茂银 ,3*
(1.贵州师范大学喀斯特研究院,贵阳 550001;2.国家喀斯特石漠化防治工程技术研究中心,贵阳 550001;3.贵州省喀斯特石漠化防治与衍生产业工程实验室,贵阳 550001)
10种淫羊藿属植物核型分析及其分类学的意义
王霖娇1,2,田静1,2,盛茂银1,2,3*
(1.贵州师范大学喀斯特研究院,贵阳 550001;2.国家喀斯特石漠化防治工程技术研究中心,贵阳 550001;3.贵州省喀斯特石漠化防治与衍生产业工程实验室,贵阳 550001)
【目的】开展10种淫羊藿属植物核型分析,为该属植物系统分类和起源进化等领域的深入研究提供细胞学证据。【方法】运用压片法对10个淫羊藿属(Epimedium)植物的根尖有丝分裂进行观察,并对其核型进行比较研究。【结果】供试的10种淫羊藿属植物根尖细胞染色体数目均为12,都为二倍体,2n=2x=12。10个种的核型基本相似,都为2A对称核型。第1对同源染色体都具有中间随体;除E.perralderianum核型公式为2n=2x=8m(2SAT)+4sm外,其他9个种核型公式均为2n=2x=6m(2SAT)+6sm。【结论】核型分析结果支持该属属下Epimedium和Rhizophyllum 2个亚属的划分,也显示淫羊藿属植物在小檗科明显是一个起源上更古老的类群,且与其他属植物的亲缘关系较远。核型分析对淫羊藿属属下类群系统学研究意义有限,但对属上分类级别的系统进化研究具有重要的参考价值。
淫羊藿;核型分析;系统学;起源;演化
淫羊藿是我国重要的传统药用植物,又名仙灵脾、羊合叶,多年生草本植物,在分类上属于小檗科(Berberidaceae)淫羊藿属(Epimedium L.)[1]。早在《神农本草经》即记载其“主阴痿绝伤、径中痛、益气力、强志”等药用功效[2]。近年来,药理研究显示,淫羊藿不仅在免疫、生殖、核酸代谢、心脑血管、抗衰老等方面具有显著的药效[3],同时在肿瘤治疗方面也具有明显的应用前景[4]。此外,淫羊藿属植物具有奇异的花朵、多变的花色、常绿多年生的植株等优良园艺特性,在欧美、日本等地是十分受欢迎的园艺植物。近年来,淫羊藿属植物的研究已成为相关领域研究的热点。
该属目前全球已报道约60种[5],间断分布于日本至北非的阿尔及利亚之间的广大区域,可以分成3个分布区:①地中海和西亚地区,分布有4个种;②日本地区,分布约有5个种;③中国地区,是该属目前的分布中心,报道有约50种、5变种[6-7]。该属种类繁多,且种间差异不明显,在分类上一直是一个难以处理的类群[8]。迄今为止,Stearn[5]对该属的分类最为全面,依据该属地理分布、花茎上叶的数目和花的形态(包括花的大小、花瓣形态、花瓣与内轮萼片的相对大小),将该属划分Epimedium和Rhizophyllum两个亚属,Epimedium亚属划分为4个组:Diphyllon(中国类群)、Macroceras、Polyphyllon 和 Epimedium,Diphyllon组下又划分为 4 个系:Campanulatae、Davidianae、Dolichocerae和Brachycerae[5]。但该属系统分类研究仍存在一些明显问题,类群间的关系以及类群界限的划分仍不清楚,其起源与进化仍没有定论[9-11]。
细胞学的核型分析已广泛应用于植物分类学研究,可以为植物系统起源进化和种间亲缘关系研究提供重要的细胞学资料[12]。尽管淫羊藿属植物核型分析已开展了大量的工作[1,12-17],但由于该属物种繁多、分布广泛、材料采集困难,导致该属细胞学核型研究一直不系统、不深入,限制了核型分析在该属植物分类学研究中的应用。本文在长期收集的淫羊藿属植物资源的基础上,开展了来源于中国、日本、德国、美国等全世界不同地区10种该属植物的核型分析,以期为该属植物的系统分类和起源进化等领域的深入研究提供细胞学证据。
1 材料和方法
1.1 供试材料及来源
实验材料1~4号由作者及其团队实地野外采集,实验材料5~10号由作者与该地区相关研究机构学术交流收集获得。凭证标本存于贵州师范大学植物标本室,活体植株种植于贵州师范大学贵州省喀斯特石漠化防治与衍生产业工程实验室植物栽培室,详情见表1。
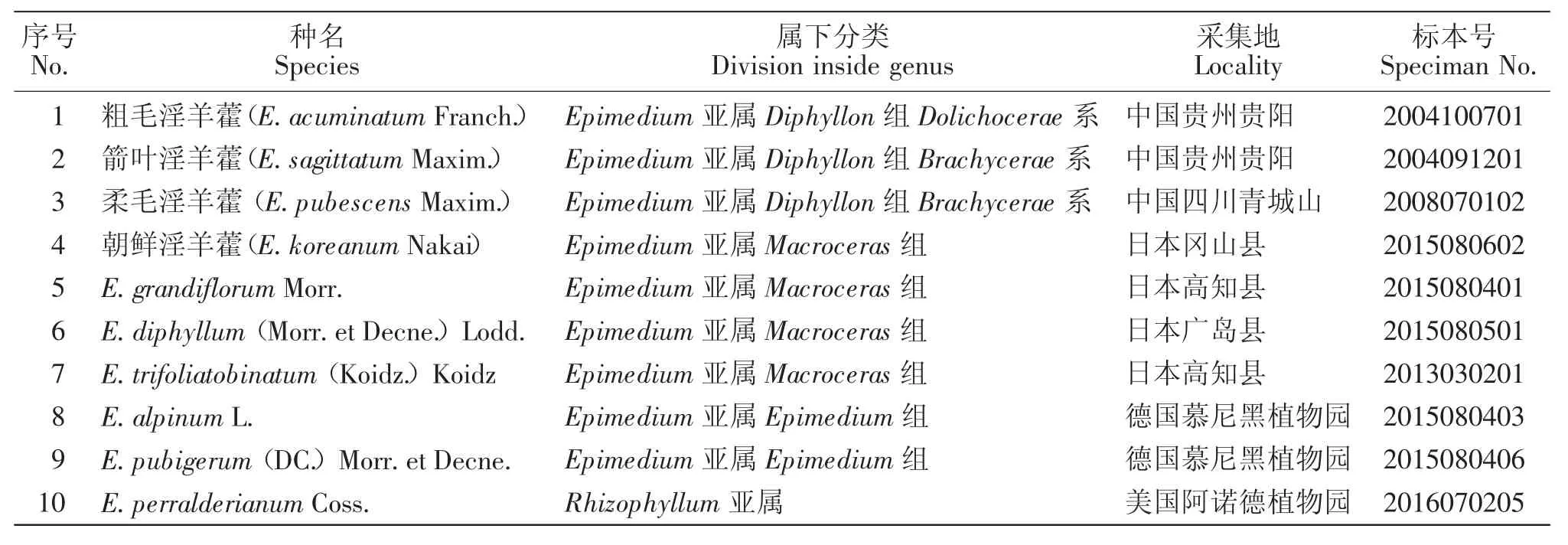
表1 供试材料及来源Table 1 Materials used in this study
1.2 实验方法
春季上午9:00—10:00从生长旺盛活体植株上剪取长1~2cm新生长的根尖,室温下置于饱和α-溴萘溶液中预处理5~6 h,卡诺I氏固定液(乙醇∶冰醋酸=3∶1)中固定 12 h 以上,1 mol/L HCl溶液 60 ℃恒温解离10min,改良苯酚品红染色30~45min,压片法制片。用显微摄像系统(Olympus BX53-DP72)选取着丝粒清晰、分散良好的中期分裂细胞相进行显微拍照。每个材料选取30个以上良好的中期分裂细胞相进行染色体计数,并选出5个最佳的中期分裂细胞相进行核型分析。核型分析按李懋学等的标准[18],染色体命名按A.Levan等的方法[19],核型分类按G.L.Stebbins的标准[20]。
2 结果与分析
对10种淫羊藿植物的根尖有丝分裂中期细胞进行染色体计数和核型分析,结果显示,该10种植物染色体数均为12条(二倍体,2n=2x=12),每个物种均有1对随体染色体,且每个物种的核型均属Stebbins标准[20]的2A型。10种淫羊藿属植物的根尖有丝分裂中期染色体参数见表2,中期染色体形态和配对核型分别见图1~3。
2.1 粗毛淫羊藿(E.acuminatum Franch.)
原产主要分布于中国西南,贵州各县份均有分布,四川、重庆、云南、广西、湖南、湖北也有分布,生于海拔290~2100 m的山坡、沟谷林下或灌丛中。花大,具长距,明显大于内轮萼片,聚伞状圆锥花序,三出复叶,小叶下面密被粗短硬毛。在Stearn分类系统中划分在Epimedium亚属Diphyllon组Dolichocerae系[5]。
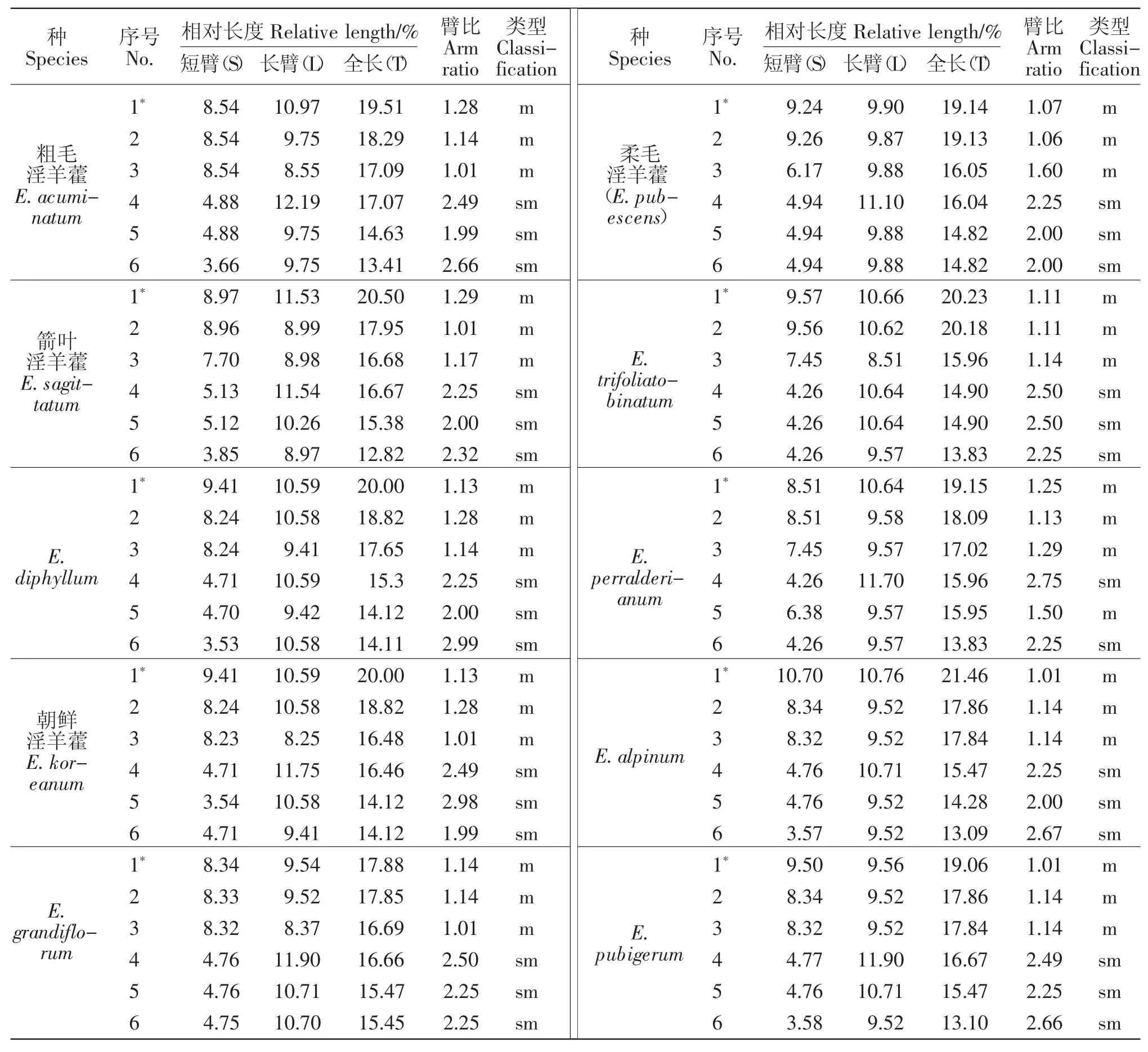
表2 10种淫羊藿属植物根尖细胞有丝分裂中期染色体核型参数Table 2 The karyotype parameters in mitosis metaphase of root tip cells of the ten Epimedium species
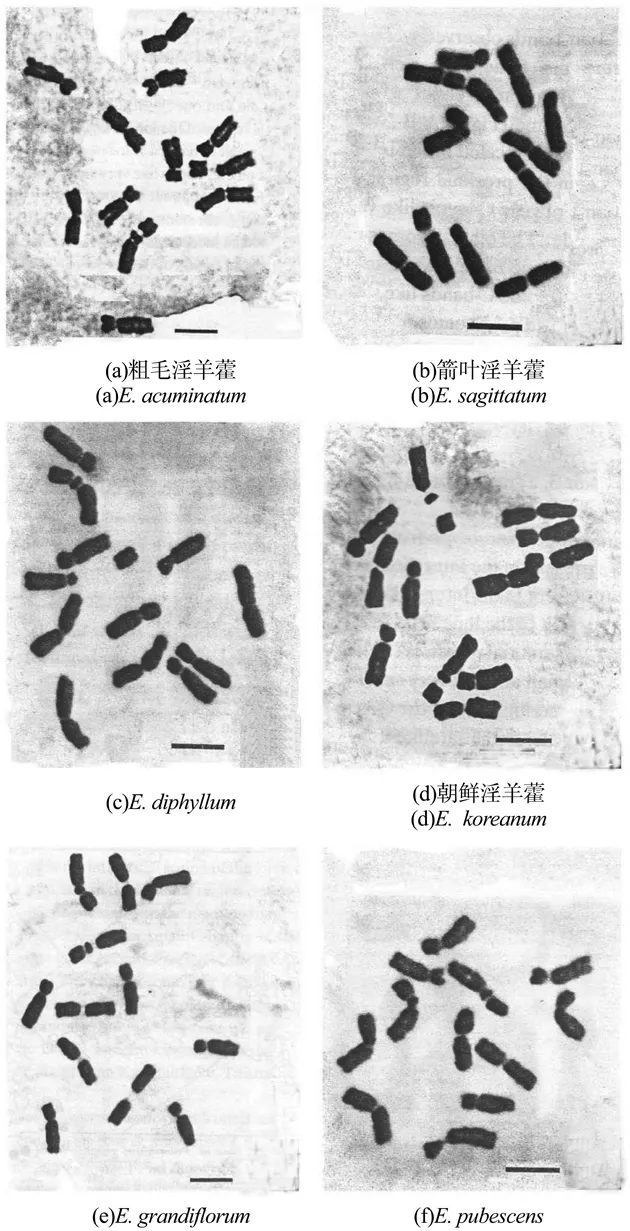
图1 6种淫羊藿属植物的根尖细胞有丝分裂中期染色体Figure 1 The metaphase chromosomes in root tip cells of the six Epimedium species
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.01~2.66,平均臂比值为1.76,最长与最短染色体之比为1.45,核型属2A型。
2.2 箭叶淫羊藿(E.sagittatum Maxim.)
广布种,原产广泛分布于中国各个地区,生于海拔500~1500 m的山坡林下、灌丛及草丛中。花小,具短距,圆锥花序,三出复叶,叶背具粗短硬毛。在Stearn分类系统中划分在Epimedium亚属Diphyllon组 Brachycerae 系[5]。
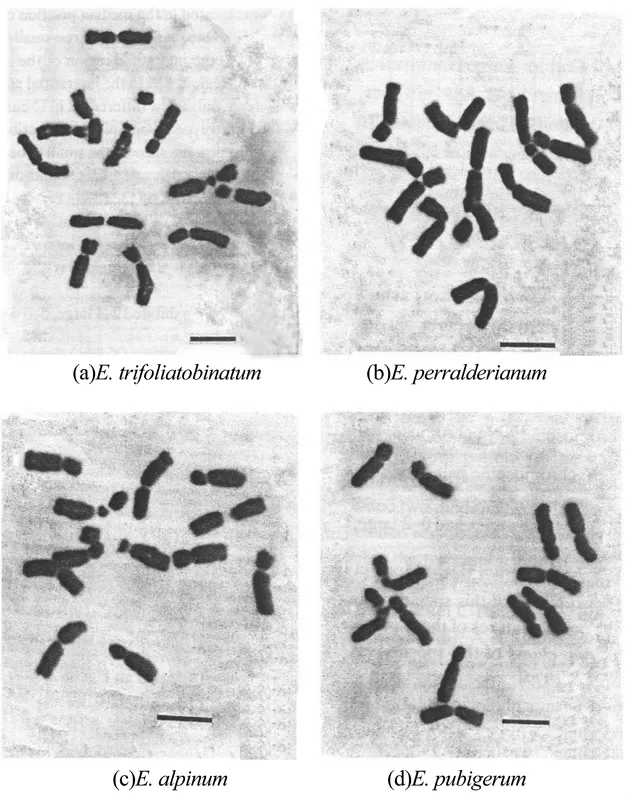
图2 4种淫羊藿属植物的根尖细胞有丝分裂中期染色体Figure 2 The metaphase chromosomes in root tip cells of the four Epimedium species
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.01~2.32,平均臂比值为1.67,最长与最短染色体之比为1.60,核型属2A型。
2.3 E.diphyllum (Morr.et Decne.) Lodd.
原产分布于日本海沿岸,生于海拔500~1500 m的山坡林下、灌丛及草丛中。三出复叶基生或茎生,花瓣无长距,短于或小于内轮萼片,圆锥花序,小叶膜质。在Stearn分类系统中划分在Epimedium亚属Macroceras组[5]。
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.13~2.99,平均臂比值为1.80,最长与最短染色体之比为1.42,核型属2A型。
2.4 朝鲜淫羊藿(E.koreanum Nakai)
原产分布于日本海沿岸,朝鲜,中国吉林、辽宁、浙江、安徽等地区。生于海拔400~1500 m的林下或灌丛中。二回三出复叶基生和茎生,小叶纸质,总状花序顶生,具4~16朵花,花大,直径2~4.5cm,花瓣具长距通常远较内萼片长。在Stearn分类系统中划分在Epimedium亚属Macroceras组[5]。
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.01~2.98,平均臂比值为1.81,最长与最短染色体之比为1.42,核型属2A型。

图3 10种淫羊藿属植物的根尖细胞有丝分裂中期染色体核型Figure 3 The karyotypes of root tip cells of the ten Epimedium species
2.5 E.grandiflorum Morr.
原产分布于日本、朝鲜北部,生于海拔400~1500m的林下或灌丛中。二回三出复叶基生或茎生,小叶纸质,卵形,花大,直径2~4.5cm,总状花序顶生,具4~16朵花,花瓣通常远较内萼片长。在Stearn分类系统中划分在Epimedium亚属Macroceras组[5]。
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.01~2.50,平均臂比值为1.72,最长与最短染色体之比为1.16,核型属2A型。
2.6 柔毛淫羊藿(E.pubescens Maxim.)
原产分布于中国陕西、甘肃、湖北、四川、河南、贵州、安徽等地,生于海拔300~2000 m的林下、灌丛中、山坡地边或山沟阴湿处。一回三出复叶基生或茎生,茎生叶2枚对生,小叶3枚,背面密被绒毛,花小,圆锥花序具30~100余朵花,花瓣远较内萼片短。在Stearn分类系统中划分在Epimedium亚属Diphyllon组Brachycerae系[5]。
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.06~2.25,平均臂比值为1.67,最长与最短染色体之比为1.29,核型属2A型。
2.7 E.trifoliatobinatum (Koidz.)Koidz
原产分布于日本、朝鲜北部,生于海拔400~1500m的林下或灌丛中。三出复叶基生或茎生,小叶革质,花大,直径 2.5~4.5cm,总状花序顶生,具4~10朵花,花瓣具长距明显较内萼片长。在Stearn分类系统中划分在Epimedium亚属Macroceras组[5]。
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.11~2.50,平均臂比值为1.77,最长与最短染色体之比为1.46,核型属2A型。
2.8 E.perralderianum Coss.
原产分布于非洲阿尔及利亚东北部,生长于海拔1200~1500 m的栎树林和雪松林的山坡。小叶心形、边缘密被锯齿状小刺,花尺寸中等,直径1.5~2.3cm,黄色,花瓣2.5mm长,短于内轮萼片。在Stearn分类系统中划分在Rhizophyllum亚属[5]。
核型公式为 2n=2x=8m(2SAT)+4sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.13~2.75,平均臂比值为1.70,最长与最短染色体之比为1.38,核型属2A型。
2.9 E.alpinum L.
淫羊藿属的模式种,原产分布于阿尔卑斯山脉,生于海拔1000~2000 m的山坡上。花较小,具短距,萼片多为深红色;总状花序;三出复叶,叶背无毛着生。在Stearn分类系统中划分在Epimedium亚属Epimedium 组[5]。
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.01~2.67,平均臂比值为1.70,最长与最短染色体之比为1.64,核型属2A型。
2.10 E.pubigerum (DC.) Morr.et Decne.
原产分布于地中海及阿尔卑斯山脉地区,生于海拔600~1200 m的山坡上。花较小而多,具短距,复总状花序,三出复叶,小叶膜质。在Stearn分类系统中划分在Epimedium亚属Epimedium组[5]。
核型公式为 2n=2x=6m(2SAT)+6sm,1、2、3 号为中部着丝粒染色体,4、5、6号为近中着丝粒染色体,1号具有中间随体。染色体臂比的变异幅度为1.01~2.66,平均臂比值为1.78,最长与最短染色体之比为1.45,核型属2A型。
3 讨论与结论
笔者自2007年以来即开展了大量的淫羊藿属植物细胞学研究,2007年分别报道了6种淫羊藿属植物核型[1]和12种淫羊藿属植物核型分析[12],2010年报道了18种淫羊藿属植物核型[7]和14种淫羊藿属植物45S和5S rDNA中期染色体定位[16]。在此基础上,本研究报道了来自中国、日本、美国、德国等广泛区域的10个淫羊藿植物的核型,其中E.grandiflorum,E.diphyllum,E.trifoliatobinatum和E.perralderianum等4个种的核型系首次报道。研究结果使该属植物细胞学核型分析研究更加系统全面,为该属植物的系统分类、起源进化等相关研究提供了更为全面的细胞学证据。
研究结果显示,供试的10种淫羊藿植物染色体数均为12条,基数均为6,均为二倍体(2n=2x=12),且每个物种第1对体染色体均具有随体染色体,这与前人的研究报道结果一致[12-17]。该属植物除印江淫羊藿(E.yinjiangense M.Y.Sheng&Tian X.J.)报道为四倍体(2n=4x=24)外[21],目前有细胞学研究报道的物种均为二倍体。系统进化研究表明,具有对称核型的物种在进化上属于古老原始类群,进化的类群在核型上表现明显的不对称[22]。本研究供试的10种淫羊藿属植物的核型均属Stebbins标准[20]的2A型,典型的对称性核型,与前人对该属植物核型分析的结果一致[12-17],进一步证实了淫羊藿属在进化上是一个较为保守的植物类群[16]。
本研究供试的10种淫羊藿属植物核型公式,除 E.perralderianum 为 2n=2x=8m(2SAT)+4sm,其他9个种均为2n=2x=6m(2SAT)+6sm。结合前人对该属植物的核型分析结果[1,12,16],可以看出淫羊藿属植物核型稳定、种间核型较一致。尽管如此,核型分析在该属植物属下分类与系统进化研究仍具有明显意义。供试材料E.perralderianum属下划分在Rhizophyllum亚属,而其他9个种划分在Epimedium亚属[5],核型研究结果支持这一形态学上亚属的划分。而Epimedium亚属内各类群的划分,尤其是Diphyllon组类群间的划分和关系,还无法从核型分析得到支撑证据,有待于开展该属植物染色体分带、rDNA等特征基因染色体定位等更深入的细胞学研究。
应用核型分析对淫羊藿属植物属下类群进行种类鉴定、亲缘关系阐明十分困难,核型系统学的意义有限,但在属上的分类级别的系统进化研究具有重要的参考价值。与小檗科其他类群比较,山荷叶属(Diphylleia)、八角莲属(Dysosma)、足叶草属(Podophyllum)、桃儿七(Sinopodophyllum)属等 4个属植物的核型公式分别为2n=2x=8m(2SAT)+2st+2t、2n=2x=8m(2SAT)+2st(2SAT)+2t、2n=2x=6m(4SAT)+2sm+2t 和 2n=2x=8m(2SAT)+2st(2SAT)+2t[23-24],淫羊藿属植物的核型与上述类群具有明显差异,且淫羊藿属植物核型明显具有对称性,反映出淫羊藿属植物在小檗科明显是一个起源上更古老的类群,且与其他属植物的亲缘关系较远[25]。这一结果被孢粉学[26-27]、分子标记[28-31]、同工酶[32]及化学分类学[3-4]等研究得到很好的支持。
[1]盛茂银,陈庆富.六种贵州淫羊藿属植物核型分析[J].广西植物,2007,27(3):440-443.
[2]李时珍.本草纲目(1977再版)[M].北京:人民卫生出版社,1977:750-751.
[3]KOGA S,SHOYAMA Y,NISHIOKA I.Studies on Epimedium species:flavonol glycosides and isozymes[J].Biochemical Systematics and Ecology,1991,19:315-318.
[4]SHENG M Y,CHEN Q F,YANG Q X.Variation in icariin and flavonoid contents of barrenwort accessions native to Guizhou,China[J].Biochemical Systematics and Ecology,2008,36:719-723.
[5]STEARN W T.The Genus Epimedium and other herbaceous Berberidaceae[M].Portland:Timber Press,2002.
[6]ZHANG M L,CHRISTIAN H U,JOACHIM W K.Phylogeny and biogeography of Epimedium/Vancouveria(Berberidaceae):Western North American-East Asian disjunctions,the origin of European mountain plant taxa,and East Asian species diversity[J].Systematic Botany,2007,32:81-92.
[7]SHENG M Y,WANG L J,TIAN X J.Karyomorphology of eighteen species of genus Epimedium(Berberidaceae)and its phylogenetic implications[J].Genetic Resources and Crop Evolution,2010,57(8):1165-1176.
[8]郭宝林,裴利宽,肖培根.淫羊藿属植物黄酮类化合物的分类学意义再探[J].植物分类学报,2008,46(6):874-885.
[9]NAKAI R,SHOYAMA Y,SHIRASHI S.Genetic characterization of Epimedium species using random amplified polymorphic DNA (RAPD)and PCR-restriction fragment length polymorphism(RFLP) diagnosis[J].Biological&Pharmaceutical Bulletin,1996,19:67-70.
[10]SUN Y.Characterization of medicinal Epimedium species by 5S rRNA gene spacer sequencing[J].Planta Medica,2004,70:287-288.
[11]SUN W,HUANG W J,LI Z N,et al.Functional and evolutionary analysis of the AP1/SEP/AGL6 superclade of MADS-box genes in the basal eudicot Epimedium sagittatum[J].Annals of Botany,2014,113(4):653-668.
[12]盛茂银,陈庆富.淫羊藿属十二个种的核型[J].云南植物研究,2007,29(3):309-315.
[13]TANAKA R,TAKAHASHI C.Comparative karyotype analysis in Epimedium species by C-banding(1)E.sempervirens var.hypoglaucum and E.perralderianum[J].Journal of Japanese Botany,1981,56:17-24.
[14]TAKAHASHI C.Karyomorphological studies on speciation of Epimedium and its allied Vancouveria with special reference to C-bands[J].Journal of Science of the Hiroshima University,1989,22(2):159-269.
[15]ZHANG Y J,DANG H S,MENG A P,et al.Karyomorphology of Epimedium(Berberidaceae)and its phylogenetic implications[J].Caryologia,2008,61(3):283-293.
[16]SHENG M Y,WANG L J.Chromosomal localization of 45S and 5S rDNA in 14 species and the implications for genome evolution of genus Epimedium[J].Plant Systematics and Evolution,2010,290:65-73.
[17]严福林,何顺志,徐文芬,等.七种中国淫羊藿属药用植物细胞分类学研究[J].广西植物,2016,36(9):1039-1045.
[18]李懋学,陈瑞阳.核型分析标准化的问题[J].武汉植物学报,1985,3(4):297-302.
[19]LEVAN A,FREDGA K,SANDBERG A A.Nomenclacture for centromeric position on chromosomes[J].Hereditas,1964,52:201-220.
[20]STEBBINS G L.Chromosomal evolution in higher plants[M].London:Edward Arnold Ltd,1971.
[21]SHENG M Y,TIAN X J.A new species of Epimedium(Berberidaceae)with 24 chromosomes from Guizhou,China[J].Novon,2011,21:262-265.
[22]李懋学,张赞平.植物染色体及其研究技术[M].北京:中国农业出版社,1996:3-40.
[23]李林初.八角莲及其近缘的核型和演化的研究[J].云南植物研究,1986,8(4):451-457.
[24]马绍宾,胡志浩.小檗科鬼臼亚科植物的核型研究[J].云南植物研究,1996,18(3):325-330.
[25]李超,赵杰,李明.基于形态性状的淫羊藿属(小檗科)的分支分析[J].植物资源与环境学报,2014,23(4):111-113.
[26]张金谈,王萍莉.小檗科花粉形态研究[J].植物分类学报,1983,21(2):130-141.
[27]王悦云,孙庆文,杨雯,等.中国淫羊藿属药用植物花粉特征及其分类学意义[J].西北植物学报,2015,35(7):1367-1377.
[28]王艇,苏应娟,朱建明,等.部分小檗科植物的RAPD分析[J].植物研究,2001.21(3):428-43l.
[29]KIM Y D,KIM S H,KIM C H,et al.Phylogeny of Berberidaceae based on sequence of the chloroplast gene ndhF[J].Biochemical Systematics and Ecology,2004,32:291-301.
[30]ZHANG Y J,YANG L L,CHEN J J,et al.Taxonomic and phylogenetic analysis of Epimedium L.based on amplified fragment length polymorphisms[J].Scientia Horticulturae,2014,170:284-292.
[31]ZHANGYJ,DULW,LIUA,etal.Thecompletechloroplastgenome sequences of five Epimedium species:lights into phylogenetic andtaxonomicanalyse[sJ].FrontiersinPlantScience,2016,7:306.
[32]SHENG M Y,WANG L J,XIONG K N.Isozyme diversity and genetic structure of buckwheat populations[J].Biochemical Systematics and Ecology,2011,39:458-470.
Karyotype Analysis of Ten Epimedium Species and Its Taxonomic Implications
WANG Lin-jiao1,2,TIAN jing1,2,SHENG Mao-yin1,2,3*
(1.Institute of Karst Research,Guizhou Normal University,Guiyang 550001,China;2.National Engineering Research Center for Karst Rocky Desertification Control,Guiyang 550001,China;3.GuizhouEngineeringLaboratoryforKarstRockyDesertificationControlandDerivativeIndustry,Guiyang550001,China)
【Objective】The aim of this study was to analyse the karyotype of ten Epimedium species,and to offer some important evidences for the further studies on phylogenesis,genetics and evolution of Epimedium genus.【Method】The metaphase chromosomes of the root tip cells of ten Epimedium species were studied by traditional chromosome tabletting technique.【Results】Ten Epimedium species used in this study were diploids with 12 chromosomes(2n=2x=12).Their karyotypes were symmetric(2A)and similar to each other,with one pair of middle satellite chromosomes.The karyotype formula of E.perralderianum was 2n=2x=8m(2SAT)+4sm but the other 9 species were 2n=2x=6m(2SAT)+6sm.【Conclusion】The results supported the conclusion that Epiemdium genus was divided into two subgenus(Epimedium and Rhizophyllum),suggesting that Epimedium is an old taxa and is far relation with other taxa in family Berberidaceae.Although karyotype analysis only provide limited data for systematic studies under genus Epimedium,it can offer important and significant evidences for the phylogenesis studies between genus Epimedium and other taxa in family Berberidaceae.
Epimedium;karyotype analysis;phylogenesis;origin;evolution
Q949.746.8;R282.2
A
1000-2650(2017)03-0389-07
10.16036/j.issn.1000-2650.2017.03.016
2017-06-17
国家“十三五”重点研发计划项目(2016YFC0502603);贵州省科技合作计划重点项目(黔科合LH字[2015]7779号)。
王霖娇,硕士,讲师。*责任作者:盛茂银,博士,教授,主要从事特色植物利用与喀斯特生态研究,E-mail:shmoy@163.com。
(本文审稿:唐宗祥;责任编辑:巩艳红;英文编辑:徐振锋)
