OT杂种系百合品种‘Cocossa’小孢子败育细胞学研究
2017-11-10赵玉萍何祥凤王文和冷平生张克中
吴 杰 赵玉萍 何祥凤 王文和 冷平生 张克中
OT杂种系百合品种‘Cocossa’小孢子败育细胞学研究
吴 杰 赵玉萍 何祥凤 王文和*冷平生 张克中
以OT百合‘Cocossa’为材料,采用醋酸洋红染色烤片法和石蜡切片法,对该品种花粉母细胞减数分裂过程以及绒毡层在花粉母细胞发育过程中的变化进行系统的细胞学观察。研究发现:(1)当百合花蕾长度在2.5~4.5 cm时,花粉母细胞处在减数分裂时期。(2)在花粉母细胞减数分裂的过程中,出现了较多染色体异常现象:中期Ⅰ出现染色体环;后期Ⅰ出现了滞后染色体和染色体桥;在末期Ⅰ和末期Ⅱ细胞出现不均等分裂和微核;处于同一花蕾和同一花药的花粉母细胞减数分裂具有不同步性,能同时观察到2个不同时期的分裂相;形成畸形、无核、多核的异常小孢子。(3)在末期Ⅱ和单核花粉粒时期,绒毡层延迟降解,引起多核花粉粒、畸形花粉粒以及花粉粒自溶现象。结果表明:OT百合‘Cocossa’小孢子败育是花粉母细胞异常减数分裂与绒毡层异常降解共同导致的。
OT百合;减数分裂;绒毡层;小孢子败育
百合(Liliumspp.)是目前世界上最受欢迎的花卉种类之一,因其花色丰富、花形多变,且部分百合有浓郁的香味。OT百合杂种系是东方百合与喇叭百合杂交而成,其花苞大、茎杆粗壮、抗逆性强等特点越来越受到百合种植者和消费者的青睐。随着近年来育种者不断培育出OT百合新品种,如‘火凤凰’、‘金龙’[1]等,OT百合逐渐有替代花卉市场上原有的东方、LA百合切花,成为百合切花市场上的主导花卉的趋势。OT百合‘Cocossa’是由2个东方百合基因组(O)和1个喇叭百合基因组(T)组成的异源三倍体[2]。三倍体植物并非完全不育,国内外的研究学者分别在不同的植物上成功的进行了验证,Harder[4]等、Winton[3]等利用三倍体美洲山杨的花粉给正常二倍体山杨授粉并成功获得了四倍体植株。梁国鲁[5]等对三倍体枇杷品种进行花粉离体和活体萌发实验,观察到小部分花粉萌发。然而,三倍体植物在减数分裂过程,易发生染色体异常现象,对植物发育和杂交育种产生不良影响。王玉蛟[6]等通过对三倍体芍药花粉母细胞的细胞学观察发现,染色体异常对配子的育性有直接影响,一般导致配子不育,需进一步挖掘其育种潜力。目前国内对百合小孢子发育的相关报道主要集中在王百合[7]、兰州百合[8]和东方百合的一些品种[9],而三倍体OT百合小孢子发育的相关研究很少。本文对OT百合‘Cocossa’小孢子发育过程中减数分裂异常现象和绒毡层变化方面做了观察和分析,揭示了两者与花粉败育之间的关系,以期在育种中利用三倍体百合的优良性状,采取有效措施,获得一定数量的可育雄配子提供细胞学基础。
1 材料和方法
1.1 材料
试验材料百合品种‘Cocossa’ 2012年种植于北京农学院北农科技园温室内,4月中旬至5月中旬从花蕾长度0.5 cm开始,每隔3 d切取不同发育时期的花蕾一次,取材时间为上午8:00~10:00,采集花蕾长度为0.5~6.0 cm。所取材料的一半用现配置的卡诺固定液(无水乙醇∶冰醋酸=3∶1)固定12 h后,换作70%的乙醇于4℃冰箱保存;另一半材料用FAA固定并保存。
1.2 方法
1.2.1 花蕾、花药长度与小孢子发育的关系
将采集到的花蕾和花药准确量取长度,并观察其颜色。
1.2.2 花粉母细胞减数分裂的观察
将所取材料的一半用卡诺固定液(乙醇∶醋酸=3∶1)固定的24 h,之后将材料转入70%的酒精中,4℃保存以备用。将固定好的材料用蒸馏水洗净,置于载玻片上,直接挤出花药内含物,用醋酸洋红染色并压片,用酒精灯外焰烤1~3 s,之后在光学显微镜下进行花粉母细胞减数分裂过程观察。
1.2.3 绒毡层细胞的观察
将所取材料的另一半用FAA固定液(90mL 70%乙醇+5mL 37%~40%甲醛+5mL乙酸)固定,将固定好的材料用常规石蜡切片法制片,采用番红—固绿双重染色法染色,用加拿大树胶封片,切片厚度为8 μm。
2 结果与分析
2.1 花蕾、花药长度与花粉母细胞发育的关系
研究发现花蕾、花药长度以及花药颜色与花粉母细胞的发育程度紧密相关。由表1可以看出:花蕾长度在2.5~4.1 cm时,花药长度在1.6~2.4 cm,在此期间的材料可以观察到花粉细胞减数分裂的各个时期;花蕾长度、花药长度与减数分裂进程相对应,随着花蕾和花药长度的增加,减数分裂也由间期进入到减数分裂Ⅰ时期和减数分裂Ⅱ时期;减数分裂期花药和其内的花粉母细胞、以及单核期花药和其内的花粉颜色基本一致,以此可为取样提供确切依据,同时也可以作为判断各分裂时期的依据。
2.2 花粉母细胞减数分裂过程的观察
本研究观察得出‘Cocossa’百合的花粉母细胞要经过减数分裂Ⅰ和减数分裂Ⅱ以及2次胞质分裂,最终形成四分体,其胞质分裂的方式为连续型。
2.2.1 减数分裂Ⅰ染色体的变化
处于花粉母细胞时期的是细胞紧密排列的多边形或近圆形(图1:A)。前期Ⅰ细线期:该时期细胞核增大,核内染色质浓缩形成细丝状染色体(图1:B)。偶线期:该时期细丝状染色体开始分散,并缩短增粗,但染色体仍为较长的细线,缠绕在一起不能区分(图1:C)。粗线期:该时期的染色体缩短变粗,配对后相互缠绕,难以区分(图1:D)。终变期:该时期染色体显著变粗变短, 并向核周边移动,配对后的染色体在核内均匀分散排列,
表1花粉母细胞减数分裂期花器官形态特征比较
Table1Floralorganmorphologycomparisonduringpollenmothercells
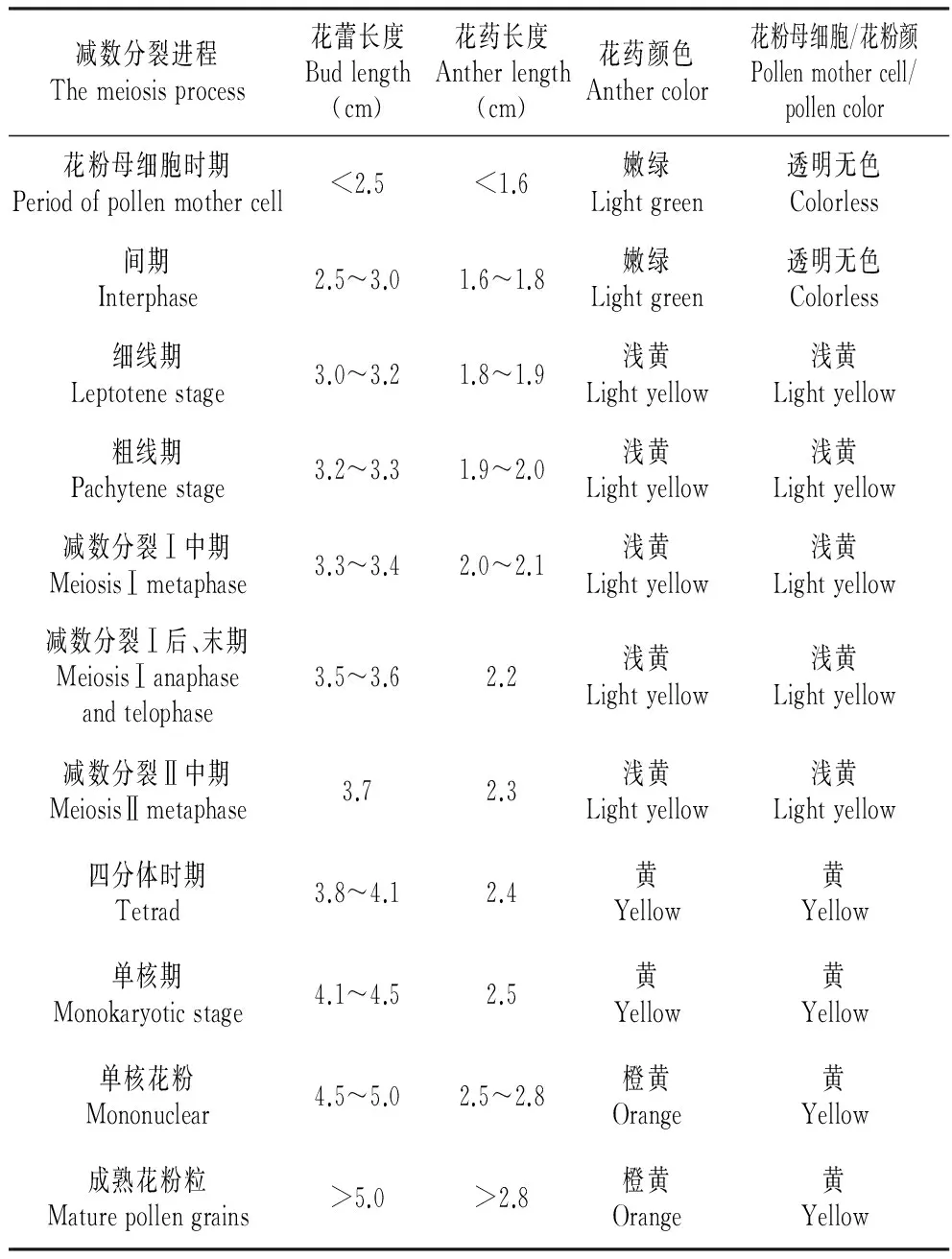
减数分裂进程Themeiosisprocess花蕾长度Budlength(cm)花药长度Antherlength(cm)花药颜色Anthercolor花粉母细胞/花粉颜Pollenmothercell/pollencolor花粉母细胞时期Periodofpollenmothercell<2.5<1.6嫩绿Lightgreen透明无色Colorless间期Interphase2.5~3.01.6~1.8嫩绿Lightgreen透明无色Colorless细线期Leptotenestage3.0~3.21.8~1.9浅黄Lightyellow浅黄Lightyellow粗线期Pachytenestage3.2~3.31.9~2.0浅黄Lightyellow浅黄Lightyellow减数分裂Ⅰ中期MeiosisⅠmetaphase3.3~3.42.0~2.1浅黄Lightyellow浅黄Lightyellow减数分裂Ⅰ后、末期MeiosisⅠanaphaseandtelophase3.5~3.62.2浅黄Lightyellow浅黄Lightyellow减数分裂Ⅱ中期MeiosisⅡmetaphase3.72.3浅黄Lightyellow浅黄Lightyellow四分体时期Tetrad3.8~4.12.4黄Yellow黄Yellow单核期Monokaryoticstage4.1~4.52.5黄Yellow黄Yellow单核花粉Mononuclear4.5~5.02.5~2.8橙黄Orange黄Yellow成熟花粉粒Maturepollengrains>5.0>2.8橙黄Orange黄Yellow
此期是对染色体计数的最好时期(图1:E)。中期Ⅰ:核膜和核仁消失,纺锤丝出现并形成纺锤体。染色体排列在赤道面上,但染色体重叠不利于对染色体形态进行观察(图1:F)。后期Ⅰ,两条同源染色体被纺锤丝牵引拉向两级,染色体随机分布在细胞的两极,染色体数目减半(图1:G)。在末期Ⅰ,到达细胞两极的染色体解螺旋,重新形成核膜,核仁重新出现,形成两个子核(图1:H)。在减数分裂Ⅰ,细胞进行第一次胞质均等分裂,产生新的胼胝质细胞壁,形成二分体(图1:I)。同时可以得出‘Cocossa’品种百合的胞质分裂方式为连续型。
2.2.2 减数分裂Ⅱ染色体的行为
前期Ⅱ:减数分裂Ⅰ结束后,DNA并不复制,经短暂的间期,染色质缩短变粗成为染色体(图1:J)。中期Ⅱ:核膜核仁再度消失,染色体分散排列在各自的赤道面上(图1:K)。后期Ⅱ:姊妹染色单体分离,由纺锤丝牵引均匀拉向两极(图1:L)。末期Ⅱ:到达两极的染色体浓缩成团,核膜核仁出现,此时进行第二次胞质分裂,各孢子形成胼胝质,而四个小孢子又被共同的胼胝质壁所包围,从而形成四分体,我们观察到百合四分体结构为“四面体”型(图1:M)或“十字”型(图1:N),至此完成减数分裂全过程。
此后,小孢子在胼胝质壁内继续发育,体积逐渐增大,形成花粉粒,最终胼胝质壁某处破裂,花粉粒从破裂处逐个释放(图1:O)。游离的花粉粒近似椭圆形,细胞核大且居中,细胞质浓厚,没有观察到明显的液泡,外壁网纹明显(图1:P)。
2.3花粉母细胞减数分裂过程中染色体异常现象的观察
本试验观察到OT百合‘Cocossa’花粉母细胞减数分裂的过程中,同一花蕾的花药内减数分裂过程大多数是同步的,而且在同一花药内大多数细胞分裂也具同步性。但在某些细胞中也会出现几个时期并存的现象如:前期Ⅰ终变期的分裂相中存在中期Ⅰ状态的染色体;后期Ⅰ的视野中,可观察到中期Ⅰ和末期Ⅰ并存的情况;在中期Ⅱ为主的分裂相中存在早期四分体等,最大跨度为2个分裂相。同时,我们观察到减数分裂中染色体和花粉粒不正常的情况,如出现不等二价体、染色体桥、微核、畸形花粉粒等。
2.3.1 染色体环
由于同源染色体存在缺失和重复区域的,因此我们在中期Ⅰ观察到有些染色体配对中出现‘染色体环’(图1:Q)。
2.3.2 染色体桥、滞后染色体和微核
后期Ⅰ细胞观察到染色体单桥现象(图1:R~S)。同时,在此时期,还观察到染色体双桥(图1:T)。在后期Ⅰ可以观察到1条或多条滞后染色体,有的位于赤道板上(图1:U)。这些滞后染色体随机分布形成微核(图1:V),我们推测由于滞后染色体形成的微核并不存在于细胞核之内,在随后的发育过程中丢失,会导致遗传物质减少,影响花粉粒的正常发育。
2.3.3 不均等分离
末期Ⅰ和末期Ⅱ‘Cocossa’百合中会出现同源染色体不均等分向两级的现象(图1:W),染色体不均等的分离会导致遗传物质不均等分配,从而导致配子不育。
2.3.4 其他异常花粉粒
小孢子尚未成熟继续在花粉囊中发育成为成熟花粉粒,花粉粒大小相差悬殊,观察到畸形花粉粒(图1:X)、连体花粉粒等(图1:Y)。
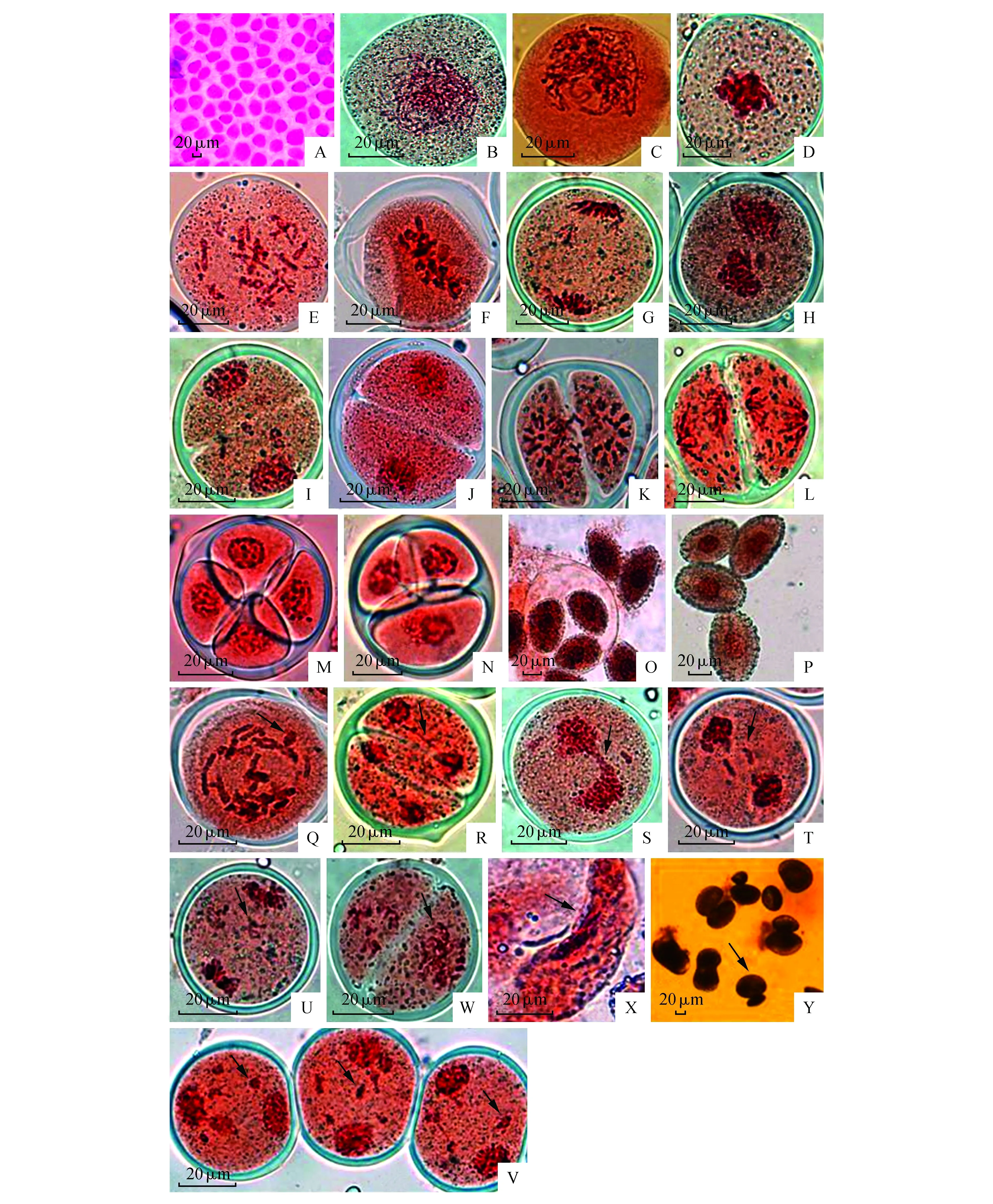
图1 花粉母细胞减数分裂过程及异常现象观察 A.花粉母细胞;B.前期Ⅰ细线期;C.前期Ⅰ偶线期;D.前期Ⅰ粗线期;E.前期Ⅰ终变期;F.中期Ⅰ;G.后期Ⅰ;H~I.末期Ⅰ;J.前期Ⅱ;K.中期Ⅱ;L.后期Ⅱ;M.末期Ⅱ四面体型四分体;N.末期Ⅱ十字型四分体;O.单核花粉粒释放;P.成熟花粉粒;Q.染色体环;R~S.染色体单桥;T.染色体双桥;U.滞后染色体;V.微核;W.染色体不均等分配;X.畸形花粉粒;Y.连体花粉粒Fig.1 Pollen mother cell meiosis and observe the anomaly A. Pollen mother cell;B. Prophase Ⅰ leptotene stage;C. Prophase Ⅰ zygotene stage;D. Prophase Ⅰ pachytene stage;E. Prophase Ⅰ diakinesis;F. Metaphase Ⅰ;G. Anaphase Ⅰ;H-I. Telophase Ⅰ;J. Prophase Ⅱ;K. Metaphase Ⅱ;L. AnaphaseⅡ;M. Tetrahedral tetrad;N. Cross tetrad;O. Mononuclear pollen grain;P. Mature pollen grains;Q. Chromosome ring;R-S. Chromosome bridges;T. Chromosome build;U. Lagging chromosomes;V. Micronuclei;W. Unequal division;X. Abnormal pollen;Y. Conjoined pollen
2.4 绒毡层的变化
绒毡层位于花药壁内层,对花粉的发育有重要的作用。花粉母细胞减数分裂接近完成时,正常发育过程中的绒毡层细胞便开始退化,在花粉发育的过程中继续解体。花粉发育后期,绒毡层细胞应解体,其产物为花粉合成DNA、RNA、蛋白质和淀粉提供原料[10]。本试验观察了近20张切片,每张切片中包含4个花粉囊,每张切片中均有绒毡层异常发育的现象,这种绒毡层异常现象是普遍存在的。我们观察到当花蕾长度为3.0~3.3 cm时,花粉母细胞处于减数第一次分裂的前期,绒毡层细胞和外面的药室内壁细胞区别不大,由大的液泡将原生质分割成原生质络,并将细胞核悬在中央,此时绒毡层细胞多数是单核的,少数为双核(图2:A);取材花蕾长度为3.3~3.9 cm时,花粉母细胞从中期Ⅰ发育到四分体时期,减数分裂接近完成时,绒毡层细胞形状仍没有明显变化,但细胞核向内侧靠拢,液泡逐渐分布于绒毡层的外侧,出现二核和多核绒毡层细胞(图2:B~C)。花蕾长度为4.4 cm,四分体解体释放出小孢子时,绒毡层细胞高度液泡化(图2:D)。当花蕾长度为5.3 cm,花粉粒发育成熟,绒毡层尚未解体,而缺少营养物质的花粉粒出现畸形或自溶等不正常发育(图2:E)。当取材花蕾长度为5.5 cm(图2:F~G),花粉粒自溶消失,绒毡层细胞开始解体。当取材花蕾长度为5.7 cm时,绒毡层完全解体,最终成熟的花药壁还包括一层表皮细胞、一层药室内壁细胞、两到多层中层细胞(图2:H)。
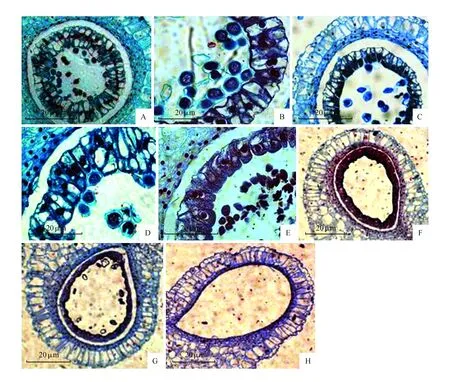
图2 ‘Cocossa’百合花粉减数分裂过程中绒毡层的变化 A.花蕾长度3.0~3.3 cm;B.花蕾长度3.3~3.9 cm;C.花蕾长度4.4 cm;D.花蕾长度5.0 cm;E.花蕾长度5.3 cm;F~G.花蕾长度5.5 cm;H.花蕾长度5.7 cmFig.2 ‘Cocossa’ lily pollen of tapetum in the process of meiosis A.Bud length 3.0-3.3 cm;B.Bud length 3.3-3.9 cm;C.Bud length 4.4 cm;D.Bud length 5.0 cm;E.Bud length 5.3 cm;F~G.Bud length 5.5 cm;H.Bud length 5.7 cm
从图2中我们可以发现,花粉母细胞减数分裂接近完成时,绒毡层细胞形状均没有明显变化,当小孢子从四分体释放形成花粉粒后,绒毡层细胞高度液泡化,当花粉已经成为成熟花粉粒时,绒毡层尚未解体。绒毡层降解和小孢子发育的不同步性,使得小孢子发育过程中,绒毡层不能为小孢子提供营养物质,以致最后小孢子败育。
3 讨论
OT百合‘Cocossa’减数分裂进程与花蕾、花药长度相关,可为取样提供确切依据,减数分裂期和单核期的花药和花粉颜色基本一致,可以非常直观的作为判断各分裂时期的依据,这与张彩霞[11~12]试验结果相一致。
本试验采用了染色体压片法并结合石蜡切片法对花粉母细胞的减数分裂过程和绒毡层的变化进行了系统的细胞学观察。结果显示在‘Cocossa’百合的同一花蕾以及同一花药中能观察到几个分裂相,这与张彩霞[13]和张伟[14]试验结果相似。‘Cocossa’的减数分裂符合二倍体植物的减数分裂过程,分裂类型属于连续型,在分裂过程中会出现染色体异常的现象,如在后期Ⅰ和末期Ⅰ出现了染色体桥、微核、不均等分离等。Errico认为减数分裂过程中的染色体桥和滞后染色体的出现,可能是臂内倒位造成的[15]。由倒位或易位形成的滞后染色体在末期会形成微核[16]。我们认为由于染色体不均等分离从而导致四分体时期遗传物质分配的不平衡甚至丢失,从而造成部分配子不育,这与Serclo[17]、雷家军等[18]的研究结果相似。
百合染色体基数较高且大,有丝分裂中易于导致染色体交错纠缠,增加断裂、缺失、易位、重复和倒位等结构变异的机率[19],本次试验材料为异源多倍体,减数分裂中非同源染色体异常配对,出现‘染色体环’。末期Ⅱ时形成的四分体类型为四面体型和十字型,在减数分裂的过程中出现遗传物质不均等分配、无核、多核小孢子等异常现象,形成畸形、无核、多核的异常小孢子,我们认为减数分裂过程中染色体异常是造成小孢子不育的原因之一。
绒毡层对花粉的正常发育也有决定作用,绒毡层为花粉的形成与发育提供所需的营养物质和水分。绒毡层发育异常,会直接导致花粉母细胞营养的缺失,使代谢旺盛的花粉母细胞不能得到所需营养,逐渐停止生长发育,最后导致花粉退化和自溶[20]。本试验观察到花粉母细胞减数分裂完成时,绒毡层细胞没有明显变化,当花粉成为成熟花粉粒时,绒毡层尚未解体,最终花粉粒因缺少所需营养物质而自溶。由此可以看出,‘Cocossa’是由于绒毡层延迟解体,使得小孢子在发育过程中,不能从绒毡层获得足够的营养物质,以致最后小孢子败育。绒毡层发育异常在是植物体小孢子败育重要的因素之一,如绒毡层不分化[21]、绒毡层提前或推迟解体[22]、绒毡层细胞质稀少高度液泡化等现象都可导致小孢子败育。Tsvetova等[9,23]对东方百合雄性不育的研究中发现,其败育是因为绒毡层提前解体导致其内含物流出形成周原质团导致败育。但是在本研究中,‘Cocossa’败育的现象与Tsvetova等研究中观察的现象不同,‘Cocossa’败育的原因是在第二次减数分裂末期,绒毡层延迟解体,导致小孢子在发育过程中不能获得足够的营养物质,一些花粉粒解体消失,从而导致雄性不育现象的发生。这种绒毡层异常的表现和黄邦全[24]、王伟[21]等人研究中发现绒毡层异常降解现象一致。
综上可以得出减数分裂异常和绒毡层延迟解体,是导致三倍体OT百合‘Cocossa’小孢子败育的主要因素。周树军[25]和袁国良[26]认为雄性不育的三倍体百合可以作为母本与二倍体或四倍体杂交,从而实现种质资源的渗入。因此探索三倍体百合小孢子发育过程,探究其有无产生2n雄配子的可能,可以为获得一定数量的可育雄配子提供理论依据,从而培育更多具有优良性状的百合品种。
1.崔光芬,王继华,贾文杰,等.OT百合新品种‘火凤凰’和‘金龙’[J].园艺学报,2013(40)10:2097-2098.
Cui G F,Wang J H,Jia W J,et al.OT Lily cultivars‘Huofenghuang’ and ‘Jinlong’[J].Acta Horticulturae Sinica,2013,(40)10:2097-2098.
2.张线线.利用GISH技术对百合(Lilium)组间杂交新品种染色体组的分析[D].杭州:浙江大学,2011.
Zhang X X.Study on genomic variation of new cultivars selected from distant hybridization inLiliumwith GISH[D].Hangzhou:Zhejiang University,2011.
3.Winton L,Einspahr D W.Tetraploid aspen production using unreduced triploid pollen[J].Forest Science,1970,16(2):180-182.
4.Harder M,Verhagen S,Winton L,et al.Tetraploid Aspen production using unreduced pollen from triploid males[J].Forest Science,1976,22(3):329-330.
5.梁国鲁,李育农,李晓林.中国苹果属植物小孢子减数分裂染色体系统研究[J].西南农业大学学报,1996,18(4)299-307.
Liang G L,Li Y N,Li X L.Evolutionary Study of The Chromosomes at Pollen Mother Cell Meiosis inMalus[J]. Journal of Southwest Agricultural University,1996,18(4)299-307.
6.王玉蛟,刘建鑫,于晓南.三倍体芍药品种花粉母细胞减数分裂行为观察与比较[J].植物研究,2016,36(3):341-347.
Wang Y J,Liu J X,Yu X N .Meiosis Observation and Comparison of Pollen Mother Cells of TriploidPaeonialactifloraCultivars[J].Bulletin of Botanical Research,2016,36(3):341-347.
7.张克中,赵祥云,梁励,等.王百合雄性不育突变体小孢子败育的细胞学研究[J].北京林业大学学报,2003,03:124-126.
Zhang K Z,Zhao X Y,Liang L,et al.Cytological studies on microspore abortion inLiliumregale male sterility mutant ‘White angel’[J].Journal of Beijing forestry university,2003,03:124-126.
8.李雪,陈丽梅,杜捷,等.兰州百合小孢子母细胞减数分裂异常现象的观察[J].西北植物学报,2003,10(23):1796-1799.
Li X,Chen L M,Du J,et al,Observations on abnormal meiosis of pollen mother cells inLiliumdavidiivar.unicolor[J].Acta Botanica Boreali-Occidentalia Sinica,2003,10(23):1796-1799.
9.郑洁,王有国,崔光芬,等.东方百合可育品种及其不育突变体花药形成的细胞学观察[J].西南农业学报,2015,03(28):1246-1250.
Zheng J,Wang Y G,Cui G F,et al,Cytological observation of anther formation ofLilium‘Oriental Hybrids’ normal line and male sterile mutant line[J].Southwest China Journal of Agricultural Sciences.2015,03(28):1246-1250.
10.胡适宜.被子植物胚胎学[M].北京:人民教育出版社,1982.
Hu S Y.Angiosperm embryology[M].Beijing:People’s education press,1982.
11.张彩霞,明军,刘春,等.百合花粉母细胞减数分裂异常现象观测分析[J].生物学通报,2010,45(5):45-47.
Zhang C X,Ming J,Liu C,et al.An abnormal meiosis of pollen mother cells observes in the Oriental hybrids lily[J].Bulletin of Biology,2010,45(5):45-47.
12.刘晓瑞.百合科植物花粉形态及花粉发育的研究Ⅱ[D].北京:首都师范大学,2008.
Liu X R.Study on the pollen morphology and pollen development of LiliaceaeⅡ[D].Beijing:Capital Normal University,2008.
13.张彩霞,明军,李博生.百合花粉母细胞减数分裂及其雄配子体发育观测[J].生物学通报,2009,44(10):53-56.
Zhang C X,Ming J,Li B S.The meiotic behavior of pollen mother cell and the formation of male gametophyte in the Oriental Hybrids Lily ‘Siberia’[J].Bulletin of Biology,2009,44(10):53-56.
14.张伟,张宁,张红亮.萝卜花粉母细胞减数分裂及其雄配子体发育[J].南京农业大学学报,2007,30(3):38-41.
Zhang W,Zhang N,Zhang H L.Pollen mother cell mitosis and male gametophyte development ofRaphanussativus[J].Journal of Nanjing Agricultural University,2007,30(3):38-41.
15.Angela E,Cammareri M,Conicella C.Meiotic nuclear restitution mechanisms in a triploid lily[J].Caryologia,2005,58(4):367-373.
16.许晓光,陈龙,彭建营.酸枣和赞皇大枣花粉母细胞减数分裂的观察[J].园艺学报,2009,36(8):1127-1133.
Xu X G,Chen L,Peng J Y.Meiotic observations on the pollen mother cells ofZiziphusspinosusHu.andZiziphusjujubeaMill ‘Zan huang Da zao’[J].Acta Horticulturae Sinica,2009,36(8):1127-1133.
17.Del Bosco S F,Tusa N,Conicella C.Microsporogenesis in a citrus interspecific tetraploid somatic hybrid and its fusion parents[J].Heredity,1999,83(4):373-377.
18.雷家军,梁印.百合三倍体种间杂种花粉母细胞减数分裂行为观察[J].吉林农业大学学报,2012,34(2):163-165.
Lei J J,Liang Y.Meiosis observation of pollen mother cells of triploid interspecific hybrids in Lily[J].Journal of Jilin Agricultural University,2012,34(2):163-165.
19.周朝鸿.三种百合属植物再生植株的染色体数量变异[J].林业科学研究,1997,10(6):663-667.
Zhou C H.Chromosomal numerical variations in regenerated plants of 3 species inLilium[J].Forest Research,1997,10(6):663-667.
20.Shivanna K R,Johri B M.The angiosperm pollen structure and function[M].Indianapolis:John Wiley & Sons Inc,1989:14-20.
21.王伟,谭敦炎,田允温,等.S351-1西瓜雄性不育的细胞学研究[J].西北植物学报,1998,18(3):361-365.
Wang W,Tan D Y,Tian Y W,et al.The Cytological study on watermelon male-sterility material S351-1[J].Acta botanica Boreali-Occidentalia Sinica,1998,18(3):361-365.
22.姚长兵,胡绍安,王春英,等.棉花细胞质雄性不育小孢子母细胞败育的细胞学研究[J].棉花学报,1994,6(S1):25-27.
Yao C B,Hu S A,Wang C Y,et al.Cytological studies on microspore mother cell abortion of cytoplasmic male sterile line of cotton[J].Acta Gossypii Sinica Cotton,1994,6(S1):25-27.
23.Tsvetova M I,Elkonin L A.Cytological investigation of male sterility in sorghum caused by a dominant mutation(Mstc) derived from tissue culture[J].Sexual Plant Reproduction,2003,16(1):43-49.
24.黄邦全,罗鹏,吴书惠.甘蓝型油菜雄性不育材料S90-8-7的获得及细胞学研究[J].湖北大学学报:自然科学版,2000,22(2):182-184.
Huang B Q,Luo P,Wu S H.Studies on male sterile line S90-8-7 derived from progenies of hybrids betweenBrassicanapusandOrychophragmusviolaceus[J].Journal of Hubei University:Natural Science Edition,2000,22(2):182-184.
25.周树军.三倍体百合的雌性可育机制及定向种质创新[C].//中国园艺学会.2012年园艺植物染色体倍性操作与遗传改良学术研讨会论文摘要集.重庆:中国园艺学会,2012:1.
Zhou S J.Female fertile mechanism and directional germplasm innovation of triploid lily[C].//A summary of the Symposium on chromosome ploidy manipulation and genetic improvement of horticultural plants in 2012.Chongqing:Chinese Society for Horticultural Science,2012:1.
26.袁国良.用三倍体作母本实现百合种质渗入育种[D].杭州:浙江大学,2014.
Yuan G L.Lily introgression breeding using triploids cultivars as maternal parents[D].Hangzhou:Zhejiang University,2014.
Beijing Collaborative innovation center for eco-environmental improvement with forestry and fruit(PXM2016_014207_000038),Beijing Municipal Education Commission;The Project of Construction of Innovative Teams and Teacher Career Development for Universities and Colleges Under Beijing Municipality(IDHT20150503),Beijing Municipal Education Commission
introduction:WU Jie(1990—),female,graduate student,Main research direction: Ornamental plant resources and breeding.
date:2016-09-14
CytologicalStudyonMicrosporeAbortioninOTHybridLilyCultivars‘Cocossa’
WU Jie ZHAO Yu-Ping HE Xiang-Feng WANG Wen-He*LENG Ping-Sheng ZHANG Ke-Zhong
(1.College of Landscape Archirecture,Beijing University of Agriculture,Beijing 102206;2.Beijing Collaborative innovation center for eco-enviromental improvement with forestry and fruit trees,Beijing 102206;3.Beijing Laboratory of Urban and Rural Ecological Environment,Beijing 102206)
We studied meiosis process and tepetum changes of OT hybrid lily cultivars ‘Cocossa’ using aceto-carmine dyeing method combined with grilled slices paraffin sections. When the bud grew to 2.5-4.5 cm, the pollen mother cell was in the period meiosis. There were some abnormalities appearances of meiosis: chromosome ring appeared in metaphase Ⅰ, chromosome bridges and lagging chromosomes ppeared in anaphaseⅠ, unequal division and micronuclei appeared in telophaseⅠand Ⅱ, it could also be observed 2 split-phase in the same plant, the same flower bud and even the same anther, that showed meiosis had not synchronized. During the telophaseⅡ and mononuclear pollen period, the tapetum delayed degradation, which led to multinuclear pollen, abnormal pollen and microspore infertility. The meiotic abnormalities and abnormal tapetum degradation could common cause the microspore abortion.
OT hybrid lily;meiosis;tapetum;microspore abortion
科技创新服务能力建设—协同创新中心—林果业生态环境功能提升协同创新中心(PXM2016_014207_000038),北京市教委;北京市属高等学校创新团队建设与教师职业发展计划(IDHT20150503),北京市教委
吴杰(1990—),女,硕士研究生。主要从事园林植物资源与育种方面的研究。
* 通信作者:E-mail:wwhals@163.com
2016-09-14
* Corresponding author:E-mail:wwhals@163.com
(1.北京农学院园林学院,北京 102206;2.北京林果业生态环境功能提升协同创新中心,北京 102206;3.城乡生态环境北京实验室,北京 102206)
Q949.71+8.23
A
10.7525/j.issn.1673-5102.2017.01.003
