燕山和太行山过渡区植物多样性垂直变化特点
2017-11-10丛明旸陈国平陈宝政孙丰宾
丛明旸 曹 迪 陈国平 陈宝政 孙丰宾
(1.河北科技师范学院生命科技学院,秦皇岛 066600; 2.黑龙江八一农垦大学生命科学技术学院,大庆 163319; 3.南开大学生命科学学院,天津 300071; 4.黑龙江八一农垦大学农学院,大庆 163319)
燕山和太行山过渡区植物多样性垂直变化特点
丛明旸1曹 迪2陈国平3陈宝政4孙丰宾4
(1.河北科技师范学院生命科技学院,秦皇岛 066600;2.黑龙江八一农垦大学生命科学技术学院,大庆 163319;3.南开大学生命科学学院,天津 300071;4.黑龙江八一农垦大学农学院,大庆 163319)
为明确山脉过渡区森林群落的建群种,以及验证植物多样性是否符合“中间高度膨胀”理论,沿天津最高峰聚仙峰设置海拔样带,分析得出:(1)乔木、灌木和草本层优势种垂直变化规律不同。乔木层物种组成具有变化规律,研究区下部是槲栎和栓皮栎,随海拔升高核桃楸占优势并混生油松,然后蒙古栎占优势地位,至山顶发展为紫椴和蒙古栎混交林,槲栎和栓皮栎为伴生种,可见栎属植物不仅在各海拔段均有分布,而且优势地位显著,是该区群落的建群种;灌木层以小花溲疏为各海拔段绝对优势种;草本层优势种无规律。(2)α多样性指数具有垂直变化规律。物种多样性S在600~700 m(中低海拔)最高;Simpson指数在800~900 m(中高海拔)最高,群落信息量和复杂度达到最大值;Shannon-Wiener和Pielou指数均为双峰格局,数值在700~800 m(中海拔)低于两侧。因此,燕山和太行山脉过渡区聚仙峰的物种多样性不支持“中间高度膨胀”理论。研究结果和结论丰富了山脉过渡区生态学理论,也提供了北方山地植物多样性信息。
过渡区;植被;重要值;物种多样性;海拔梯度
林分结构决定林分生态服务功能,树种多样性是林分非空间结构的重要组成部分,是反映植物群落稳定程度的重要指标,可以通过计算物种多样性指数量化非空间结构[1],从而揭示过渡区森林植物多样性变化规律,有利于发挥森林生态系统服务功能。植物多样性的海拔格局作为其中一项重要内容,一直是生态学家研究的兴趣点[2],也是保护生物多样性的基础。比较不同山脉物种多样性垂直格局,有利于归纳海拔格局的普适性规律,为理解森林生态系统的生态过程提供新的机理,也是发挥生态系统服务功能和林业可持续发展的内在要求[3]。国内外对山地植物多样性沿海拔变化已有大量研究,主要集中在人为干扰小、生境多样化的山脉。然而,现有研究却较少关注两山脉过渡汇合区。过渡区具有生态脆弱性,其环境容量小,抗干扰能力弱,若植被长势欠佳,甚至会导致水土流失。
天津八仙山国家级自然保护区具有全球同纬度罕见的天然林区,是华北多雨中心和于桥水库水源涵养地,对维护和改善天津生态环境、稳定华北平原农业生态系统起至关重要作用。保护区地处暖温带到温带、湿润到半干旱、华北到东北及内蒙古的复合过渡带,是燕山和太行山两大山脉的汇合处,特殊地理位置促进来自两条山脉不同植物区系的融合,决定了植被具有过渡性和复杂性。以往对该区植被的研究较为薄弱,多数研究集中在植物区系方面。比如徐华鑫[4]、韩英兰[5]、王天罡[6]、沙汀[7]等专家学者先后对维管植物区系进行分析,证实该区地理成分复杂,以华北植物区系为主,具有典型北温带性质;本研究团队在近几年的调查中发表植物新记录[8],为该区植物区系组成添加新成员。然而,有关过渡区植被多样性的研究鲜有报道,仅有的报道是针对典型森林群落分析其物种组成及多样性[9]。国内外山地多样性垂直格局主要有以下6种:(1)与海拔负相关;(2)与海拔正相关;(3)单峰分布格局;(4)双峰分布格局;(5)先降低后恒定的“低矮高原”格局;(6)与海拔无关。多数研究结果以(1)格局较为常见[10],符合“中间高度膨胀”理论,即中间海拔高度具有最高的物种多样性[11]。国内外研究学者关于此理论进行大量验证,结果不一致。然而,植物多样性在山脉过渡区是否符合“中间高度膨胀”理论,需要验证和探讨。
聚仙峰是天津市最高峰,位于八仙山国家级自然保护区,分布大面积天然次生林,保持原始森林风貌,是研究过渡区植物多样性垂直分布特点的天然基地。本研究沿聚仙峰设置垂直样带,拟回答以下2个科学问题:(1)山脉过渡交汇区森林生态系统群落建群种是什么,乔木、灌木和草本层优势种沿海拔是否具有变化规律?(2)植物多样性沿海拔梯度具有怎样的变化特点,是否符合“中间高度膨胀”理论?研究结果和结论将为过渡地区生物多样性保护提供理论参考,为森林经营管理提供理论依据。
1 材料与方法
1.1 研究区概况
燕山山脉位于河北省北部,大体呈东西走向;太行山纵跨河北省,呈东北—西南走向[12~13]。八仙山国家级自然保护区地处40°07′24″~40°13′53″N,117°30′35″~117°36′24″E,是燕山南翼和太行山尾翼交汇处,是天津市山峰最多、地势最高、降水最多、生境最优越、植物资源最丰富的地区,研究区聚仙峰海拔1 052 m,森林覆盖率高达95%[14],气候类型为暖温带大陆性季风气候,四季分明,昼夜温差大,雨季较短,集中于7~8月。
1.2 样方设置及外业调查
于2012年6月~2016年8月,采用典型样方法开展外业调查,选取林相整齐、能代表群落特征的地段,按照方精云等人发表的植物群落清查技术规范[15],沿海拔梯度(500~1 000 m)共设置37个大样方(图1)。样方面积为20 m×20 m,调查样方内全部乔木树种;在每个大样方对角位置设置2个10 m×10 m灌木样方,并调查全部灌木物种;在大样方内随机选取4个1 m×1 m样方,并调查全部草本物种。由于地势原因,个别样方面积设置为100,200和600 m2。
外业调查记录的指标有群落类型、样地面积、调查地点、经度、纬度、海拔、地形、森林起源、郁闭度及垂直结构。群落层次按乔木层、灌木层和草本层划分。乔木层记录植株胸径(DBH)≥3 cm的种名、胸径、树高及健康状况;灌木层记录各物种(包括乔木幼苗和幼树)的种名、平均基径、平均高、株数及盖度;草本层记录各物种的种名、盖度、平均高及多度。调查样方概况见表1。
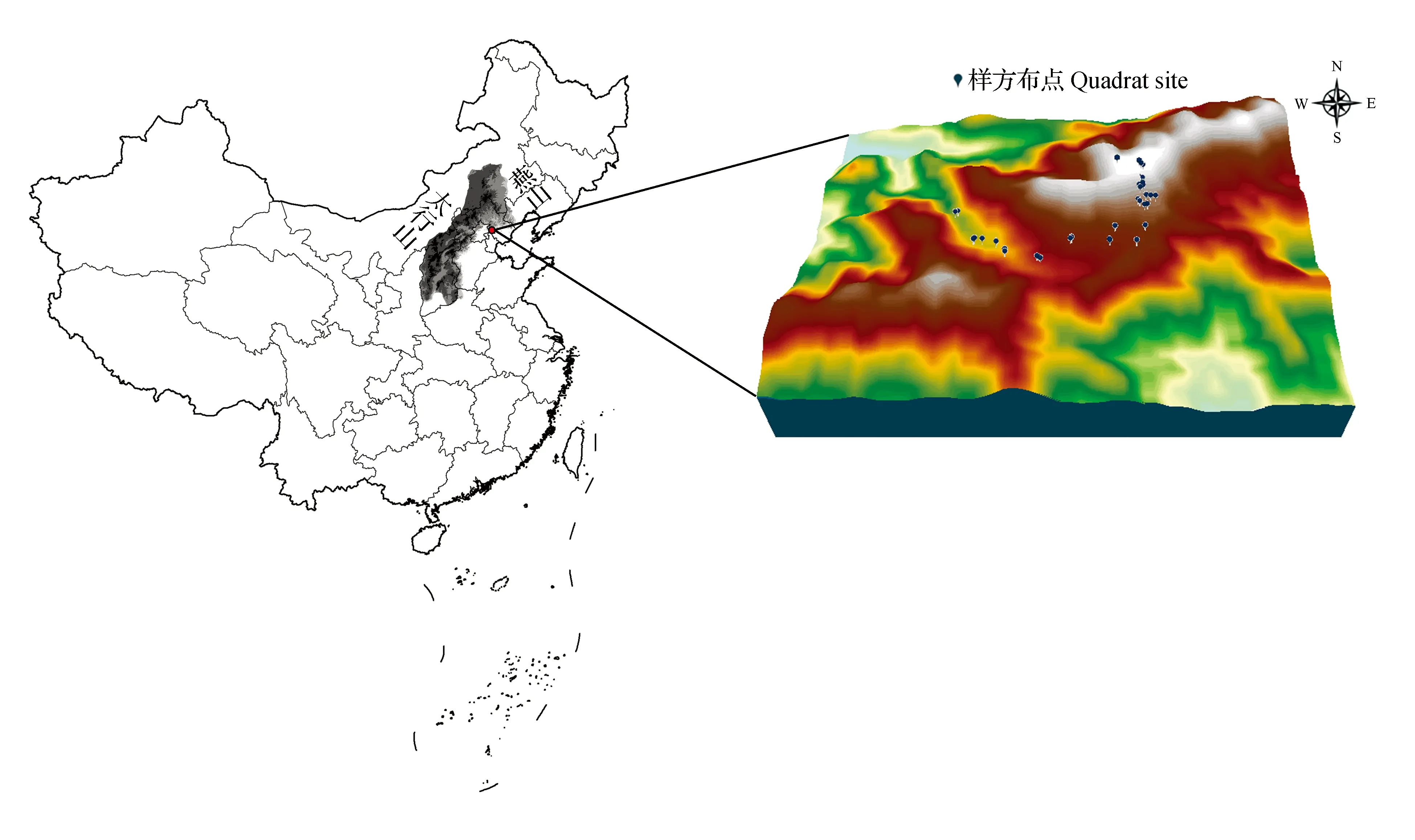
图1 燕山和太行山过渡区聚仙峰样方布设示意图Fig.1 Sketch map and quadrat sites of transitional zone Juxianfeng between Mount. Yan and Mount. Taihang
1.3 数据处理
将500~1 000 m共划分为5个海拔区间:Ⅰ(500~600 m)、Ⅱ(600~700 m)、Ⅲ(700~800 m)、Ⅳ(800~900 m)和Ⅴ(900~1 000 m)。重要值的计算公式如下:
Ⅳ乔木=(相对密度+相对优势度+相对频度)/3
Ⅳ灌木=(相对密度+相对盖度+相对高度)/3
Ⅳ草本=(相对多度+相对盖度+相对高度)/3
采用Biodap(Biodiversity Data Analysis Package)软件计算广泛应用的4个多样性指数,即物种丰富度指数S、Simpson优势度指数、Shannon-Wiener多样性指数和Pielou均匀度指数,再用Origin 9.9.0软件作图。
2 结果与分析
2.1 聚仙峰森林植物群落各层优势种
样方内共记录维管植物51科,109属,157种。群落各层重要值计算结果为:聚仙峰乔木层优势种是壳斗科(Fagaceae)栎属(Quercus)植物,包括蒙古栎(Quercusmongolica)、栓皮栎(Quercusvariabilis)及槲栎(Quercusaliena),表明栎属植物是燕山和太行山过渡区内暖温带落叶阔叶林的优势种;常见伴生树种主要是桦木科(Betulaceae)、胡桃科(Juglandaceae)、椴树科(Tiliaceae)、芸香科(Rutaceae)植物(表2)。灌木层优势种是虎耳草科(Saxifragaceae)植物(表3)。草本层优势种主要是豆科(Leguminosae)、禾本科(Gramineae)、菊科(Compositae)及毛茛科(Ranunculaceae)植物(表4)。
2.2聚仙峰森林植物群落各层优势种沿海拔区间的变化规律
聚仙峰森林植物群落各层优势种的垂直分布规律性各异(表5)。乔木层垂直变化规律最为显著,主要体现在优势种组成上,500~600 m区间是以槲栎和栓皮栎为优势种的落叶阔叶林分布区;600~700 m区间群落以核桃楸(Juglansmandshurica)占优势,油松(Pinustabulaeformis)次之,栓皮栎和槲栎的优势地位逐渐减弱;700~800 m区间除油松林外,核桃楸为单优势种,伴生种主要有蒙古栎及栓皮栎;800~900 m海拔区间的蒙古栎逐步取代核桃楸而成为群落优势地位明显的优势种,稀疏分布的栓皮栎为伴生种,此时油松逐渐退出群落,属于混生有稀疏针叶林的落叶阔叶林群落分布区;900~1 000 m区间为蒙古栎和紫椴(Tiliaamurensis)共优种混交林,而槲栎和栓皮栎稀疏分散生长。再次验证了2.1的研究结果,即该过渡区是以栎属植物占显著优势地位的集中分布区。灌木层的小花溲疏(Deutziaparviflora)在各海拔区段均占绝对优势。草本层的优势种无变化规律。
表2聚仙峰森林植物群落乔木层物种重要值
Table2SpeciesimportancevalueoftreelayerinJuxianfeng

物种Species相对密度Relativedensity相对优势度Relativesignificance相对频度Relativefrequency重要值Importantvalue蒙古栎Quercusmongolica8.165522.33532.355710.9521油松Pinustabulaeformis11.187113.12412.35578.8889栓皮栎Quercusvariabilis9.280611.76461.06017.3684鹅耳枥Carpinusturczaninowii13.20144.07323.41586.8968核桃楸Juglansmandshurica6.115111.73082.59136.8124紫椴Tiliaamurensis5.14399.64102.12015.6350臭檀Euodiadaniellii5.97124.01402.35574.1136白蜡树Fraxinuschinensis7.69781.55542.82694.0267元宝槭Acertruncatum6.29502.31783.06243.8917槲栎Quercusaliena4.17274.31791.88463.4584大叶朴Celtiskoraiensis3.16550.67663.18022.3408糠椴Tiliamandschurica1.94243.18101.41342.1789大果榆Ulmusmacrocarpa2.37411.46082.47352.1028黄檗Phellodendronamurense1.11512.49351.17791.5955桑Morusalba1.33091.13191.88461.4491沙梾Swidabretchneideri1.07910.85761.41341.1167
注:重要值小于1.0000的物种未列出。
Note:Important value of plants less than 1.0000 were not listed.
表3聚仙峰森林植物群落灌木层物种重要值
Table3SpeciesimportancevalueofshrublayerinJuxianfeng

物种Species相对密度Relativedensity相对盖度Relativecoverage相对频度Relativefrequency重要值Importantvalue小花溲疏Deutziaparviflora35.451029.60622.944622.6673大花溲疏Deutziagrandiflora11.74907.47471.06016.7613三裂绣线菊Spiraeatrilobata10.54123.40091.76685.2363大叶朴Celtiskoraiensis3.74907.21043.18024.7132雀儿舌头Leptopuschinensis5.70985.40151.76684.2927
注:重要值小于4.0000的物种未列出。
Note:Important value of plants less than 4.0000 were not listed.
表4聚仙峰森林植物群落草本层物种重要值
Table4SpeciesimportancevalueofherblayerinJuxianfeng

物种Species相对高度Relativeheight相对盖度Relativecoverage相对频度Relativefrequency重要值Importantvalue歪头菜Viciaunijuga22.89294.06081.649085.8082求米草Oplismenusundulatifolius12.60499.85951.295671.2801三脉紫菀Asterageratoides18.12292.57580.942364.9229大叶铁线莲Clematisheracleifolia3.594614.33982.120160.1636升麻Cimicifugafoetida15.91521.76340.117853.3891臭草Melicascabrosa1.234612.51981.531245.8568糙苏Phlomisumbrosa2.40557.15571.413432.9238
注:重要值小于30.0000的物种未列出。
Note:Important value of plants less than 30.0000 were not listed.
2.3聚仙峰森林植物群落物种多样性垂直变化规律
物种丰富度S(图2:A)随海拔升高呈单峰分布格局,且在600~700 m海拔区间物种数目最多。Simpson优势度指数(图2:B)表现为单峰分布格局,群落复杂度均在800~900 m海拔区间最高,表明该区间包含的信息量最大。Shannon-Wiener多样性指数(图2:C)和Pielou均匀度指数(图2:D)表现为几乎一致的“M”型双峰分布格局,中海拔区间700~800 m数值低于两侧(600~700和800~900 m)。
3 讨论
3.1 山脉过渡区建群种及伴生种区系来源
建群种控制着群落的性质和环境,对森林生态系统的稳定性起至关重要作用。海拔是产生山地水热和土壤养分异质性的主要因素,植物群落优势种分异主要源于海拔变化,因此物种组成和多样性也不同。
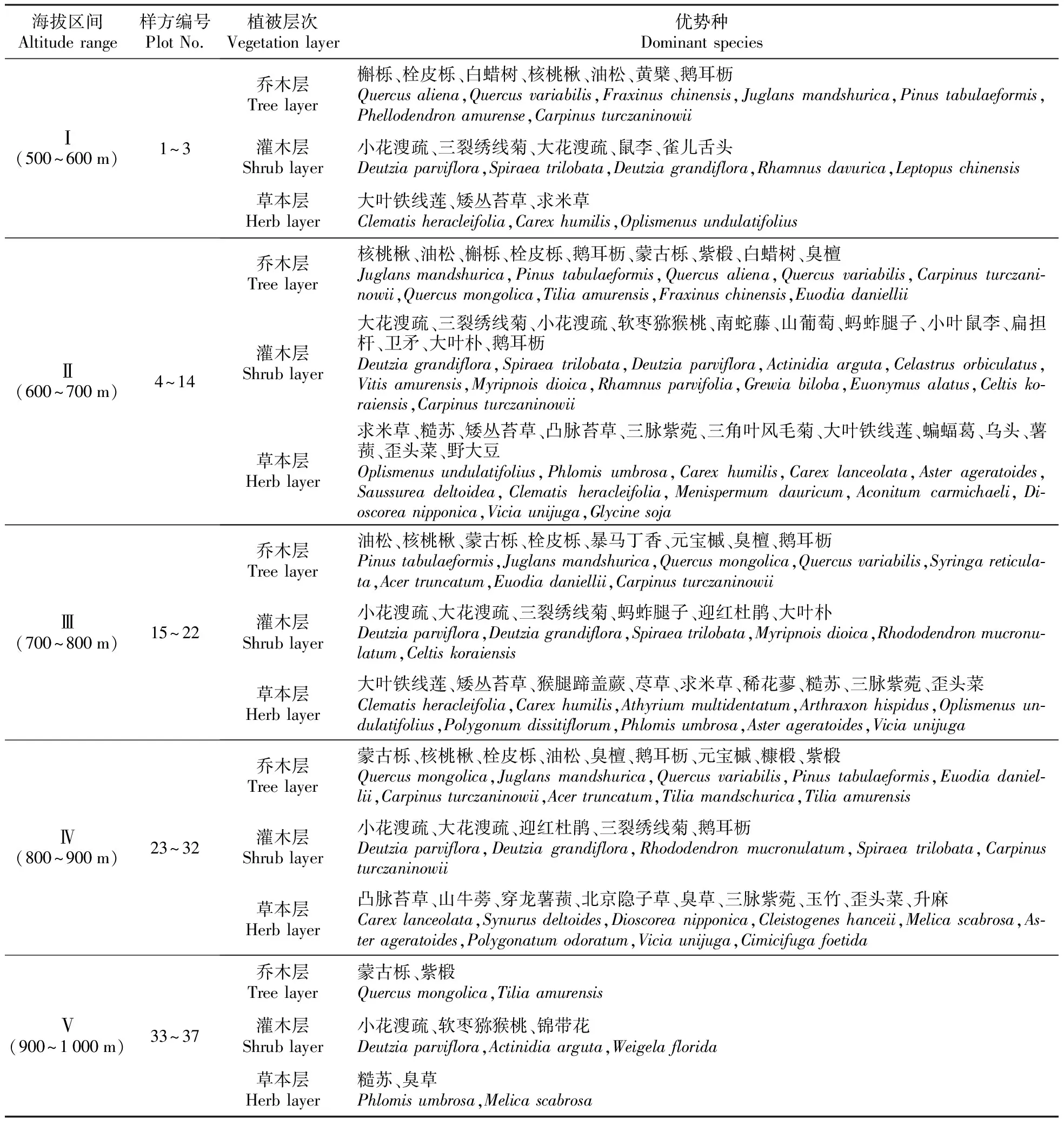
表5 不同海拔区间植物群落乔木、灌木和草本层的优势种

图2 聚仙峰森林群落物种多样性指数垂直变化特点A.物种数目;B. Simpson指数;C. Shannon-Wiener指数;D. Pielou指数 Ⅰ. 500~600 m;Ⅱ. 600~700 m;Ⅲ. 700~800 m;Ⅳ. 800~900 m;Ⅴ. 900~1 000 m Fig.2 Vertical characteristics of diversity indexes of forest communities in Juxianfeng A. Species number; B. Simpson index; C. Shannon-Wiener; D. Pielou index Ⅰ. 500-600 m;Ⅱ. 600-700 m;Ⅲ. 700-800 m;Ⅳ. 800-900 m;Ⅴ. 900-1 000 m
燕山和太行山山脉均具有植被垂直带谱,二者交汇区聚仙峰虽然已是天津最高峰,但是植被却不具备垂直带谱,原因是相对海拔高度差值较小。表5结果表明垂直变化规律主要体现在群落内部的物种组成上,从调查起点到山顶,栎属植物始终是优势种或次优种。自然保护区的设立使早期被破坏的植被受到较好保护,具备优越的水、热、立地条件,现已发展成栎属植物为建群种的暖温带落叶阔叶林,并且在过渡带中发挥至关重要的作用,可能是影响该区生态系统的结构和功能的重要植物类群。桦木科、胡桃科、椴树科、芸香科等植物为常见伴生树种。
相比于燕山和太行山山脉,聚仙峰的群落物种组成与二者存在异同。对于燕山山脉,北部典型群落主要是华北落叶松(Larixprincipis-rupprechtii)人工林和杨桦(Populusdavidiana,Betulaplatyphylla)天然次生林[16];最高峰雾灵山位于燕山中段,以油松、辽东栎、山杨(Populusdavidiana)、白桦(Betulaplatyphylla)为优势种,伴生种有核桃楸、槲栎等[17]。对于太行山脉,小五台山国家级自然保护区位于太行山最北端,优势树种是白桦和红桦(Betulaalbosinensis)[18];太行山中段的乔木主要白桦、沙棘(Hippophaerhamnoides)[19];中南段乔木以栓皮栎、槲栎、核桃楸、山杨、漆树(Toxicodendronvernicifluum)、鹅耳枥为优势种[20];低山丘陵主要是灌丛,如荆条(Vitexnegundovar.heterophylla)、酸枣(Ziziphusjujubevar.heterophylla)等[21];东灵山乔木优势种有核桃楸、黄花柳(Salixcaprea)、棘皮桦(Betuladahurica)、山杨、蒙椴(Tiliamongolica)、坚桦(Betulachinensis)、五角枫(Acermono)、辽东栎等[22]。
由此可见,地理位置越接近过渡区,栎属植物出现频率越增加,聚仙峰栎属植物种源主要来自于燕山山脉中段以南和太行山山脉中南段直至尾翼,由于聚仙峰独特的地理位置,调查中发现了推测可能是栎属的杂交种,还是研究栎属植物遗传多样性的天然基因库。另外,伴生种核桃楸也是来源于两条山脉种源的汇合。
3.2 “中间高度膨胀”理论在山地过渡区的验证
山地多样性研究的一个核心问题就是物种多样性沿海拔的变化规律[23]。用于解释物种多样性垂直分布格局的多种理论中,“中间高度膨胀”理论作为其中的一种,其核心论点是低山地区受水分限制,高海拔地段受热量限制,因此中海拔地区植物多样性最高。
聚仙峰植物物种多样性指数具有垂直变化规律(图2)。物种多样性S和Simpson指数的峰值分别出现在600~700和800~900 m区间,即中低、中高海拔区间群落信息量和物种复杂度达到最大值。Shannon-Wiener多样性指数和Pielou均匀度指数变化趋势相同,均为“M”型双峰格局,中间海拔700~800 m区间的数值低于两端,形成这种格局的原因可能是,该海拔段的优势种油松是成立保护区以前栽种的人工林,其物种组成较为单一,这也从另外一个角度证实了自然植被在物种多样性维持中所起的重要作用。燕山山脉最高峰雾灵山的植物多样性沿海拔变化规律符合“中间高度膨胀”理论[17]。太行山山脉最高峰小五台山[18],以及另一座王屋山[24],其多样性指数符合“中间高度膨胀”理论,而山脉中段群落物种多样性随海拔升高而升高[19]。
由此可见,虽然燕山山脉和太行山脉的最高峰符合“中间高度膨胀”理论,表明物种丰富度在较大的尺度上具有相似性,但是在两者过渡区聚仙峰,多样性不符合“中间高度膨胀”理论。聚仙峰多样性格局的形成原因可能有两方面。一方面,推测是聚仙峰相对海拔高度不大,因此,山顶和低山的水分和光照条件的差异小。比如热带香格里拉县种子植物多样性格局符合该理论,其海拔差近4 000 m[25];再如,对泰山和崂山两山脉的研究发现乔木物种丰富度也符合该理论,海拔差分别为1 500和1 300 m[26];而与本研究结果类似的近期研究是Tanaka等人对日本中部种子植物研究,指出尤其海拔梯度差值小的山地,物种多样性沿海拔呈不对称驼峰型分布,即不符合该理论[27],均表明相对海拔梯度可能是产生“M”型双峰格局的主要原因。另一方面,推测可能是地形因素通过改变光、热、水、土的空间分配,从而影响群落多样性[23]。近期对太行山丘陵生态实验站(114°16′E,37°53′N)区域内山地的研究表明,坡向和坡位对植物物种分布格局和多样性产生重要影响,南坡(阳坡)以耐旱物种居多,北坡(阴坡)以喜湿润的物种为主;中间坡位由于其较高的热量和土壤干燥度,物种多样性最低[28]。对燕山最高峰雾灵山的研究结果表明,低海拔到中高海拔段湿度是主要限制因子,中海拔到山顶热量是限制因子[17]。类似于太行山和燕山地区的结论在本研究中也有体现,因此有理由推测中间海拔700~800 m区间由于坡向以阳坡(南坡)为主(图1),加之油松林下土壤较为干旱和贫瘠,形成目前中间海拔多样性低于两侧的海拔格局。
关于山脉过渡区的研究相对薄弱,后续研究应在相对海拔更高的山体开展,并将环境和土壤因子考虑其中,基于物种生境的相关性分析更加有利于理解山脉过渡区群落构建、物种共存机制,本研究至少为进一步深入研究指明方向。
致谢野外调查过程中,得到八仙山国家级自然保护区管理局的赵铁建、天津市野生动植物保护管理站张小锟的支持,在此谨致谢意!
1.曹小玉,李际平.林分空间结构指标研究进展[J].林业资源管理,2016(4):65-73.
Cao X Y,Li J P.Research progress on indicators of the stand spatial structure[J].Forest Resources Management,2016(4):65-73.
2.Santos L M,Lenza E,Dos Santos J O,et al.Diversity,floristic composition,and structure of the woody vegetation of the Cerrado in the Cerrado-Amazon transition zone in Mato Grosso,Brazil[J].Brazilian Journal of Botany,2015,38(4):877-887.
3.Hooper D U,Chapin Ⅲ F S,Ewel J J,et al.Effects of biodiversity on ecosystem functioning:a consensus of current knowledge[J].Ecological Monographs,2005,75(1):3-35.
4.徐华鑫,张启良,张聪.天津蓟县八仙山自然保护区的特点与功能[J].自然资源,1994(2):74-78.
Xu H X,Zhang Q L,Zhang C.Characteristics and function of Baxianshan natural reserve in Ji Coutry of Tianjin[J].Resources Science,1994(2):74-78.
5.韩英兰,苏卫国.天津市蓟县八仙山自然保护区木本植物区系的研究[J].天津农学院学报,1996,3(4):1-9.
Han Y L,Su W G.Research on the woody plant zone in Baxianshan Mountain Natural Protective area of Ji Country in Tianjin[J].Journal of Tianjin Agricultural College,1996,3(4):1-9.
6.王天罡,邢韶华,林大影,等.天津八仙山自然保护区维管束植物分析[J].河北林果研究,2007,22(2):134-139.
Wang T G,Xing S H,Lin D Y,et al.Floristic analysis of vascular bundle plant in Baxianshan nature reserve,Tianjin[J].Hebei Journal of Forestry and Orchard Research,2007,22(2):134-139.
7.沙汀,张文辉,刘新成.天津市种子植物区系成分分析[J].植物研究,2009,29(1):96-102.
Sha T,Zhang W H,Liu X C.Floristic analysis of seed plants in Tianjin[J].Bulletin of Botanical Research,2009,29(1):96-102.
8.丛明旸,宫乐,张玉婷,等.天津市植物资源新记录Ⅰ[J].南开大学学报:自然科学版,2015,48(3):13-18.
Cong M Y,Gong L,Zhang Y T,et al.New records of plant species in Tianjin[J].Acta Scientiarum Naturalium Universitatis Nankaiensis,2015,48(3):13-18.
9.丛明旸,石会平,张小锟,等.八仙山国家级自然保护区典型森林群落结构及物种多样性研究[J].南开大学学报:自然科学版,2013,46(4):44-52.
Cong M Y,Shi H P,Zhang X K,et al.Analyses on community structure and species diversity of typical forest in Baxianshan National Natural Reserve[J].Acta Scientiarum Naturalium Universitatis Nankaiensis,2013,46(4):44-52.
10.唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20-28.
Tang Z Y,Fang J Y.A review on the elevational patterns of plant species diversity[J].Biodiversity Science,2004,12(1):20-28.
11.黄甫昭,丁涛,李先琨,等.弄岗喀斯特季节性雨林不同群丛物种多样性随海拔的变化[J].生态学报,2016,36(14):4509-4517.
Huang F Z,Ding T,Li X K,et al.Species diversity for various associations along an altitudinal gradient in the karst seasonal rainforest in Nonggang[J].Acta Ecologica Sinica,2016,36(14):4509-4517.
12.严慧玲,刘东明,李作恒,等.层次分析法在河北省太行山区植被恢复植物种类筛选中的运用[J].植物研究,2015,35(5):751-758.
Yan H L,Liu D M,Li Z H,et al.Analytic hierarchy process in plant species selection of revegetation in the Taihang Mountains in Hebei Province[J].Bulletin of Botanical Research,2015,35(5):751-758.
13.李军玲,张金屯.太行山中段植物群落优势种生态位研究[J].植物研究,2006,26(2):156-162.
Li J L,Zhang J T.Niche of dominant species in the Midst of Taihang Mountain[J].Bulletin of Botanical Research,2006,26(2):156-162.
14.陈国平,俎丽红,高张莹,等.八仙山不同立地落叶阔叶林凋落物养分特征及土壤肥力评价研究[J].植物研究,2016,36(6):878-885.
Chen G P,Zu L H,Gao Z Y,et al.The characteristics of forest floor nutrients and soil fertility assessment for deciduous broad-leaved forest with different site conditions[J].Bulletin of Botanical Research,2016,36(6):878-885.
15.方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
Fang J Y,Wang X P,Shen Z H,et al.Methods and protocols for plant community inventory[J].Biodiversity Science,2009,17(6):533-548.
16.李东胜,王丽,许中旗,等.燕山北部山地典型植物群落的物种多样性[J].河北农业大学学报,2014,37(3):44-50.
Li D S,Wang L,Xu Z Q,et al.Species diversity of typical plant communities in the north region of Yanshan Mountain[J].Journal of Agricultural University of Hebei,2014,37(3):44-50.
17.奚为民.雾灵山国家自然保护区森林群落物种多样性研究[J].生物多样性,1997,5(2):121-125.
Xi W M.Study on species diversity of forest in Wuling Mountain natural reserve[J].Biodiversity Science,1997,5(2):121-125.
18.刘增力,郑成洋,方精云.河北小五台山北坡植物物种多样性的垂直梯度变化[J].生物多样性,2004,12(1):137-145.
Liu Z L,Zheng C Y,Fang J Y.Changes in plant species diversity along an elevation gradient on Mt.Xiaowutai,Hebei,China[J].Biodiversity Science,2004,12(1):137-145.
19.李军玲,张金屯.太行山中段植物群落物种多样性与环境的关系[J].应用与环境生物学报,2006,12(6):766-771.
Li J L,Zhang J T.Plant species diversity in the middle part of the Taihang Mountain[J].Chinese Journal of Applied & Environmental Biology,2006,12(6):766-771.
20.唐伟斌.太行山脉东坡中南段种子植物区系初步分析[J].植物研究,2005,25(3):366-372.
Tang W B.A floristic analysis of the seed plants in the middle-south area of east slope of Taihang Mountains[J].Bulletin of Botanical Research,2005,25(3):366-372.
21.曾歆花,张万军,宋以刚,等.河北太行山低山丘陵区植被恢复过程中物种多样性与土壤养分变化[J].生态学杂志,2013,32(4):852-858.
Zeng X H,Zhang W J,Song Y G,et al.Species diversity and soil nutrient dynamics along a chronosequence of vegetation restoration in Taihang Mountains hilly region,Hebei province of North China[J].Chinese Journal of Ecology,2013,32(4):852-858.
22.马克平,叶万辉,于顺利,等.北京东灵山地区植物群落多样性研究Ⅷ.群落组成随海拔梯度的变化[J].生态学报,1997,17(6):593-600.
Ma K P,Ye W H,Yu S L,et al.Studies on plant community diversity in Dongling Mountain,Beijing,China VIII.Variation of composition along elevational gradient[J].Acta Ecologica Sinica,1997,17(6):593-600.
23.方精云,沈泽昊,崔海亭.试论山地的生态特征及山地生态学的研究内容[J].生物多样性,2004,12(1):10-19.
Fang J Y,Shen Z H,Cui H T.Ecological characteristics of mountains and research issues of mountain ecology[J].Biodiversity Science,2004,12(1):10-19.
24.郑敬刚,张有福,王云,等.太行山中段植被分布特征及其多样性研究[J].河南科学,2009,27(3):292-294.
Zheng J G,Zhang Y F,Wang Y,et al.The characteristics of plant distribution and diversity in the middle section of Taihang Mountain[J].Henan Science,2009,27(3):292-294.
25.李瑞年,杜凡,李云琴,等.香格里拉县种子植物多样性海拔分布格局[J].西南林业大学学报,2013,33(6):13-18.
Li R N,Du F,Li Y Q,et al.Altitudinal distribution pattern of seed plant diversity in Shangri-la county[J].Journal of Southwest Forestry University,2013,33(6):13-18.
26.Zhang W X,Huang D Z,Wang R Q,et al.Altitudinal patterns of species diversity and phylogenetic diversity across temperate mountain forests of northern China[J].PLoS One,2016,11(7):e0159995,doi:10.1371/journal.pone.0159995.
27.Tanaka T,Sato T.Species richness of seed plants and ferns along a temperate elevational gradient in central Japan[J].Plant Ecology,2014,215(11):1299-1311.
28.Zeng X H,Zhang W J,Song Y G,et al.Slope aspect and slope position have effects on plant diversity and spatial distribution in the hilly region of Mount. Taihang,North China[J].Journal of Food,Agriculture & Environment,2014,12(1):391-397.
Research Foundation for Introduction of Talent(XYB2016-01);National Natural Science Foundation of China(31400444);Heilongjiang Province Training Programs of Innovation and Entrepreneurship for Undergraduates(201410223022)
introduction:CONG Ming-Yang(1986—),female,lecturer,Ph.D,Major in botany and ecology.
date:2017-01-03
VerticalCharacteristicsofPlantDiversityinTransitionBetweenMount.YanandMount.Taihang
CONG Ming-Yang1CAO Di2CHEN Guo-Ping3CHEN Bao-Zheng4SUN Feng-Bin4
(1.College of Life Science and Technology,Hebei Normal University of Science & Technology,Qinhuangdao 066600;2.College of Life Science and Technology,Heilongjiang Bayi Agricultural University,Daqing 163319;3.College of Life Sciences,Nankai University,Tianjin 300071;4.Agronomy College,Heilongjiang Bayi Agricultural University,Daqing 163319)
We clarified forest constructive species in transition area, and put ‘Intermediate expansion hypothesis’ to the test. We took the highest peak Juxianfeng in Tianjin of China as study site, and set transect sampling along elevation vertically. The results were as follows: (1) Change rules of dominant species in every layer differed from each other. In tree layer, dominant species in lower area wereQuercusalienaandQ.variabilis. Increasing with altitude,Juglansmandshuricagradually became dominant, mixed withPinustabuliformis.Q.mongolicaplayed an important role in community. Until top of the mountain, the plant communities wereTiliaamurensis-Q.mongolicamixed forests. However,Q.alienaandQ.variabilisbecame companion species at the same time.Quercusdistributed everywhere and were keystone species in this transition area.Deutziaparviflorawas absolute dominant species in shrub layer within all altitude. Dominant species in herb layer showed disordered. (2) α diversity indexes changed vertically. Total species richnessSreached the maximum in 600-700 m. The peak of Simpson index appeared in 800-900 m which was in middle high altitude, suggesting more information and complexity. Shannon-Wiener and Pielou indexes showed the same law that was bimodal pattern, namely numerical value in 700-800 m was lower than both two sides. Therefore, our results didn’t support ‘Intermediate expansion hypothesis’ in transition area. Our study enriched the ecology theory in mountain transition region, and provided plant diversity information of the north mountain in China.
transition zone;vegetation;importance value;species diversity;elevation gradient
学成、引进人才科研启动金(XYB2016-01);国家自然青年基金(31400444);黑龙江省教育厅(201410223022)
丛明旸(1986—),女,讲师,博士研究生,现主要从事植物学和生态学研究。
2017-01-03
Q145+.2
A
10.7525/j.issn.1673-5102.2017.05.006
