Effect of glial cells on remyelination after spinal cord injury
2017-11-08JaifengwangXingkaiLiuRuiLiPingZhangZeChuChunliwangJuaruiLiuJunQiGuoyueLvGuangyiwangBinLiuYanLiYuanyiwang
Jai-feng wang, Xing-kai Liu, Rui Li, Ping Zhang, Ze Chu, Chun-li wang, Jua-rui Liu, Jun Qi, Guo-yue Lv, Guang-yi wang,Bin Liu, Yan Li, Yuan-yi wang
1 Department of Neurosurgery, First Hospital of Jilin University, Changchun, Jilin Province, China
2 Department of Hepatobiliary and Pancreas Surgery, First Hospital of Jilin University, Changchun, Jilin Province, China
3 Hand & Foot Surgery and Reparative & Reconstruction Surgery Center, Second Hospital of Jilin University, Changchun, Jilin Province, China
4 Department of Emergency, First Hospital of Jilin University, Changchun, Jilin Province, China
5 Department of Cardiology, First Hospital of Jilin University, Changchun, Jilin Province, China
6 Department of Surgery, School of Medicine, University of Louisville, Louisville, KY, USA
7 Department of Orthopedics, First Hospital of Jilin University, Changchun, Jilin Province, China
How to cite this article: Wang HF, Liu XK, Li R, Zhang P, Chu Z, Wang CL, Liu HR, Qi J, Lv GY, Wang GY, Liu B, Li Y, Wang YY (2017)Effect of glial cells on remyelination aer spinal cord injury. Neural Regen Res 12(10):1724-1732.
Funding: is work was supported by the National Natural Science Foundation of China, No. 81601957.
Effect of glial cells on remyelination after spinal cord injury
Jai-feng wang1, Xing-kai Liu2, Rui Li3, Ping Zhang2, Ze Chu4, Chun-li wang2, Jua-rui Liu2, Jun Qi2, Guo-yue Lv2, Guang-yi wang2,Bin Liu5,*,#, Yan Li6, Yuan-yi wang7,*,#
1 Department of Neurosurgery, First Hospital of Jilin University, Changchun, Jilin Province, China
2 Department of Hepatobiliary and Pancreas Surgery, First Hospital of Jilin University, Changchun, Jilin Province, China
3 Hand & Foot Surgery and Reparative & Reconstruction Surgery Center, Second Hospital of Jilin University, Changchun, Jilin Province, China
4 Department of Emergency, First Hospital of Jilin University, Changchun, Jilin Province, China
5 Department of Cardiology, First Hospital of Jilin University, Changchun, Jilin Province, China
6 Department of Surgery, School of Medicine, University of Louisville, Louisville, KY, USA
7 Department of Orthopedics, First Hospital of Jilin University, Changchun, Jilin Province, China
How to cite this article: Wang HF, Liu XK, Li R, Zhang P, Chu Z, Wang CL, Liu HR, Qi J, Lv GY, Wang GY, Liu B, Li Y, Wang YY (2017)Effect of glial cells on remyelination aer spinal cord injury. Neural Regen Res 12(10):1724-1732.
Remyelination plays a key role in functional recovery of axons aer spinal cord injury. Glial cells are the most abundant cells in the central nervous system. When spinal cord injury occurs, many glial cells at the lesion site are immediately activated, and different cells differentially affect inflammatory reactions aer injury. In this review, we aim to discuss the core role of oligodendrocyte precursor cells and crosstalk with the rest of glia and their subcategories in the remyelination process. Activated astrocytes influence proliferation, differentiation, and maturation of oligodendrocyte precursor cells, while activated microglia alter remyelination by regulating the inflammatory reaction aer spinal cord injury. Understanding the interaction between oligodendrocyte precursor cells and the rest of glia is necessary when designing a therapeutic plan of remyelination aer spinal cord injury.
nerve regeneration; spinal cord injury; remyelination; oligodendrocyte precursor cells; astrocytes;oligodendrocytes; microglia; glial scar; demyelination; myelin; central nervous system; neural regeneration
Introduction
Spinal cord injury (SCI) is common and involves widespread damage to the central nervous system (CNS). SCI often leads to severe neurological symptoms such as varying degree of paralysis, paresthesia, urinary obstruction, and other progressive neurological abnormalities. SCI also involves social loss: data on western countries show that governments spend $40,000–$180,000 for each patient depending on the site of injury. Patients lose their jobs and receive medical treatment, rehabilitation, and maintenance, and each patient costs the country millions of dollars (Ning et al., 2012). In the 1920s, SCI cases increased from 6.7 to 60 per million in some regions of China (Ning et al., 2012).
The pathological process of SCI can be divided into two stages: primary injury and secondary injury. Primary injury occurs immediately aer the initial injury, and its pathological processes include demyelination of the spinal cord and necrosis of neurons and axons (Yu et al., 2016). Secondary injury occurs throughout the disease, and its pathological processes include demyelination, axonal and neuronal necrosis, nervous tissue ischemia and edema, oxidative stress,inflammatory reaction, and glial scar formation (Balentine,1978; Kwo et al., 1989; Wrathall et al., 1996; Azbill et al., 1997;Ray et al., 2016). Among these pathological reactions, demyelination occurs immediately aer injury, and is induced by oligodendrocyte necrosis after mechanical damage. At the stage of secondary injury, because of extensive apoptosis and autophagy of oligodendrocytes, axons that have not been damaged or are slightly damaged become necrotic owing to demyelination (Almad et al., 2011).
Myelin can be regenerated. When demyelinating lesions occur, newly generated oligodendrocytes can repair or reconstruct damaged myelin. Regeneration of myelin, with oligodendrocyte generation as the main physiological process, can last up to three months aer SCI. A recent study found that most oligodendrocytes required for remyelination aer demyelination are derived from oligodendrocyte precursor cells (OPCs) and neural progenitor cells. OPCs can be labeled by neural/glial antigen 2 (NG2) or platelet-derived growth factor (PDGF) receptor alpha, and show very active proliferation in the CNS (Alizadeh et al., 2015).Previously, OPCs were discovered to have a role in repairing myelin (Hackett and Lee, 2016). Moreover, OPCs have been called the fourth glial cells, in addition to astrocytes,microglia, and oligodendrocytes. OPCs become mature oligodendrocytes through migration, proliferation, differentiation, and maturation, and subsequently repair injured myelin. Nevertheless, the amount of new myelin is unable to cover all exposed axons, and the remyelination rate cannot keep up with the speed of demyelination. A negative myelin balance increases the number of naked axons, and thereby results in disability, degeneration, and sensory and motor disorders in residual nerves. In the CNS, the interaction between various glial cells and neurons is consistently demonstrated. Further, many of the physiological and pathological responses are strongly associated with intercellular biological signaling pathways. Increasing evidence shows that cellular interactions play a significant role in demyelination and remyelination (Domingues et al., 2016). After central nerve injury, various glial cells directly or indirectly damage myelin. Simultaneously, these glial cells also affect myelin regeneration. Here, the aim of this review is to summarize latest research results and discuss the effect of glial cells on remyelination in nervous tissue aer SCI.
Myelin and Demyelination
Myelin is composed of cytoplasm and the membrane of oligodendrocytes and Schwann cells. Myelin wraps around axons forming a special sheath-like structure. In the nervous system, the resistance of myelin is high, which reduces the capacitance of ensheathed axons. Consequently, myelin provides the structural basis for saltatory conduction of nerve signals (Nave and Werner, 2014). Myelin also provides nutritional support for ensheathed axons (Li and Leung, 2015).Only CNS oligodendrocytes generate myelin. Moreover,oligodendrocytes are associated with nerve signal transduction. A previous study suggested that myelin damage leads to abnormal neurological behavior (Love, 2006). Demyelination occurs immediately aer SCI.e mechanism of demyelination remains unclear, but one likely reason is death of oligodendrocytes induced by various factors (Nave and Trapp, 2008).ese factors include tumor necrosis factor-alpha- and interleukin-1 beta-mediated inflammatory reactions, glucose–adenosine triphosphate-mediated cytotoxicity, edema, and various free radical-induced ischemia/reperfusion injury (Almad et al., 2011; Plemel et al., 2014). A previous study demonstrated that a single oligodendrocyte can be involved with 30–80 axons, with each connection wrapping into an internode (O’Rourke et al., 2014). Thus,accidental death of each oligodendrocyte can cause a series of demyelination (Chong et al., 2012; Young et al., 2013).Physiologically, there is a special signaling pathway between myelin and the axon, with one reason for demyelination being that the axon–oligodendrocyte signaling pathway is damaged after axonal injury (Alizadeh et al., 2015). In the absence of axonal nutritional support, oligodendrocyte degeneration rapidly occurs, resulting in demyelinating lesions(Lappe-Siee et al., 2003).
OPCs and Remyelination
OPCs are small cells of bipolar or tripolar structure, which can be found in the white and gray matter of the CNS.e number of OPCs is greater in the white matter than gray matter (Dawson et al., 2003; Dincman et al., 2012). As precursor cells, a new view of OPC outcome has recently been developed from in vivo and in vivo studies. Purified rat OPCs can dedifferentiate into neural stem cells and then differentiate into neurons, oligodendrocytes, and type I and II astrocytes (Kondo and Raff, 2000; Belachew et al., 2003;Nunes et al., 2003). In contrast, Tognatta et al. (2017) meticulously labeled differentiating OPCs in mice with 2′,3′-cyclic nucleotide 3′-phosphodiesterase (CNP)-Cre, but obtained insufficient evidence of OPC differentiation to neurons. OPCs can be specifically labeled by PDGF receptor alpha and NG2 proteoglycans (Tripathi and McTigue, 2007; Barnabe-Heider et al., 2010). OPCs can directly differentiate into oligodendrocytes without cell division (Hughes et al., 2013), but do not express NG2 aer differentiating into oligodendrocytes.
Nevertheless, the quality and integrity of regenerated myelin cannot meet demands owing to environmental change aer injury (Alizadeh et al., 2015). In the microenvironment after SCI, degenerative myelin secretes many inhibitory molecules. Simultaneously, the extracellular matrix, glial cell proliferation, and downregulation of nutrients and growthfactors affect remyelination (Meletis et al., 2008; Gauthier et al., 2013; Lukovic et al., 2015). The extracellular matrix can inhibit remyelination by blocking OPC migration (Siebert et al., 2011). Interleukin-beta limits OPC recruitment by activating the interleukin-1 receptor type 1 pathway(Kuroiwa et al., 2014).e glial scar produced by glial cells not only hinders OPC migration, but also results in a microenvironment that is not suitable for OPC proliferation.Degenerative myelin activates multiple microglial signaling pathways leading to release of inflammatory mediators (Sun et al., 2010). These molecules and cytokines inhibit axonal regeneration and destroy myelin integrity through the complement system (Chen et al., 2000).

Table 1 Factors known that regulate remyelination via different effects on OPCs
In summary, there are several reasons for lack of remyelination: (1) the remyelination process lacks the necessary growth factors for promoting formation of intact mature myelin from newborn oligodendrocytes; or (2) there is death of newborn OPCs as there are not enough biochemical factors to promote production of related cells and myelin.Consequently, the microenvironment at the injured site after SCI has an inhibitory effect on remyelination. In view of this, OPCs should be at the core of studies on remyelination,relieving inhibition, and promoting proliferation and differentiation of OPCs.
Recently, increasing research has focused on promoting remyelination by improving OPC migration, proliferation,differentiation, and maturation after SCI. Many drugs,hormones, and even treatments have been used clinically and are shown to be effective (Table 1). A previous study reported that as a hormone, progesterone improves OPC survival rate at the injury site by mitigating the inflammatory response and improving reactive gliosis after SCI(Huang et al., 2015). The Chinese herbal medicine, dried tangerine peel, can improve remyelination by increasing bone morphogenetic protein (BMP) 2.5 expression and elevating Ddx54 expression in cerebral ventricles, the subventricular zone, and corpus callosum (Tokunaga et al.,2016). Amiloride is a potassium-conserving diuretic that has been shown to promote remyelination by reducing the endoplasmic reticulum stress response and reducing OPC apoptosis (Kuroiwa et al., 2014). In addition to these drugs,there is evidence that physical therapy also has a role in promoting remyelination. Huang et al. (2015) reported that electroacupuncture promotes OPC proliferation, reduces OPC death, and improves remyelination. There have also been breakthroughs in promoting remyelination by overexpressing certain molecules in OPCs. Yao et al. (2017) reported that PGDF-AA-overexpressing OPC transplantation in rats induces remyelination. Myelin regulatory factor (MRF)overexpression was also reported to stimulate OPC differentiation (Xie et al., 2016). Although the mechanism of remyelination is not fully understood, there are numerous ways to promote remyelination. Most of these methods are supported by compelling evidence, but there is still considerable distance between these factors and clinical applications, and a need for continued innovation.
Astrocytes and Remyelination
Astrocytes are widely present in the CNS. They are the most abundant glial cells in white matter and gray matter,and have a crucial role in neurophysiology. A recent study demonstrated that two kinds of astrocytes in brain tissue: fibrous astrocytes in the white matter of the corpus callosum,and protoplasmic astrocytes in the gray matter (Ding, 2014).e primary function of astrocytes was initially thought to support and supply neurons, but nowadays there is plenty of evidence showing that astrocytes are strongly associated with microglia, oligodendrocytes, and other astrocytes in the nervous system. Astrocytes regulate neurotransmitters, participate in synaptogenesis, mediate the immune response, express extracellular matrix molecules, promote cell migration, and promote differentiation and maturation of the CNS (Walz, 1989; Westergaard et al., 1995; Sofroniew and Vinters, 2010; Clarke and Barres, 2013). Astrocytes are associated with many pathological CNS processes, including inflammation, ischemia, infection, and degeneration. Aer activation, changes in cell morphology, gene expression, and cell physiology are observed in astrocytes (Sofroniew and Vinters, 2010). A previous study confirmed that astrocytes directly affect proliferation and survival of the oligodendrocyte line (Li et al., 2016), demonstrating that oligodendrocytes are strongly associated with remyelination. Astrocytes are involved in regulating the balance between Schwann cells and oligodendrocyte remyelination, with oligodendrocyte remyelination only observed in areas where astrocytes are present. A recent study showed that testosterone promoted oligodendrocyte remyelination via astrocyte recruitment(Bielecki et al., 2016). Indeed, increasing evidence shows that astrocytes directly or indirectly affect remyelination by acting on OPCs or oligodendrocytes (Figure 1).
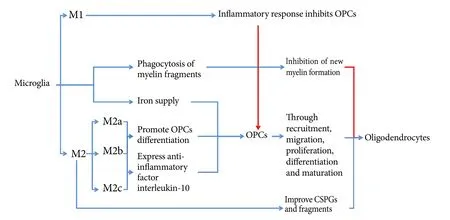
Figure 2 Microglia and remyelination.
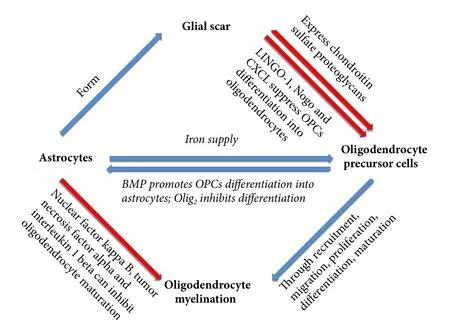
Figure 1 Astrocytes affect remyelination by affecting OPC differentiation and maturation or directly acting on oligodendrocytes.
OPCs and astrocytes are homologous during development, and OPCs can directly differentiate into astrocytes in vitro (Raff et al., 1983). Furthermore, an in vivo study found that immature astrocytes are present within NG2+cells aer SCI (Lytle et al., 2009). While another study confirmed that after SCI, OPCs that differentiate into oligodendrocytes are limited. Further, some OPCs (4–13%) do not differentiate into oligodendrocytes, and instead differentiate into astrocytes (Sozmen et al., 2016). OPC differentiation into astrocytes will affect remyelination. BMP and Olig2 may be involved in differentiation of OPCs into astrocytes. Regarding BMP, current understanding is that BMP4 increases aer SCI, with its potential source being reactive astrocytes aer injury (Wang et al., 2011). BMP4 contributes to OPC differentiation into astrocytes, although BMP4 antagonists have only a limited inhibitory effect on differentiation into astrocytes (Hampton et al., 2007). Olig2 may inhibit OPC differentiation into astrocytes: Olig2 overexpression reduces differentiation of neural stem cells into astrocytes in vitro(Fukuda et al., 2004). During development, a large number of Olig2 knockout OPCs differentiate into astrocytes instead of myelin (Zhu et al., 2012). Reticulon 4 receptor (NgR1) is a Nogo receptor that can suppress OPC differentiation into oligodendrocytes. Its antagonist promotes OPC differentiation into mature oligodendrocytes (Hampton et al., 2007;Sozmen et al., 2016). Molecules that promote OPC differentiation into astrocytes also include hyaluronan, janus kinase(JAK)-Stat1, and jagged-1 (Back et al., 2005; Zhang et al.,2009). In addition, inhibition of leucine rich repeat and Ig domain containing 1 (LINGO-1) promotes OPC differentiation into mature oligodendrocytes, and LINGO-1 inhibitors have been used for treatment of multiple sclerosis (Mi et al.,2013). In conclusion, OPC differentiation into astrocytes and oligodendrocytes ensures remyelination is a fluctuating process. Specifically, excessive OPC differentiation into astrocytes reduces the number of mature oligodendrocytes.Astrocytes have a significant inhibitory effect on remyelination and axonal regeneration.us, recovery of neurological function worsens aer SCI. Inhibition of astrocyte differentiation contributes to remyelination and ensures recovery of neurological function aer SCI.
Activated astrocytes lead to specific reactive gliosis. During this process, their morphology changes significantly and a large amount of intermediate filament proteins, mainly glial fibrillary acidic protein (GFAP) and nestin, are secreted to form the glial scar (Karimi-Abdolrezaee and Billakanti,2012). There are two sources of activated astrocytes after SCI: (1) ependymal cell GFAP-astrocytes; and (2) in situ activated GFAP+astrocytes.ey play different roles in glial scar formation (Meletis et al., 2008; Barnabe-Heider et al.,2010). Activated astrocytes are harmful to remyelination and involved in scar tissue formation, inhibition of OPC migration, survival and differentiation aer SCI, and even axonal regeneration (Wang et al., 2011). As a type of immunocyte in the CNS, astrocytes express many protein kinases, glycoproteins, and chondroitin sulfate proteoglycans aer activation.ese molecules induce inflammatory responses and the glial scar directly or indirectly causes severe damage to oligodendrocytes and neurons, chemically or physically (Silver and Miller, 2004). Glial scar formation limits the inflammatory reaction around the injury site, isolates damaged nerve tissue from normal tissue, and plays a supporting role in injured tissue. Simultaneously, the glial scar has a negative effect on remyelination and axonal regeneration. When axonal regeneration is inhibited, the link between axon and myelin is destroyed, and remyelination is not possible. Transplantation of OPCs and neural precursor cells into the injury site at the subacute stage contributes to axon myelination, but does not achieve a good outcome.is indicates that the internal environment aer injury around the glial scar has an inhibitory effect on remyelination or myelination of axons (Keirstead et al., 2005; Karimi-Abdolrezaee et al., 2006). Activated astrocytes secrete a variety of chondroitin sulfate proteoglycans,mainly consisting of neuroncan and brevican, and versican in the nervous system (Yamada et al., 1994).ey all have an inhibitory effect on remyelination and axonal regeneration(Dyck and Karimi-Abdolrezaee, 2015). Activated astrocytes affect OPC recruitment and maturation, and axonal ensheathment by secreting chondroitin sulfate proteoglycans,and finally inhibiting remyelination (Dyck and Karimi-Abdolrezaee, 2015). Chondroitin sulfate proteoglycans not only affect OPCs, but Karimi-Abdolezaee et al. (2010) found that chondroitin sulfate proteoglycans and the glial scar affect differentiation of neural precursor cells to oligodendrocytes.e glial scar is not only composed of astrocytes and microglia, and reactive activated OPCs are also involved in scar formation. OPCs also express chondroitin sulfate proteoglycans to inhibit axonal regeneration and repair myelin (Chen et al.,2002). Another inhibitory molecule secreted by astrocytes is hyaluronan, which is extensively found in the extracellular matrix and white matter of the CNS (Sherman et al., 2002).Hyaluronan can act on CD44 receptors of T cells and OPCs,and affect OPC maturation (Back et al., 2005; Lundgaard et al., 2014).
Reactive activated astrocytes aer SCI participate in glial scar formation. Changes in their own cell products and the microenvironment surrounding glial scars have a strong inhibitory effect on remyelination (Wang et al., 2015). Some inflammatory factors mitigate scar formation in reactive gliosis by inhibiting astrocyte activation, which may be a way to improve remyelination aer SCI. Wang et al. (2015)suggested that blocking the signaling pathway of platelet activating factor can reduce reactive gliosis and inhibit demyelination after SCI. Ishii et al. (2016) found that the RAS-related C3 botulinum substrate 1 (Rac1)–G1 to S phase transition 1 (GSPT1) signaling pathway is a new axis for regulating gliosis aer SCI.ese studies provide evidence for remyelination aer SCI.
Microglia and Remyelination
Microglia are macrophages present in the nervous system,and are involved in cellular immunity of the nervous system.Microglia are usually in a resting state, and in this state are in a “cruising” state to detect a pathological reaction at any time (Hanisch and Kettenmann, 2007). When a “crisis” arises, microglia can be immediately activated from the resting state, migrate to the injury site, and participate in formation of the outer layer of the glial scar to isolate damaged tissue from normal tissue (Davalos et al., 2005). Nevertheless,excessively activated microglia secrete large amounts of inflammatory factors, cytotoxic agents, and free radicals,thereby causing a severe inflammatory response, which undoubtedly inhibits remyelination. However, in recent years,more and more studies have focused on promoting the effect of microglia on remyelination. Microglia can be divided into different subtypes in the CNS, which play distinct roles in remyelination (Figure 2).
Microglia have an important effect on remyelination.With demyelinating lesions following SCI, some myelin fragments may remain outside residual axons. If these residual myelin fragments cannot be removed, they will have an impact on new myelin. Microglia are responsible for removal of fragments (Kotter et al., 2006; Neumann et al., 2009).Both in vivo and in vivo, these residual fragments can influence differentiation, maturation, and myelination of OPCs(Nave, 2010). A previous study reported that this microglial function is dependent on downstream activation of the DAP12 signaling pathway by triggering receptor expressed on myeloid cells 2 (TREM2) (Poliani et al., 2015). A residual amount of these fragments is associated with phagocytic function of microglia/macrophages. Moreover, this function is largely determined by the age of the organism. If the blood of young animals is injected into the body of older animals, remyelination of older animals is improved (Miron and Franklin, 2014). Astrocytes recruit microglia to the site of injury by expressing the chemokine, CXCL10, which enhances phagocytosis of myelin fragments. If astrocytes are removed from the culture medium, removal of myelin fragments can be affected, resulting in inhibition of proliferation and myelination of OPCs (Skripuletz et al., 2013). Receptors associated with microglial phagocytosis of myelin fragments include CR3, SRA, and Fc gamma. A previous study found that CR3 can reduce phagocytosis by activating or downregulating microglial phagocytosis to act on phosphorylated cofilin via the spleen tyrosine kinase (Syk) signaling pathway(Hadas et al., 2012). Simultaneously, CR3 and SRA interact to mediate phagocytosis following axonal injury (Makranz et al., 2004). Expression of galectin-3/MAC-2 can alter phagocytosis of microglia by modulating CR3 and SRA (Rotshenker et al., 2008).e TLR4 agonist, E6020, promotes repair of damaged myelin by stimulating microglia phagocytosis and myelinating cell recruitment (Church et al., 2017). It also blocks the TLR4 signaling pathway leading to delayed phagocytosis and altered expression of cytokines such as insulin-like growth factor-1, fibroblast growth factor-2, and interleukin-1 beta, which ultimately reduces remyelination aer SCI (Church et al., 2016).
Macrophages/microglia secrete a variety of cytokines,chemokines, and growth factors to affect remyelination aer SCI. Microglia are divided into two subtypes, namely,M1 cells involved in the inflammatory response and M2 cells with anti-inflammatory and repair effects (Kigerl et al.,2009). M1 cells are strongly associated with the inflammatory response and suppress remyelination. M2 cells are classified into three subtypes: M2a, M2b, and M2c (Gensel and Zhang, 2015). Kigerl et al. (2009) found that M2 microglia ameliorate chondroitin sulfate proteoglycan-induced axonal degeneration and reduce residual myelin fragments. M2 microglia gradually occupy a dominant activated microglial position at 3–10 days aer demyelination.is time window coincides with OPC recruitment and differentiation into mature oligodendrocytes at the site of injury (Miron et al.,2013). A further study verified that M2a and M2c microglia promote differentiation and maturation of oligodendrocytes by selectively removing M2 microglia (Miron and Franklin,2014).ese above studies confirm that M1 microglia inhibit remyelination aer SCI. M2a and M2c (suspected) microglia may promote remyelination by promoting recruitment,proliferation, differentiation, and maturation of OPCs aer SCI. Interleukin-10 is secreted by M2b microglia and an anti-inflammatory cytokine. Aer SCI, with activation of M2a cells, M2b cells reach a peak at 4–5 days aer injury. Another study demonstrated that M2b cells protect against axonal degeneration. Although it is not clear if M2b cells have a direct effect on remyelination, there is enough evidence to show that M2b and M2c cells promote spinal cord tissue repair by modulating cell proliferation (including OPCs) at the proliferative stage aer SCI (Gensel and Zhang, 2015). Bartus et al.(2014) have found that lentiviral introduction of the ChABC gene immediately aer SCI promotes a neuroprotective form of M2 microglia and increases storage of neurons and axons after 12 weeks of SCI. They also reported that this effect of ChABC may be produced by increasing expression of the anti-inflammatory factor, interleukin-10, and reducing the inflammatory factor, interleukin-12 beta (Didangelos et al.,2014). To date, increasing pathways have been shown to shiM1/M2 polarization.e amount of M1/M2 polarization is associated with age, with more M1 polarization detected in infarcted brain from older stroke models and more M2 labels found in younger ones (Suenaga et al., 2015). Also, many mediators (such as interleukin-4 and -13) can enhance M2 polarization (Wang et al., 2014; Roszer, 2015). Wang et al.(2017) reported that heterochromatin protein 1c (HP-1c)activates the 5′AMP-activated protein kinase (AMPK)-Nrf2 pathway to alter M1/M2 polarization and reduce the inflammatory reaction in stroke models. Cocoa polyphenolic extract is reported to shiM1/M2 polarization, in which M1 polarization is reduced and alternatively, M2 polarization induced (Dugo et al., 2017). Although quite a few pathways are related to M1/M2 polarization, and many molecules have shown their anti-inflammatory potential by reducing/inducing M1/M2 polarization, alteration of M1/M2 polarization aer SCI has yet to be fully understood.
Besides astrocytes, microglia are also associated with the iron supply chain in the nervous system. Increasing iron content in microglia increases the survival rate of co-cultured OPCs, verifying that microglia are a source of iron in OPCs (Zhang et al., 2006). Considering a similar role of astrocytes, microglia may improve iron protein content in both types of glial cells after SCI, improve iron supply in OPC–oligodendrocyte lines, and be helpful for remyelination aer injury.
Summary
With an increasing number of SCI patients, the study of demyelination/remyelination aer SCI has become increasingly significant. In addition to neurons, glial cells are resident cells in the CNS. Glial cells play supporting, nutritional, and immunological roles in the CNS. Simultaneously, glial cells are intimately associated with each other. Aer SCI, various signaling pathways are initiated, which can activate/injure glial cells and induce an inflammatory response, glial scar formation, neuronal injury, necrosis, and demyelination.In demyelinating lesions, OPCs in nerves replace lost oligodendrocytes and become new myelin via migration, proliferation, differentiation, and maturation. However, aer glial cell activation, the surrounding environment is changed and OPC myelination is affected by many factors. Astrocytes are the most abundant glial cells in the CNS.ey secrete chon-droitin sulfate proteoglycans after activation. Astrocytes also induce glial scar formation, which has a large effect on remyelination. Microglia as major immune cells of the CNS initiate an inflammatory response aer injury. Inflammatory cytokines expressed in microglia affect remyelination. M2 microglia promote OPC proliferation, differentiation, and maturation. Taken together, controlling reactive activation of glial cells after SCI to improve remyelination is an important approach to treat injured spinal cord and promote recovery of neurological function.
Author contributions:HFW conceived the manuscript. XKL, PZ, ZC,HRL and JQ collected data. YL searched bibliography. RL organized figures and table. YYW draed and revised the paper. BL wrote the paper.GYL and GYW revised the paper. All authors approved the final version of the paper.
Conflicts of interest: None declared.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by ienticate.
Peer review:Externally peer reviewed.
Open access statement:is is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under identical terms.
Open peer reviewer:Na Lin, Kunming Medical University, Basic Medical Sciences, China.
Alizadeh A, Dyck SM, Karimi-Abdolrezaee S (2015) Myelin damage and repair in pathologic CNS: challenges and prospects. Front Mol Neurosci 8:35.
Almad A, Sahinkaya FR, McTigue DM (2011) Oligodendrocyte fate aer spinal cord injury. Neurotherapeutics 8:262-273.
Azbill RD, Mu X, Bruce-Keller AJ, Mattson MP, Springer JE (1997)Impaired mitochondrial function, oxidative stress and altered antioxidant enzyme activities following traumatic spinal cord injury.Brain Res 765:283-290.
Back SA, Tuohy TM, Chen H, Wallingford N, Craig A, Struve J, Luo NL,Banine F, Liu Y, Chang A, Trapp BD, Bebo BF, Jr., Rao MS, Sherman LS (2005) Hyaluronan accumulates in demyelinated lesions and inhibits oligodendrocyte progenitor maturation. Nat Med 11:966-972.
Badaracco ME, Siri MV, Pasquini JM (2010) Oligodendrogenesis: the role of iron. BioFactors (Oxford, England) 36:98-102.
Balentine JD (1978) Pathology of experimental spinal cord trauma.I. The necrotic lesion as a function of vascular injury. Lab Invest 39:236-253.
Bansal R, Pfeiffer SE (1997) FGF-2 converts mature oligodendrocytes to a novel phenotype. J Neurosci Res 50:215-228.
Barnabe-Heider F, Goritz C, Sabelstrom H, Takebayashi H, Pfrieger FW, Meletis K, Frisen J (2010) Origin of new glial cells in intact and injured adult spinal cord. Cell Stem Cell 7:470-482.
Barres BA, Raff MC (1994) Control of oligodendrocyte number in the developing rat optic nerve. Neuron 12:935-942.
Barres BA, Burne JF, Holtmann B,oenen H, Sendtner M, Raff MC(1996) Ciliary neurotrophic factor enhances the rate of oligodendrocyte generation. Mol Cell Neurosci 8:146-156.
Bartus K, James ND, Didangelos A, Bosch KD, Verhaagen J, Yanez-Munoz RJ, Rogers JH, Schneider BL, Muir EM, Bradbury EJ(2014) Large-scale chondroitin sulfate proteoglycan digestion with chondroitinase gene therapy leads to reduced pathology and modulates macrophage phenotype following spinal cord contusion injury.J Neurosci 34:4822-4836.
Belachew S, Chittajallu R, Aguirre AA, Yuan X, Kirby M, Anderson S,Gallo V (2003) Postnatal NG2 proteoglycan-expressing progenitor cells are intrinsically multipotent and generate functional neurons. J Cell Biol 161:169-186.
Bielecki B, Mattern C, Ghoumari AM, Javaid S, Smietanka K, Abi Ghanem C, Mhaouty-Kodja S, Ghandour MS, Baulieu EE, Franklin RJM,Schumacher M, Traiffort E (2016) Unexpected central role of the androgen receptor in the spontaneous regeneration of myelin. Proc Natl Acad Sci U S A 113:14829-14834.
Brambilla R, Morton PD, Ashbaugh JJ, Karmally S, Lambertsen KL,Bethea JR (2014) Astrocytes play a key role in EAE pathophysiology by orchestrating in the CNS the inflammatory response of resident and peripheral immune cells and by suppressing remyelination. Glia 62:452-467.
Chen MS, Huber AB, van der Haar ME, Frank M, Schnell L, Spillmann AA, Christ F, Schwab ME (2000) Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature 403:434-439.
Chen ZJ, Negra M, Levine A, Ughrin Y, Levine JM (2002) Oligodendrocyte precursor cells: Reactive cells that inhibit axon growth and regeneration. J Neurocytol 31:481-495.
Chong SY, Rosenberg SS, Fancy SP, Zhao C, Shen YA, Hahn AT, Mc-Gee AW, Xu X, Zheng B, Zhang LI, Rowitch DH, Franklin RJ, Lu QR, Chan JR (2012) Neurite outgrowth inhibitor Nogo-A establishes spatial segregation and extent of oligodendrocyte myelination. Proc Natl Acad Sci U S A 109:1299-1304.
Church JS, Kigerl KA, Lerch JK, Popovich PG, McTigue DM (2016)TLR4 deficiency impairs oligodendrocyte formation in the injured spinal cord. J Neurosci 36:6352-6364.
Church JS, Milich LM, Lerch JK, Popovich PG, McTigue DM (2017)E6020, a synthetic TLR4 agonist, accelerates myelin debris clearance,Schwann cell infiltration, and remyelination in the rat spinal cord.Glia 65:883-899.
Clarke LE, Barres BA (2013) Emerging roles of astrocytes in neural circuit development. Nat Rev Neurosci 14:311-321.
Davalos D, Grutzendler J, Yang G, Kim JV, Zuo Y, Jung S, Littman DR,Dustin ML, Gan WB (2005) ATP mediates rapid microglial response to local brain injury in vivo. Nat Neurosci 8:752-758.
Dawson MR, Polito A, Levine JM, Reynolds R (2003) NG2-expressing glial progenitor cells: an abundant and widespread population of cycling cells in the adult rat CNS. Mol Cell Neurosci 24:476-488.
Deng Y, Xie D, Fang M, Zhu G, Chen C, Zeng H, Lu J, Charanjit K(2014) Astrocyte-derived proinflammatory cytokines induce hypomyelination in the periventricular white matter in the hypoxic neonatal brain. PLoS One 9:e87420.
Didangelos A, Iberl M, Vinsland E, Bartus K, Bradbury EJ (2014) Regulation of IL-10 by chondroitinase ABC promotes a distinct immune response following spinal cord injury. J Neurosci 34:16424-16432.
Dincman TA, Beare JE, Ohri SS, Whittemore SR (2012) Isolation of cortical mouse oligodendrocyte precursor cells. J Neurosci Methods 209:219-226.
Domingues HS, Portugal CC, Socodato R, Relvas JB (2016) Oligodendrocyte, astrocyte, and microglia crosstalk in myelin development,damage, and repair. Front Neurol 4:71.
Dugo L, Belluomo MG, Fanali C, Russo M, Cacciola F, Maccarrone M,Sardanelli AM (2017) Effect of Cocoa polyphenolic extract on macrophage polarization from proinflammatory M1 to anti-inflammatory M2 state. Oxid Med Cell Longev 2017:6293740.
Dyck SM, Karimi-Abdolrezaee S (2015) Chondroitin sulfate proteoglycans: Key modulators in the developing and pathologic central nervous system. Exp Neurol 269:169-187.
Franklin RJ, Ffrench-Constant C (2008) Remyelination in the CNS:from biology to therapy. Nat Rev Neurosci 9:839-855.
Fukuda S, Kondo T, Takebayashi H, Taga T (2004) Negative regulatory effect of an oligodendrocytic bHLH factor OLIG2 on the astrocytic differentiation pathway. Cell Death Differ 11:196-202.
Gauthier MK, Kosciuczyk K, Tapley L, Karimi-Abdolrezaee S (2013)Dysregulation of the neuregulin-1-ErbB network modulates endogenous oligodendrocyte differentiation and preservation aer spinal cord injury. Eur J Neurosci 38:2693-2715.
Gensel JC, Zhang B (2015) Macrophage activation and its role in repair and pathology aer spinal cord injury. Brain Res 1619:1-11.
Goddard DR, Berry M, Butt AM (1999) In vivo actions of fibroblast growth factor-2 and insulin-like growth factor-I on oligodendrocyte development and myelination in the central nervous system. J Neurosci Res 57:74-85.
Hackett AR, Lee DH, Dawood A, Rodriguez M, Funk L, Tsoulfas P, Lee JK (2016) STAT3 and SOCS3 regulate NG2 cell proliferation and differentiation aer contusive spinal cord injury. Neurobiol Dis 89:10-22.
Hadas S, Spira M, Hanisch UK, Reichert F, Rotshenker S (2012) Complement receptor-3 negatively regulates the phagocytosis of degenerated myelin through tyrosine kinase Syk and cofilin. J Neuroinflammation 9:166.
Hampton DW, Asher RA, Kondo T, Steeves JD, Ramer MS, Fawcett JW(2007) A potential role for bone morphogenetic protein signalling in glial cell fate determination following adult central nervous system injury in vivo. Eur J Neurosci 26:3024-3035.
Hanisch UK, Kettenmann H (2007) Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci 10:1387-1394.
Hesp ZC, Goldstein EZ, Miranda CJ, Kaspar BK, McTigue DM (2015)Chronic oligodendrogenesis and remyelination aer spinal cord injury in mice and rats. J Neurosci 35:1274-1290.
Hill RA, Patel KD, Medved J, Reiss AM, Nishiyama A (2013) NG2 cells in white matter but not gray matter proliferate in response to PDGF. J Neurosci 33:14558-14566.
Huang S, Tang C, Sun S, Cao W, Qi W, Xu J, Huang J, Lu W, Liu Q,Gong B, Zhang Y, Jiang J (2015) Protective effect of electroacupuncture on neural myelin sheaths is mediated via promotion of oligodendrocyte proliferation and inhibition of oligodendrocyte death after compressed spinal cord injury. Mol Neurobiol 52:1870-1881.
Hughes EG, Kang SH, Fukaya M, Bergles DE (2013) Oligodendrocyte progenitors balance growth with self-repulsion to achieve homeostasis in the adult brain. Nat Neurosci 16:668-676.
Ishibashi T, Lee PR, Baba H, Fields RD (2009) Leukemia inhibitory factor regulates the timing of oligodendrocyte development and myelination in the postnatal optic nerve. J Neurosci Res 87:3343-3355.
Ishii T, Ueyama T, Shigyo M, Kohta M, Kondoh T, Kuboyama T, Uebi T, Hamada T, Gutmann DH, Aiba A, Kohmura E, Tohda C, Saito N(2016) A novel Rac1-GSPT1 signaling pathway controls astrogliosis following central nervous system injury. J Biol Chem 292:1240-1250.
Ji B, Li M, Wu WT, Yick LW, Lee X, Shao Z, Wang J, So KF, McCoy JM,Pepinsky RB, Mi S, Relton JK (2006) LINGO-1 antagonist promotes functional recovery and axonal sprouting aer spinal cord injury. Mol Cell Neurosci 33:311-320.
Karimi-Abdolrezaee S, Billakanti R (2012) Reactive astrogliosis after spinal cord injury-beneficial and detrimental effects. Mol Neurobiol 46:251-264.
Karimi-Abdolrezaee S, Eekharpour E, Wang J, Morshead CM, Fehlings MG (2006) Delayed transplantation of adult neural precursor cells promotes remyelination and functional neurological recovery after spinal cord injury. J Neurosci 26:3377-3389.
Karimi-Abdolrezaee S, Eekharpour E, Wang J, Schut D, Fehlings MG(2010) Synergistic effects of transplanted adult neural stem/progenitor cells, chondroitinase, and growth factors promote functional repair and plasticity of the chronically injured spinal cord. J Neurosci 30:1657-1676.
Keirstead HS, Nistor G, Bernal G, Totoiu M, Cloutier F, Sharp K, Steward O (2005) Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion aer spinal cord injury. J Neurosci 25:4694-4705.
Kigerl KA, Gensel JC, Ankeny DP, Alexander JK, Donnelly DJ, Popovich PG (2009) Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J Neurosci 29:13435-13444.
Kim HM, Hwang DH, Choi JY, Park CH, Suh-Kim H, Kim SU, Kim BG(2011) Differential and cooperative actions of Olig1 and Olig2 transcription factors on immature proliferating cells aer contusive spinal cord injury. Glia 59:1094-1106.
Kondo T, Raff M (2000) Oligodendrocyte precursor cells reprogrammed to become multipotential CNS stem cells. Science 289:1754-1757.
Kotter MR, Li WW, Zhao C, Franklin RJ (2006) Myelin impairs CNS remyelination by inhibiting oligodendrocyte precursor cell differentiation. J Neurosci 26:328-332.
Kuroiwa M, Watanabe M, Katoh H, Suyama K, Matsuyama D, Imai T,Mochida J (2014) Effect of amiloride on endoplasmic reticulum stress response in the injured spinal cord of rats. Eur J Neurosci 40:3120-3127.
Kwo S, Young W, Decrescito V (1989) Spinal cord sodium, potassium,calcium, and water concentration changes in rats aer graded contusion injury. J Neurotrauma 6:13-24.
Lappe-Siefke C, Goebbels S, Gravel M, Nicksch E, Lee J, Braun PE,Griffiths IR, Nave KA (2003) Disruption of Cnp1 uncouples oligodendroglial functions in axonal support and myelination. Nat Genet 33:366-374.
Li J, Zhang L, Chu Y, Namaka M, Deng B, Kong J, Bi X (2016) Astrocytes in oligodendrocyte lineage development and white matter pathology. Front Cell Neurosci 10:119.
Li N, Leung GK (2015) Oligodendrocyte precursor cells in spinal cord injury: a review and update. Biomed Res Int 2015:235195.
Love S (2006) Demyelinating diseases. J Clin Pathol 59:1151-1159.
Lukovic D, Stojkovic M, Moreno-Manzano V, Jendelova P, Sykova E,Bhattacharya SS, Erceg S (2015) Concise review: reactive astrocytes and stem cells in spinal cord injury: good guys or bad guys? Stem Cells 33:1036-1041.
Lundgaard I, Osorio MJ, Kress BT, Sanggaard S, Nedergaard M (2014)White matter astrocytes in health and disease. Neurosci 276:161-173.
Lytle JM, Chittajallu R, Wrathall JR, Gallo V (2009) NG2 cell response in the CNP-EGFP mouse after contusive spinal cord injury. Glia 57:270-285.
Makranz C, Cohen G, Baron A, Levidor L, Kodama T, Reichert F,Rotshenker S (2004) Phosphatidylinositol 3-kinase, phosphoinositide-specific phospholipase-Cgamma and protein kinase-C signal myelin phagocytosis mediated by complement receptor-3 alone and combined with scavenger receptor-AI/II in macrophages. Neurobiol Dis 15:279-286.
Mayer M, Bhakoo K, Noble M (1994) Ciliary neurotrophic factor and leukemia inhibitory factor promote the generation, maturation and survival of oligodendrocytes in vitro. Development 120:143-153.
Meletis K, Barnabe-Heider F, Carlen M, Evergren E, Tomilin N,Shupliakov O, Frisen J (2008) Spinal cord injury reveals multilineage differentiation of ependymal cells. PLoS Biol 6:e182.
Mi S, Pepinsky RB, Cadavid D (2013) Blocking LINGO-1 as a therapy to promote CNS repair: from concept to the clinic. CNS Drugs 27:493-503.
Miron VE, Franklin RJ (2014) Macrophages and CNS remyelination. J Neurochem 130:165-171.
Miron VE, Boyd A, Zhao JW, Yuen TJ, Ruckh JM, Shadrach JL, van Wijngaarden P, Wagers AJ, Williams A, Franklin RJ, ffrench-Constant C (2013) M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat Neurosci 16:1211-1218.
Moore CS, Cui QL, Warsi NM, Durafourt BA, Zorko N, Owen DR,Antel JP, Bar-Or A (2015) Direct and indirect effects of immune and central nervous system-resident cells on human oligodendrocyte progenitor cell differentiation. J Immunol 194:761-772.
Nave KA (2010) Myelination and the trophic support of long axons.Nat Rev Neurosci. 11:275-283.
Nave KA, Trapp BD (2008) Axon-glial signaling and the glial support of axon function. Annu Rev Neurosci 31:535-561.
Nave KA, Werner HB (2014) Myelination of the nervous system:mechanisms and functions. Annu Rev Cell Dev Biol 30:503-533.
Neumann H, Kotter MR, Franklin RJM (2009) Debris clearance by microglia: an essential link between degeneration and regeneration.Brain 132:288-295.
Ning GZ, Wu Q, Li YL, Feng SQ (2012) Epidemiology of traumatic spinal cord injury in Asia: a systematic review. J Spinal Cord Med 35:229-239.
Nunes MC, Roy NS, Keyoung HM, Goodman RR, McKhann G, 2nd,Jiang L, Kang J, Nedergaard M, Goldman SA (2003) Identification and isolation of multipotential neural progenitor cells from the subcortical white matter of the adult human brain. Nat Med 9:439-447.
O’Rourke M, Gasperini R, Young KM (2014) Adult myelination: wrapping up neuronal plasticity. Neural Regen Res 9:1261-1264.
Ossola B, Zhao C, Compston A, Pluchino S, Franklin RJ, Spillantini MG (2016) Neuronal expression of pathological tau accelerates oligodendrocyte progenitor cell differentiation. Glia 64:457-471.
Plemel JR, Keough MB, Duncan GJ, Sparling JS, Yong VW, Stys PK,Tetzlaff W (2014) Remyelination aer spinal cord injury: is it a target for repair? Prog Neurobiol 117:54-72.
Poliani PL, Wang Y, Fontana E, Robinette ML, Yamanishi Y, Gilfillan S, Colonna M (2015) TREM2 sustains microglial expansion during aging and response to demyelination. J Clin Invest 125:2161-2170.
Raasch J, Zeller N, van Loo G, Merkler D, Mildner A, Erny D, Knobeloch K-P, Bethea JR, Waisman A, Knust M, Del Turco D, Deller T,Blank T, Priller J, Brück W, Pasparakis M, Prinz M (2011) IκB kinase 2 determines oligodendrocyte loss by non-cell-autonomous activation of NF-κB in the central nervous system. Brain 134:1184-1198.
Raff MC, Miller RH, Noble M (1983) A glial progenitor cell that develops in vitro into an astrocyte or an oligodendrocyte depending on culture medium. Nature 303:390-396.
Ray SK, Samntaray S, Banik NL (2016) Future directions for using estrogen receptor agonists in the treatment of acute and chronic spinal cord injury. Neural Regen Res 11:1418-1419.
Roszer T (2015) Understanding the mysterious M2 macrophage through activation markers and effector mechanisms. Mediators Inflamm 2015:816460.
Rotshenker S, Reichert F, Gitik M, Haklai R, Elad-Sfadia G, Kloog Y(2008) Galectin-3/MAC-2, Ras and PI3K activate complement receptor-3 and scavenger receptor-AI/II mediated myelin phagocytosis in microglia. Glia 56:1607-1613.
Salgado-Ceballos H, Guizar-Sahagun G, Feria-Velasco A, Grijalva I,Espitia L, Ibarra A, Madrazo I (1998) Spontaneous long-term remyelination aer traumatic spinal cord injury in rats. Brain Res 782:126-135.
Schwab ME, Strittmatter SM (2014) Nogo limits neural plasticity and recovery from injury. Curr Opin Neurobiol 27:53-60.
Sherman LS, Struve JN, Rangwala R, Wallingford NM, Tuohy TM,Kuntz Ct (2002) Hyaluronate-based extracellular matrix: keeping glia in their place. Glia 38:93-102.
Siebert JR, Stelzner DJ, Osterhout DJ (2011) Chondroitinase treatment following spinal contusion injury increases migration of oligodendrocyte progenitor cells. Exp Neurol 231:19-29.
Silver J, Miller JH (2004) Regeneration beyond the glial scar. Nat Rev Neurosci 5:146-156.
Skripuletz T, Hackstette D, Bauer K, Gudi V, Pul R, Voss E, Berger K, Kipp M, Baumgartner W, Stangel M (2013) Astrocytes regulate myelin clearance through recruitment of microglia during cuprizone-induced demyelination. Brain 136:147-167.
Sofroniew MV, Vinters HV (2010) Astrocytes: biology and pathology.Acta Neuropathol 119:7-35.
Sozmen EG, Rosenzweig S, Llorente IL, DiTullio DJ, Machnicki M,Vinters HV, Havton LA, Giger RJ, Hinman JD, Carmichael ST (2016)Nogo receptor blockade overcomes remyelination failure aer white matter stroke and stimulates functional recovery in aged mice. Proc Natl Acad Sci U S A 113:E8453-8462.
Suenaga J, Hu X, Pu H, Shi Y, Hassan SH, Xu M, Leak RK, Stetler RA,Gao Y, Chen J (2015) White matter injury and microglia/macrophage polarization are strongly linked with age-related long-term deficits in neurological function aer stroke. Exp Neurol 272:109-119.
Sun X, Wang X, Chen T, Li T, Cao K, Lu A, Chen Y, Sun D, Luo J, Fan J, Young W, Ren Y (2010) Myelin activates FAK/Akt/NF-kappaB pathways and provokes CR3-dependent inflammatory response in murine system. PLoS One 5:e9380.
Todorich B, Pasquini JM, Garcia CI, Paez PM, Connor JR (2009) Oligodendrocytes and myelination: the role of iron. Glia 57:467-478.
Tognatta R, Sun W, Goebbels S, Nave KA, Nishiyama A, Schoch S,Dimou L, Dietrich D (2017) Transient Cnp expression by early progenitors causes Cre-Lox-based reporter lines to map profoundly different fates. Glia 65:342-359.
Tokunaga H, Seiwa C, Yoshioka N, Mizoguchi K, Yamamoto M, Asou H, Aiso S (2016) An extract of chinpi, the dried peel of the citrus fruit unshiu, enhances axonal remyelination via promoting the proliferation of oligodendrocyte progenitor cells. Evid Based Complement Alternat Med 2016:8692698.
Tripathi A, Parikh ZS, Vora P, Frost EE, Pillai PP (2016) pERK1/2 peripheral recruitment and filopodia protrusion augment oligodendrocyte progenitor cell migration: combined effects of PDGF-A and fibronectin. Cell Mol Neurobiol 37:183-194.
Tripathi R, McTigue DM (2007) Prominent oligodendrocyte genesis along the border of spinal contusion lesions. Glia 55:698-711.
Walz W (1989) Role of glial cells in the regulation of the brain ion microenvironment. Prog Neurobiol 33:309-333.
Wang N, Liang H, Zen K (2014) Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front Immunol 5:614.
Wang Y, Cheng X, He Q, Zheng Y, Kim DH, Whittemore SR, Cao QL(2011) Astrocytes from the contused spinal cord inhibit oligodendrocyte differentiation of adult oligodendrocyte precursor cells by increasing the expression of bone morphogenetic proteins. J Neurosci 31:6053-6058.
Wang Y, Gao Z, Zhang Y, Feng SQ, Liu Y, Shields LB, Zhao YZ, Zhu Q, Gozal D, Shields CB, Cai J (2015) Attenuated reactive gliosis and enhanced functional recovery following spinal cord injury in null mutant mice of platelet-activating factor receptor. Mol Neurobiol 53:3448-3461.
Wang Y, Huang Y, Xu Y, Ruan W, Wang H, Zhang Y, Saavedra JM,Zhang L, Huang Z, Pang T (2017) A dual AMPK/Nrf2 activator reduces brain inflammation aer stroke by enhancing microglia M2 polarization. Antioxid Redox Signal doi: 10.1089/ars.2017.7003.
Wegener A, Deboux C, Bachelin C, Frah M, Kerninon C, Seilhean D, Weider M, Wegner M, Nait-Oumesmar B (2015) Gain of Olig2 function in oligodendrocyte progenitors promotes remyelination.Brain 138:120-135.
Westergaard N, Sonnewald U, Schousboe A (1995) Metabolic trafficking between neurons and astrocytes: the glutamate/glutamine cycle revisited. Dev Neurosci 17:203-211.
Wrathall JR, Teng YD, Choiniere D (1996) Amelioration of functional deficits from spinal cord trauma with systemically administered NBQX, an antagonist of non-N-methyl-D-aspartate receptors. Exp Neurol 137:119-126.
Xie XM, Shi LL, Shen L, Wang R, Qi Q, Wang QY, Zhang LJ, Lu HZ,Hu JG (2016) Co-transplantation of MRF-overexpressing oligodendrocyte precursor cells and Schwann cells promotes recovery in rat aer spinal cord injury. Neurobiol Dis 94:196-204.
Yamada H, Watanabe K, Shimonaka M, Yamaguchi Y (1994) Molecular cloning of brevican, a novel brain proteoglycan of the aggrecan/versican family. J Biol Chem 269:10119-10126.
Yao ZF, Wang Y, Lin YH, Wu Y, Zhu AY, Wang R, Shen L, Xi J, Qi Q,Jiang ZQ, Lu HZ, Hu JG (2017) Transplantation of PDGF-AA-overexpressing oligodendrocyte precursor cells promotes recovery in rat following spinal cord injury. Front Cell Neurosci 11:79.
Young KM, Psachoulia K, Tripathi RB, Dunn SJ, Cossell L, Attwell D, Tohyama K, Richardson WD (2013) Oligodendrocyte dynamics in the healthy adult CNS: evidence for myelin remodeling. Neuron 77:873-885.
Yu YQ, Hu NC, Duan JA, Li DP, Liu C (2016) Neuroprotective effects of sufentanil preconditioning on spinal cord injury in mouse models.Zhongguo Zuzhi Gongcheng Yanjiu 20:5966-5972.
Zawadzka M, Rivers LE, Fancy SP, Zhao C, Tripathi R, Jamen F, Young K, Goncharevich A, Pohl H, Rizzi M, Rowitch DH, Kessaris N, Suter U, Richardson WD, Franklin RJ (2010) CNS-resident glial progenitor/stem cells produce Schwann cells as well as oligodendrocytes during repair of CNS demyelination. Cell Stem Cell 6:578-590.
Zhang X, Surguladze N, Slagle-Webb B, Cozzi A, Connor JR (2006)Cellular iron status influences the functional relationship between microglia and oligodendrocytes. Glia 54:795-804.
Zhang Y, Argaw AT, Gurfein BT, Zameer A, Snyder BJ, Ge C, Lu QR,Rowitch DH, Raine CS, Brosnan CF, John GR (2009) Notch1 signaling plays a role in regulating precursor differentiation during CNS remyelination. Proc Natl Acad Sci U S A 106:19162-19167.
Zhu X, Zuo H, Maher BJ, Serwanski DR, LoTurco JJ, Lu QR, Nishiyama A (2012) Olig2-dependent developmental fate switch of NG2 cells. Development 139:2299-2307.
*Correspondence to:
Yuan-yi Wang, M.D. or Bin Liu, M.D.,tedwangyy@foxmail.com or 1466802342@qq.com,359382923@qq.com.
orcid:
0000-0002-6163-5144
(Yuan-yi Wang)
0000-0007-7182-8644
(Bin Liu)
10.4103/1673-5374.217354
Accepted: 2017-09-14
Copyedited by James R, Frenchman B, Wang J, Li CH, Qiu Y, Song LP,Zhao M
杂志排行
中国神经再生研究(英文版)的其它文章
- Matrix bound vesicles and miRNA cargoes are bioactive factors within extracellular matrix bioscaffolds
- Diffusion tensor tractography studies on mechanisms of recovery of injured fornix
- Using 3D bioprinting to produce mini-brain
- Beta secretase activity in peripheral nerve regeneration
- Embracing oligodendrocyte diversity in the context of perinatal injury
- On the road towards the global analysis of human synapses