水盐梯度对闽江河口湿地土壤有机碳组分的影响
2017-11-07刘兴土陈晓旋陈优阳牟晓杰万斯昂
王 纯,刘兴土*,仝 川,陈晓旋,陈优阳,牟晓杰,万斯昂
水盐梯度对闽江河口湿地土壤有机碳组分的影响
王 纯1,刘兴土1*,仝 川2,3,陈晓旋2,3,陈优阳2,3,牟晓杰1,万斯昂1
(1.中国科学院东北地理与农业生态研究所,湿地生态与环境重点实验室,吉林长春130120;2.福建师范大学地理科学学院,福建福州350007;3.福建师范大学亚热带湿地研究中心,福建福州350007)
为了揭示水盐梯度对河口湿地土壤有机碳组分的影响,对闽江河口不同淹水环境和盐度下短叶茳芏()湿地土壤活性有机碳含量进行了测定与分析.结果表明,无论半咸水湿地还是淡水湿地,土壤微生物生物量碳(MBC)含量均随淹水频率增加而增加,增幅分别为67.8%和38.8%.半咸水湿地高低潮滩的土壤MBC含量均低于淡水湿地,高低潮滩降幅分别为52.9%和43.1%.半咸水湿地高低潮滩土壤可溶性有机碳(DOC)含量均高于淡水湿地,增幅分别为56.7%和105.6%.2种湿地土壤易氧化有机碳(EOC)含量均随淹水频率增加而降低,半咸水湿地高低潮滩间降幅18.0%,淡水湿地降幅50.1%.半咸水湿地高低潮滩土壤EOC含量均高于淡水湿地,增幅分别为20.2%和97.4%.微生物熵以及DOC和EOC占SOC的比值分别为0.42%~1.76%、0.39%~0.85% 和20.14%~36.49%.微生物熵随盐度增加而降低,土壤DOC和EOC的分配比例随盐度的增加而增加.相对于淹水环境变化,土壤TN含量和电导率对SOC及其活性组分含量影响的贡献更大.土壤DOC、EOC与SOC显著正相关,土壤MBC与SOC、EOC和DOC均呈负相关,暗示底物的有效性和土壤MBC周转速率是影响土壤微生物活性和碳库积累的重要因子.淹水频率增加提高了土壤微生物的数量,但土壤微生物对淹水环境有一定的适应机制.盐度增加可提高土壤DOC、EOC含量,但降低土壤MBC含量.土壤氮含量和盐度是影响闽江河口湿地生态系统土壤碳库演变的重要限制性参数.
盐度;淹水环境;土壤有机碳;活性组分;闽江河口
土壤活性有机碳是指土壤中稳定性差、易分解矿化、周转速率快的碳素[1-2].其中土壤微生物生物量碳(MBC)、可溶性有机碳(DOC)和易氧化有机碳(EOC)是其主要的表征指标[3].虽然其占土壤总有机碳的比例很小,但它直接参与土壤营养元素生物化学转化过程,反应了土壤养分循环和土壤质量[4].相对于背景含量较高的土壤总有机碳,土壤活性有机碳对外界环境变化的响应更敏感[5],已成为预测土壤质量和土壤碳库演变的重要指标[6].
河口湿地位于陆地和海洋生态系统的交错区,是对全球变化和人类活动响应敏感的生态系统[7].盐度为入海河口区一个常见的环境胁迫因子,可通过改变土壤理化性质和微生物群落结构和功能,影响滨海河口湿地碳源汇功能[8].特别是在全球气候变暖、海平面上升背景下,含盐量高的海水入侵加上周期性的潮汐涨落,对滨海湿地土壤碳截获能力产生深刻影响.目前有关盐度对河口湿地生态系统影响的研究已引起了国内外学者的广泛关注,包括盐度变化影响土壤有机质的矿化途径[9]、温室气体的产生与排放[10-13]、微生物群落结构与功能[8,14]及生态系统生产力[15]等.河口区由于河流和潮汐作用引起的水文波动对湿地生态系统的影响也受到广泛关注,包括水文变化影响沼泽湿地沉积[16-17]、营养元素特征[18-21]、温室气体排放[22-23]、土壤碳库组分[24]及微生物群落结构[25]、湿地植物生长[26-27]及枯落物分解[28]等.同时考虑盐度和水文两个滨海河口湿地最为典型的影响因子对湿地生态系统的影响也逐步受到关注,并已取得了一些成果,包括盐度和水文协同变化影响温室气体排放[29]、生态系统呼吸和净生产力[30]、湿地植物生长[31]、枯落物分解和营养物质动态[32]等.但对于同时探讨盐度和水文双因子对湿地土壤碳库特征,特别是对环境干扰敏感的土壤活性有机碳组分的综合影响,这方面认识仍存在一定局限.本文以我国东南沿海典型的感潮河口—闽江河口为研究区,并沿盐度梯度选择闽江入海口的半咸水湿地(鳝鱼滩湿地)和溯流而上的淡水湿地(塔礁洲湿地)2个典型的河口湿地,同时在每个湿地选择2个潮水淹水环境差异显著的高低潮滩作为研究样地.旨在探讨不同盐度和水文条件下河口湿地土壤活性有机碳组分的空间分布特征;明确盐度和水文条件变化对河口湿地土壤活性有机碳组分空间分布影响的大小.
1 材料与方法
1.1 研究区概况
研究区位于闽江河口区,闽江河口区拥有闽江流域最大的天然河口湿地群(119°5′36″~ 119°41′5″E,25°50′43″~26°9′42″N),总面积约为467.6km2.该区气候暖热湿润,年平均气温为19.7℃,年降水日数为153d,年降水量约为1 346mm,降水集中在3~9月,表现为双峰型,峰值分别出现在6月和8月,前者为梅雨期,后者为台风期[33].潮汐属正规半日潮,5~12月潮水盐度均值为4.2‰[34].闽江入海口的鳝鱼滩湿地是闽江河口湿地群中面积最大的潮汐湿地,土壤以滨海盐土为主[35].上游方向的塔礁洲湿地则为感潮区淡水湿地,土壤电导率一般< 0.5mS/cm[13].短叶茳芏()是这2个河口湿地的主要土著优势植物群落.本研究分别在鳝鱼滩和塔礁洲短叶茳芏湿地沿高程选择一个典型样带(高低潮滩)开展实验(图1).
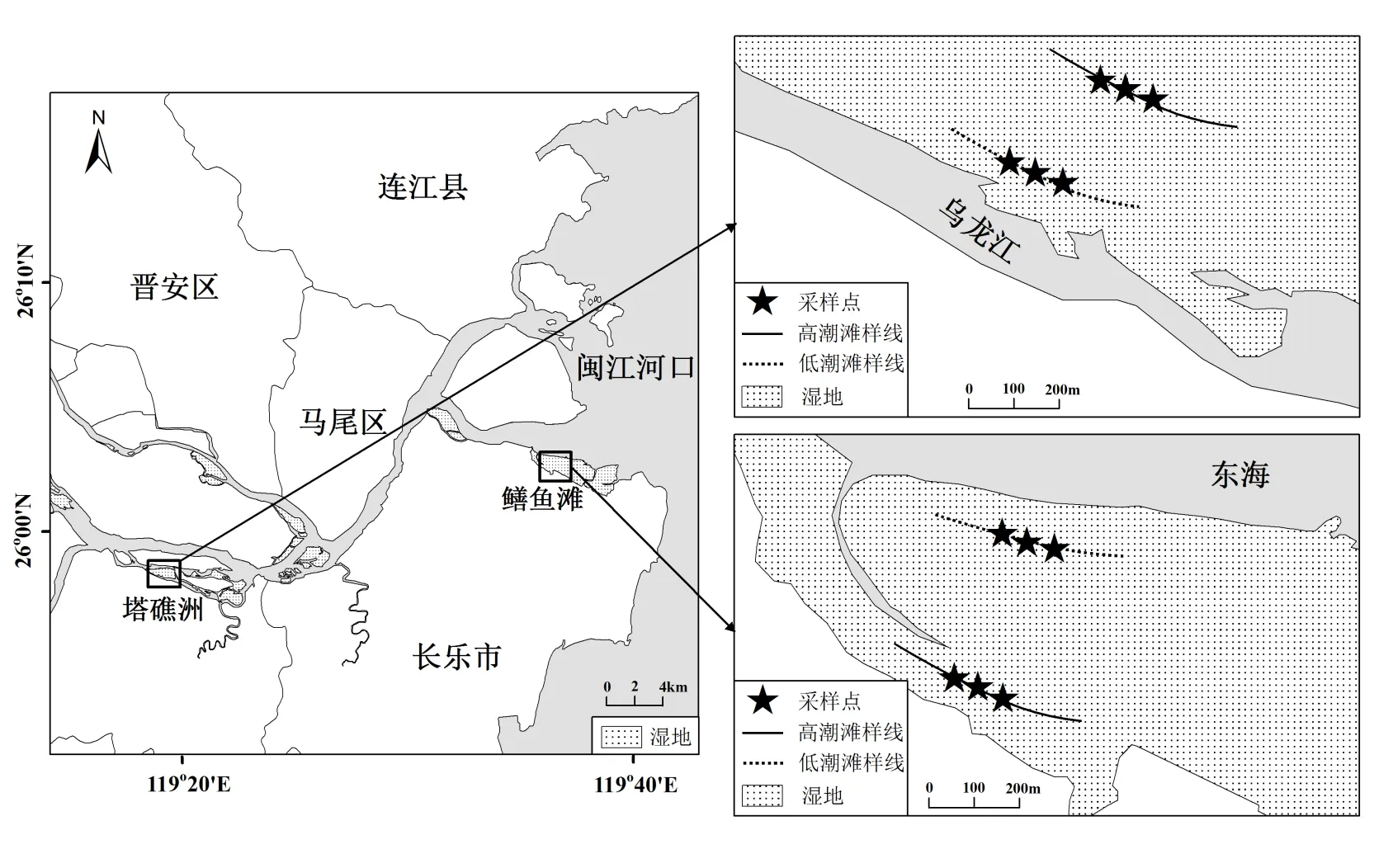
图1 研究区和采样点示意
1.2 土样采集与处理
由于6~10月是闽江河口区台风的多发季节[36],且该区从10月开始进入枯水期.河流径流减少叠加台风诱发的风暴潮,导致10月海水上溯现象十分严重,河口湿地土壤碳库特征对盐度和水文变化的响应尤为典型.故本研究于2016年10月在落潮后时段采集土壤样品.分别在上述2个湿地短叶茳芏群落典型地段,沿高程先确定淹水频率低的高潮滩和淹水频率高的低潮滩样线,然后沿海岸线或河岸线平行方向在高潮滩和低潮滩样线分别随机布置3个采样点(样点间距约5m),设为3个重复.取样时先去除样地土壤表层凋落物,然后用直径5.5cm不锈钢取土器钻取0~10cm深度的土柱.将采集的土样装入相应样品采集袋,置于有冰袋的便携式保鲜盒中运回实验室.回实验室后,先测量土壤电导率、含水率和容重.然后将采集的每个土样挑除根系和砂砾,混合均匀后分成2份,一份过10目筛,4℃冷藏,在48h之内测定土壤MBC和DOC含量;另一份自然风干,过100目筛,用于测定土壤pH值、土壤全碳含量(TC)、全氮(TN)含量及EOC含量.
1.3 土壤理化性质和活性碳组分测定与计算
土壤电导率采用2265FS便携式电导(Spectrum Technologies Inc,USA)测定,用环刀法结合烘干法测定土壤含水率和容重.土壤pH值采用IQ150便携式pH仪(IQ Scientific Instruments, USA)测定,土壤TC和TN含量用碳氮元素分析仪(vario MAX, Germany)测定.
土壤MBC含量采用氯仿熏蒸法--0.5mol/L K2SO4浸提法.浸提液用TOC-VCPH总有机碳分析仪(Shimadzu, Japan)测定,土壤MBC含量为熏蒸与未熏蒸土壤有机碳含量的差值除以浸提系数,本研究取值0.38[37].
土壤DOC含量采用0.5mol/L K2SO4浸提法.称10g过10目筛的鲜土样品,加入40mL 0.5mol/L K2SO4,密封振荡30min(250r/min),离心20min(4000r/min).浸提液用TOC-VCPH总有机碳分析仪(Shimadzu, Japan)直接测定.
土壤EOC采用K2MnO4氧化-比色法测定.称1g 过100目筛的干土样品,加入25mL 333mmol/L K2MnO4,密封振荡1h(120r/min),离心5min(4000r/min).取上清液用去离子水稀释250倍,用分光光度计在波长为565nm处进行比色.根据K2MnO4的消耗量计算土壤EOC含量,每减少1mmol/L的K2MnO4代表碳减少9mg[38].
同时,计算土壤活性有机碳占土壤总有机碳的比例.由于闽江河口湿地土壤总碳主要以有机碳为主(平均占比约93%)[25],因此,本研究拟定土壤总碳(TC)等同于土壤总有机碳(SOC).即计算土壤MBC/SOC、DOC/SOC和 EOC/SOC的比值大小,以明确土壤各活性有机碳组分在SOC中的分配比例,其中土壤MBC/SOC为微生物熵.
1.4 数据处理与分析
采用EXCEL 2010软件计算平均值和标准误.采用origin 8.0绘图软件作图.采用SPSS 17.0统计软件进行数据统计分析.首先,检验所有数据是否符合正态分布和方差齐性,当未通过检验时,将所有原始数据进行数据转换,直到数据符合条件后,进行方差分析.采用独立样本检验比较同一湿地不同潮滩或者不同湿地相同潮滩土壤活性有机碳的差异以及土壤理化性质的差异.采用Pearson相关分析和逐步线性回归方法分析土壤有机碳组分与土壤理化性质的关系以及土壤有机碳组分之间的关系.
2 结果与分析
2.1 水盐梯度下土壤活性有机碳含量比较
2种湿地土壤MBC含量为74.67~220.05mg/ kg,同一湿地高低潮滩间土壤MBC含量均表现为低潮滩大于高潮滩(>0.05),即土壤MBC含量随淹水频率增加而增加,半咸水湿地高低潮滩间增幅67.8%,淡水湿地增幅38.8%.淡水湿地高低潮滩土壤MBC含量均高于半咸水湿地(>0.05),即土壤MBC含量随盐度增加而降低,不同湿地高潮滩的降幅为52.9%,低潮滩的降幅为43.1% (表1).2种湿地土壤DOC含量为68.95~ 141.78mg/kg,2种湿地高低潮滩土壤DOC含量差异均不显著(>0.05).半咸水湿地高低潮滩土壤DOC含量均显著高于淡水湿地(<0.05),即土壤DOC含量随盐度增加而增加,不同湿地高潮滩的增幅为56.7%,低潮滩的增幅为105.6%(表1).2种湿地土壤EOC含量为2.50~6.03g/kg,2种湿地土壤EOC含量均表现为低潮滩低于高潮滩(> 0.05),即土壤EOC含量随淹水频率增加而降低,半咸水湿地高低潮滩间降幅18.0%,淡水湿地降幅50.1%.半咸水湿地高低潮滩土壤EOC含量均高于淡水湿地,即土壤EOC含量随盐度增加而增加,不同湿地高潮滩的增幅为20.2%,低潮滩的增幅为97.4%(表1).
2.2 水盐梯度下土壤活性有机碳占土壤总有机碳的比例
2种湿地土壤微生物熵为0.42%~1.76%,塔礁洲湿地低潮滩土壤微生物熵显著高于高潮滩,也显著高于鳝鱼滩湿地低潮滩(<0.05)(表2). 2种湿地土壤DOC/SOC的比值为0.39%~0.85%,2种湿地高低潮滩土壤DOC含量的分配比例均无显著差异(>0.05),鳝鱼滩湿地高低潮滩土壤DOC含量的分配比例均显著高于塔礁洲湿地(<0.05) (表2). 2种湿地土壤EOC/SOC的比值为20.14~ 36.49%,2种湿地高低潮滩土壤EOC含量的分配比例均无显著差异(>0.05),鳝鱼滩湿地高潮滩土壤EOC含量的分配比例显著高于塔礁洲湿地(<0.05)(表2).
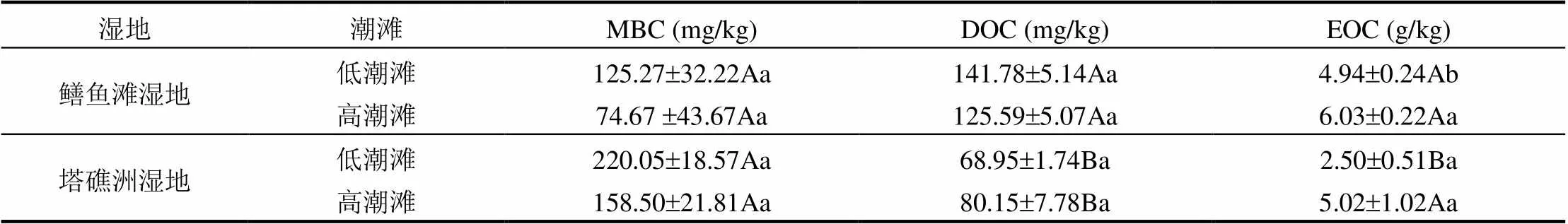
表1 不同湿地土壤MBC、DOC和EOC含量
注:同列小写字母表示同一湿地高低潮滩之间的差异,大写字母表示不同湿地相同潮滩之间的差异,显著性<0.05.
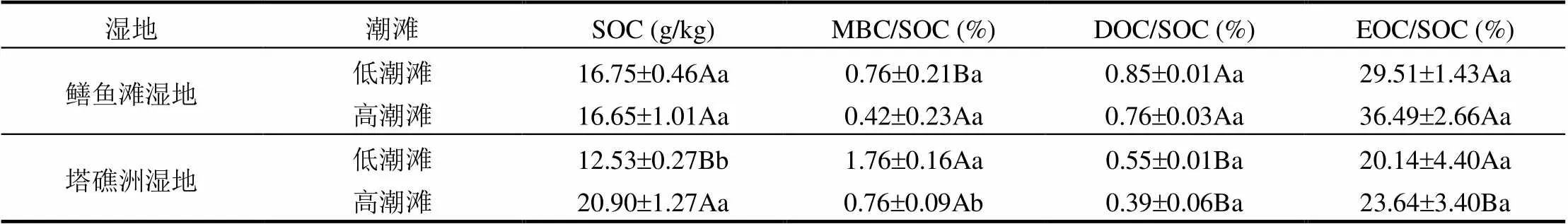
表2 土壤活性有机碳占SOC的比例
注:同列小写字母表示同一湿地高低潮滩之间的差异,大写字母表示不同湿地相同潮滩之间的差异,显著性<0.05.
2.3 水盐梯度下土壤理化性质差异
鳝鱼滩半咸水湿地高低潮滩土壤电导率均显著高于塔礁洲淡水湿地(<0.05),塔礁洲湿地低潮滩土壤电导率显著高于高潮滩(<0.05)(表3).土壤含水率和容重在2种湿地高低潮滩间均无显著差异(>0.05)(表3). 2种湿地高低潮滩土壤pH值差异均不显著(>0.05),鳝鱼滩湿地高潮滩土壤pH值显著高于塔礁洲湿地(<0.05)(表3).土壤TN和TC含量在2种湿地高低潮滩间具有相同的趋势,均表现为塔礁洲湿地高潮滩显著高于低潮滩(<0.05),鳝鱼滩湿地低潮滩显著高于塔礁洲湿地(<0.05)(表3).
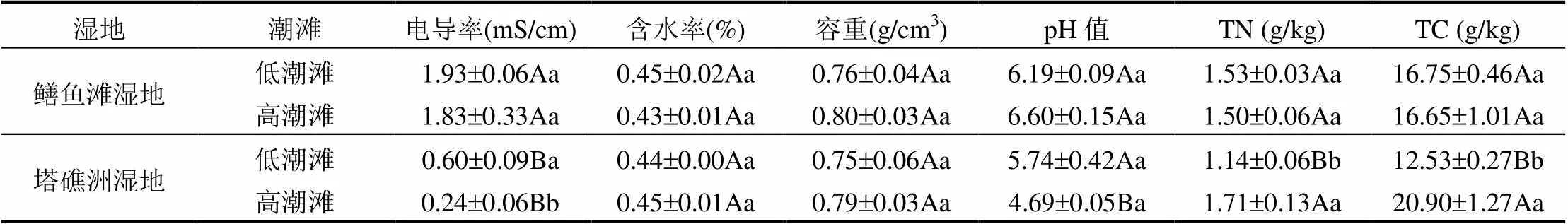
表3 土壤基本理化性质
注:同列小写字母表示同一湿地高低潮滩之间的差异,大写字母表示不同湿地相同潮滩之间的差异,显著性<0.05.
2.4 土壤有机碳组分与土壤理化性质之间的关系
相关分析(表4)表明, SOC含量与土壤EOC含量和C:N显著正相关(<0.05),与土壤TN含量极显著正相关(<0.01).土壤MBC含量与DOC含量显著负相关(<0.05).土壤DOC含量与电导率极显著正相关(<0.01),与土壤pH显著正相关(<0.05).土壤EOC含量与TN含量极显著正相关(<0.01).微生物熵与MBC含量极显著正相关(<0.01),与土壤DOC、EOC和TN含量呈显著或极显著负相关(<0.05和<0.01).土壤DOC/SOC与DOC含量、电导率和pH极显著正相关(<0.01).土壤EOC/SOC与DOC、EOC含量和电导率显著或极显著正相关(<0.05和<0.01),与土壤MBC含量显著负相关(<0.05).
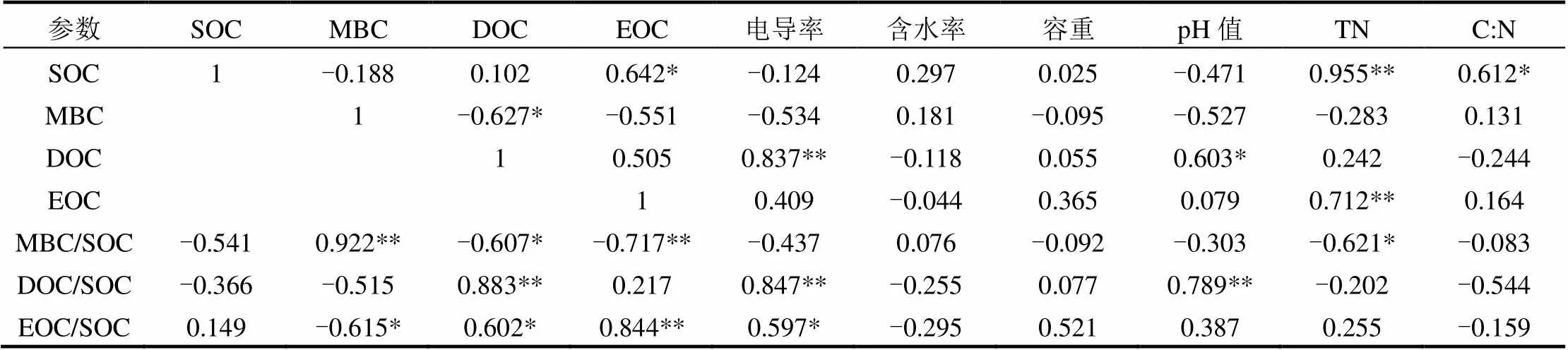
表4 土壤活性有机碳与土壤基本理化性质的相关分析
注:* 表示显著相关< 0.05,** 表示极显著相关< 0.01.

表5 土壤活性有机碳与土壤理化性质之间的回归分析
为了进一步探讨土壤理化性质对SOC及其各组分含量贡献的大小,利用逐步线性回归分析方法建立了SOC及其各组分与土壤理化性质之间的最优回归方程(表5).结果表明,本研究区SOC含量主要受土壤TN含量、C:N以及土壤电导率的共同影响,三者对SOC含量的综合贡献率达99.9%.土壤DOC含量主要受电导率影响,其贡献率达67%.土壤TN含量对土壤EOC含量的影响较大,其贡献率为45.7%.土壤TN含量和pH对微生物熵的综合贡献率为52.9%.土壤电导率对土壤DOC/SOC比值的贡献率达69%.土壤EOC/SOC比值主要受土壤电导率和容重影响,二者综合贡献率为59.3%.
3 讨论
3.1 水盐梯度下土壤活性有机碳组分的差异分析
水分是控制和维持湿地生态系统结构和功能的重要环境因子[39].水分条件的变化(包括土壤含水率和地表水位波动等)会影响湿地土壤-植物-大气连续体之间碳元素生物地球化学诸过程的方向与强度[40].在本研究中无论半咸水湿地还是淡水湿地,淹水频率高的低潮滩土壤MBC含量稍高于高潮滩,增幅分别为67.8%和38.8% (表1).这一结论与以往研究存在一定差异.侯翠翠等[41]通过野外考察三江平原生长季内不同水分条件下湿地表层土壤轻组有机碳与微生物活性发现,淹水环境下,由于供氧不足,土壤中微生物活性受到抑制,土壤MBC含量减少.万忠梅等[42]通过野外盆栽控制试验表明,土壤水分过干或过湿均不利于土壤MBC积累,干湿交替下土壤MBC含量最高.土壤MBC含量对水分变化的差异响应可能因为土壤微生物活性不仅受水分变化的限制,同时还受土壤营养物质可获得性和其他土壤理化性质(如土壤含盐量、pH和质地)等的影响[43-45].Lipson等[46]认为,枯落物的输入也是影响土壤MBC含量的重要因素.Sun等[32]的研究发现,在中国黄河口淹水频率高的低潮滩湿地相对高潮滩有更高的枯落物分解速率,因而可为土壤微生物提供更多有效的碳底物和营养元素,进而有相对较高的土壤MBC含量.同时,本研究通过分析高低潮滩土壤理化性质发现,无论半咸水湿地还是淡水湿地高低潮滩间土壤含水率和容重均无显著差异(表3),表明本研究中尽管高低潮滩间地表淹水频率不同,但土壤水分含量和通气性已无显著差异.已有研究表明土壤水分含量在有机碳分解矿化过程中有重要作用,Laiho 等[47]的研究表明,随着土壤水分含量增加,土壤微生物会由于氧气供给不足而使新陈代谢受阻,但适当的土壤水分含量促进土壤微生物活性,有利于有机质的分解矿化.本研究中高低潮滩间土壤微生物的表观活性差异不显著,暗示这种水分体系适合微生物的生长代谢,也表明土壤微生物对淹水环境有一定的适应机制.相关分析表明土壤MBC含量与土壤各理化因子间关系不显著(表4),逐步线性回归分析也无土壤理化因子进入回归方程(表5),表明在这种淹水环境下土壤各理化因子在控制土壤MBC含量上影响较小,不足以产生空间异质性上的显著影响.
微生物熵在2种湿地类型均表现为低潮滩高于高潮滩,且淡水湿地低潮滩显著高于半咸水湿地低潮滩(表2).微生物熵反映了输入土壤中的有机质向MBC的转化效率[48].微生物熵越高,表明微生物对土壤有机质的利用效率越高[49].不同水盐梯度下微生物熵的差异分布可能与研究区湿地土壤TN含量有关.因为氮限制环境下湿地生产力和有机质含量均降低[50],微生物为了维持其正常新陈代谢必须提高其对土壤有机质的利用效率,进而有更高的微生物熵.相关分析也发现微生物熵与土壤TN含量显著负相关(表4),逐步线性回归分析也表明土壤TN含量和pH对微生物熵的负作用综合贡献率达50%以上(表5).并且,塔礁洲淡水湿地和鳝鱼滩半咸水湿地相比,低潮滩面积相对较小,在频繁的潮水冲刷过程中,更不利于有机质的滞留和颗粒沉积物的沉降.因而在土壤TN含量、潮水及土壤pH值等的综合作用下,表现出2种湿地类型低潮滩均高于高潮滩,且淡水湿地低潮滩显著高于半咸水湿地低潮滩的空间分布特征.
土壤DOC,其含量在2种湿地类型高低潮滩间差异均不显著(表1).土壤DOC的含量取决于输入土壤中的DOC与输出之间的平衡.土壤DOC主要来源于近期的植物枯落物、根系分泌物及土壤有机质中的腐殖质等,是土壤微生物可直接利用的有机碳源[2,51],其输出主要通过微生物矿化成CO2释放到大气中或随水流失.本研究中2种湿地类型高低潮滩间差异均不显著可能与其相似的土壤微生物活性有关.而土壤EOC含量2种湿地均表现为高潮滩高于低潮滩(表1).土壤EOC是由一些分子结构简单、易分解的腐殖质和多糖等有机分子组成,短时间内可为土壤微生物提供能源[52].2种湿地土壤EOC含量高潮滩均高于低潮滩可能因为高潮滩有较低的淹水频率,受潮水冲刷作用小,有利于有机质的滞留和颗粒沉积物的沉降.
盐度是滨海湿地常见的环境胁迫因子之一.本研究中无论高低潮滩,土壤MBC含量均随盐度增加而降低,降幅分别为52.9%和43.1%(表1).滨海湿地盐度增加主要由海水入侵引起,海水入侵过程携带大量氯化钠和硫酸根等离子可通过引起过高离子强度[53]以及硫酸盐还原菌淘汰产甲烷菌[54]等途径来影响湿地土壤微生物群落组成与活性.一般认为,较高的离子强度可引起微生物的渗透压失衡,干扰细胞的生长和繁殖[55],从而影响土壤微生物的数量与活性.
本研究中,无论高低潮滩,土壤DOC和EOC含量均随盐度增加而增加,低潮滩的增幅更是达90%以上.这可能因为盐度能增加土壤有机质的溶解性,从而有利于不稳定的活性有机碳从土壤中释放出来[56].Wang等[57]在中国南部英罗湾红树林湿地研究中也发现土壤DOC和EOC含量均随盐度增加而增加.相关分析也表明土壤DOC和EOC含量及其分配比例均与土壤电导率呈正相关(表4).逐步线性回归分析发现土壤电导率对DOC及其分配比例的贡献率均达60%以上(表5).此外,土壤SOC含量和EOC含量与土壤TN含量极显著正相关(表4),逐步线性回归分析表明土壤TN含量和土壤电导率对SOC及其活性组分含量的影响显著(进入回归方程次数最多的环境因子,表5),进一步表明,相对水文条件变化,土壤氮素含量和盐度对本研究区湿地生态系统植物生长和土壤碳库演变影响的贡献更大.
3.2 土壤有机碳组分之间的相关性
以往一些研究证明土壤SOC与各活性组分之间彼此依存、关系密切.有研究表明土壤MBC与SOC和其他有机碳活性组分之间存在显著正相关[45,58].但本研究中,土壤MBC含量与SOC和其他有机碳活性组分之间均呈负相关,且与土壤DOC含量呈显著负相关(表4).土壤MBC含量与SOC、DOC和EOC含量之间的负相关可从以下几方面解释.一方面,SOC的质量,即底物的有效性,是限制土壤MBC的重要因子[59].本研究中输入土壤中的有机碳可能被土壤团聚体物理包裹而不被微生物分解利用,或者通过土壤黏粒的吸附作用而被保护起来[60].闽江河口位于中亚热带和南亚热带过渡地区,土壤发育较为成熟,铁铝富集.有大量研究表明铁铝矿物,特别是弱晶质的铁矿可与SOC通过化学或物理化学的键合作用,形成土壤有机-矿质复合体,从而保护土壤SOC不被微生物分解矿化[60-62].因而,SOC及其活性组分(DOC和EOC)在土壤颗粒的物理或化学结合保护下,不易被土壤微生物分解利用,导致尽管碳源较丰富,但MBC含量仍相对较低的情况.盛浩等[59]在对亚热带不同稻田土壤MBC含量分布特征的研究中也观测到这一现象.这一现象也从侧面印证了SOC的滞留和稳定不是分子属性问题,而是生态系统属性问题[63-64].另一方面,土壤MBC含量与SOC及其活性组分的负相关可能与土壤MBC周转速率高有关.土壤MBC的周转不仅反映微生物的活性,还指示土壤与微生物之间在有机碳周转和积累机理上的差异[65].土壤MBC周转速率越高,微生物活性越强,被分解矿化的有机碳就越多,SOC及其活性组分积累的就越少.
4 结论
4.1 闽江河口区无论半咸水湿地还是淡水湿地,土壤MBC含量随淹水频率增加而增加.同时,土壤微生物对这种淹水环境有一定的适应机制,水文条件变化不足以产生空间异质性上的显著差异.
4.2 土壤DOC、EOC含量及其分配比例随盐度的增加而增加,而土壤MBC含量和微生物熵随盐度的增加而降低.
4.3 相关分析与回归分析表明,土壤氮含量和盐度是影响闽江河口湿地生态系统植物生长和土壤碳库演变的重要限制性参数.
4.4 土壤MBC含量与SOC、DOC和EOC之间均呈负相关.暗示底物的有效性是限制土壤MBC的重要因子.也表明随着土壤MBC周转速率加快,SOC及其活性组分由于分解矿化而使积累量降低.
[1] 杨丽霞,潘剑君.土壤活性有机碳库测定方法研究进展[J]. 土壤通报, 2004,35(4):502-506.
[2] 万忠梅,郭 岳,郭跃东.土地利用对湿地土壤活性有机碳的影响研究进展[J]. 生态环境学报, 2011,20(3):567-570.
[3] Liang B C, MacKenzie A F, Schnitzer M, et al. Management- induced change in labile soil organic matter under continuous corn in eastern Canadian soils [J]. Biology and Fertility of Soils, 1997,26(2):88-94.
[4] 倪进治,徐建民,谢正苗.土壤生物活性有机碳库及其表征指标的研究 [J]. 植物营养与肥料学报, 2001,7(1):56-63.
[5] Ghani A, Dexter M, Perrott K W. Hot-water extractable carbon in soils: a sensitive measurement for determining impacts of fertilisation, grazing and cultivation [J]. Soil Biology and Biochemistry, 2003,35(9):1231-1243.
[6] 王清奎,汪思龙,冯宗炜,等.土壤活性有机质及其与土壤质量的关系[J]. 生态学报, 2005,25(3):513-519.
[7] Lunau M, Voss M, Erickson M, et al. Excess nitrate loads to coastal waters reduces nitrate removal efficiency: mechanism and implications for coastal eutrophication [J]. Environmental Microbiology, 2013,15(5):1492-1504.
[8] 曾志华,杨民和,佘晨兴,等.闽江河口区淡水和半咸水潮汐沼泽湿地土壤产甲烷菌多样性[J]. 生态学报, 2014,34(10):2674- 2681.
[9] Weston N B, Dixon R E, Joye S B. Ramifications of increased salinity in tidal freshwater sediments: Geochemistry and microbial pathways of organic matter mineralization [J]. Journal of Geophysical Research: Biogeosciences, 2006,111(G1):1-14.
[10] Chambers L G, Reddy K R, Osborne T Z. Short-term response of carbon cycling to salinity pulses in a freshwater wetland [J]. Soil Science Society of America Journal, 2011,75(5):2000-2007.
[11] Neubauer S C, Franklin R B, Berrier D J. Saltwater intrusion into tidal freshwater marshes alters the biogeochemical processing of organic carbon [J]. Biogeosciences, 2013,10(12):8171-8183.
[12] 王 纯,张璟钰,黄佳芳,等.盐度对感潮区淡水沼泽土壤甲烷产生潜力的影响.湿地科学, 2015,13(5):593-601.
[13] Wang C, Tong C, Chambers L G, et al. Identifying the salinity thresholds that impact greenhouse gas production in subtropical tidal freshwater marsh soils [J]. Wetlands, 2017,1-13.doi: 10.1007/s13157-017-0890-8.
[14] Morrissey E M, Gillespie J L, Morina J C, et al. Salinity affects microbial activity and soil organic matter content in tidal wetlands [J]. Global change biology, 2014,20(4):1351-1362.
[15] Pierfelice K N, Graeme Lockaby B, Krauss K W, et al. Salinity influences on aboveground and belowground net primary productivity in tidal wetlands [J]. Journal of Hydrologic Engineering, 2015:D5015002.
[16] Kaase C T, Kupfer J A. Sedimentation patterns across a Coastal Plain floodplain: The importance of hydrogeomorphic influences and cross-floodplain connectivity [J]. Geomorphology, 2016,269: 43-55.
[17] Boyd B M, Sommerfield C K, Elsey-Quirk T. Hydrogeomorphic influences on salt marsh sediment accumulation and accretion in two estuaries of the US Mid-Atlantic coast [J]. Marine Geology, 2017,383:132-145.
[18] 曾从盛,王维奇,翟继红.闽江河口不同淹水频率下湿地土壤全硫和有效硫分布特征[J]. 水土保持学报, 2011,(6):246-250.
[19] 王维奇,仝 川,贾瑞霞,等.不同淹水频率下湿地土壤碳氮磷生态化学计量学特征[J]. 水土保持学报, 2010,(3):238-242.
[20] Zhang W L, Zeng C S, Tong C, et al. Spatial distribution of phosphorus speciation in marsh sediments along a hydrologic gradient in a subtropical estuarine wetland, China [J]. Estuarine, Coastal and Shelf Science, 2015,154:30-38.
[21] Duhamel S, Nogaro G, Steinman A D. Effects of water level fluctuation and sediment–water nutrient exchange on phosphorus biogeochemistry in two coastal wetlands [J]. Aquatic Sciences, 2017,79(1):57-72.
[22] 仲启铖,关阅章,刘 倩,等.水位调控对崇明东滩围垦区滩涂湿地土壤呼吸的影响[J]. 应用生态学报, 2013,24(8):2141-2150.
[23] 盛宣才,吴 明,邵学新,等.模拟水位变化对杭州湾芦苇湿地夏季温室气体日通量的影响[J]. 生态学报, 2016,36(15).
[24] 高灯州,曾从盛,章文龙,等.闽江口湿地土壤有机碳及其活性组分沿水文梯度分布特征[J]. 水土保持学报, 2014,28(6):216- 221.
[25] 高灯州,章文龙,曾从盛,等.闽江河口湿地土壤生物和非生物因子与水淹频率的关系[J]. 湿地科学, 2016,(1):27-36.
[26] 仲启铖,王江涛,周剑虹,等.水位调控对崇明东滩围垦区滩涂湿地芦苇和白茅光合,形态及生长的影响[J]. 应用生态学报, 2014,25(2):408-418.
[27] 管 博,栗云召,夏江宝,等.黄河三角洲不同水位梯度下芦苇植被生态特征及其与环境因子相关关系[J]. 生态学杂志, 2014, 33(10):2633-2639.
[28] 仝 川,刘白贵.不同水淹环境下河口感潮湿地枯落物分解及营养动态[J]. 地理研究, 2009,(1):118-128.
[29] Chambers L G, Osborne T Z, Reddy K R. Effect of salinity pulsing events on soil organic carbon loss across an intertidal wetland gradient: A laboratory experiment [J]. Biogeochemistry, 2013,115(1):363-383.
[30] Neubauer S C. Ecosystem responses of a tidal freshwater marsh experiencing saltwater intrusion and altered hydrology [J]. Estuaries and Coasts, 2013,36(3):491-507.
[31] Howard R J, Biagas J, Allain L. Growth of common brackish marsh macrophytes under altered hydrologic and salinity regimes [J]. Wetlands, 2016,36(1):11-20.
[32] Sun Z, Mou X, Sun W. Potential effects of tidal flat variations on decomposition and nutrient dynamics of,andlitter in newly created marshes of the Yellow River estuary, China [J]. Ecological Engineering, 2016,93:175-186.
[33] 刘剑秋,曾从盛,陈 宁.闽江河口湿地研究[M]. 北京:科学出版社, 2006.
[34] 仝 川,曾从盛,王维奇,等.闽江河口芦苇潮汐湿地甲烷通量及主要影响因子[J]. 环境科学学报, 2009,29:207-216.
[35] Mou X J, Liu X T, Tong C, Sun Z G.Responses of CH4emissions to nitrogen addition andinvasion in Minjiang River estuary, southeast of China [J]. Chinese Geographical Science, 2014,24(5):562-574.
[36] 杨 平,徐 辉,万金红,等.台风“苏力”对闽江河口沼泽土壤间隙水中溶解性甲烷和乙酸等的影响[J]. 湿地科学, 2015,(5): 622-629.
[37] 鲁如坤.土壤化学农业分析方法[M]. 北京:中国农业科技出版社, 1999.
[38] Blair G J, Lefroy R D B, Lisle L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems [J]. Australian Journal of Agricultural Research, 1995,46(7):1459-1466.
[39] 盂宪民.湿地与全球环境变化.地理科学, 1999,19(5):385-391.
[40] Ström L, Christensen T R. Below ground carbon turnover and greenhouse gas exchanges in a sub-arctic wetland [J]. Soil Biology and Biochemistry, 2007,39(7):1689-1698.
[41] 侯翠翠,宋长春,李英臣,等.不同水分条件沼泽湿地土壤轻组有机碳与微生物活性动态[J]. 中国环境科学, 2012,32(1): 113-119.
[42] 万忠梅,宋长春,郭跃东,等.毛苔草湿地土壤酶活性及活性有机碳组分对水分梯度的响应[J]. 生态学报, 2008,28(12):5980- 5986.
[43] Xu M, Lou Y, Sun X, et al. Soil organic carbon active fractions as early indicators for total carbon change under straw incorporation [J]. Biology and Fertility of Soils, 2011,47(7):745-752.
[44] Siczek A, Frąc M. Soil microbial activity as influenced by compaction and straw mulching [J]. International Agrophysics, 2012,26(1):65-69.
[45] Wang W, Lai D Y F, Wang C, et al. Effects of rice straw incorporation on active soil organic carbon pools in a subtropical paddy field [J]. Soil and Tillage Research, 2015,152:8-16.
[46] Lipson D A, Schmidt S K, Monson R K. Carbon availability and temperature control the post-snowmelt decline in alpine soil microbial biomass [J]. Soil Biology and Biochemistry, 2000, 32(4):441-448.
[47] Laiho R, Laine J, Trettin C C, et al. Scots pine litter decomposition along drainage succession and soil nutrient gradients in peatland forests, and the effects of inter-annual weather variation [J]. Soil Biology and Biochemistry, 2004,36(7): 1095-1109.
[48] 张金波,宋长春.土地利用方式对土壤碳库影响的敏感性评价指标[J]. 生态环境, 2003,12(4):500-504.
[49] 黄 宇,汪思龙,冯宗炜,等.不同人工林生态系统林地土壤质量评价[J]. 应用生态学报, 2004,15(12):2199-2205.
[50] Bragazza L, Freeman C, Jones T, et al. Atmospheric Nitrogen Deposition Promotes Carbon Loss from Peat Bogs [J]. Proceedings of the National Academy of Sciences of the United States of America, 2006,103(51):19386-9.
[51] Guggenberger G, Kaiser K. Dissolved organic matter in soil: challenging the paradigm of sorptive preservation [J]. Geoderma, 2003,113(3):293-310.
[52] Zhang M, Zhang X, Liang W, et al. Distribution of soil organic carbon fractions along the altitudinal gradient in Changbai Mountain, China [J]. Pedosphere, 2011,21(5):615-620.
[53] Chambers L G, Guevara R, Boyer J N, et al. Effects of salinity and inundation on microbial community structure and function in a mangrove peat soil [J]. Wetlands, 2016,36(2):361-371.
[54] Weston N B, Vile M A, Neubauer S C, et al. Accelerated microbial organic matter mineralization following salt-water intrusion into tidal freshwater marsh soil [J]. Biogeochemistry, 2011, 102(1):135-151.
[55] Ikenaga M, Guevara R, Dean A L, et al. Changes in community structure of sediment bacteria along the Florida coastal everglades marsh–mangrove–seagrass salinity gradient [J]. Microbial ecology, 2010,59(2):284-295.
[56] Thurman E M. Organic geochemistry of natural waters (Vol. 2) [M]. Springer Science & Business Media, 2012.
[57] Wang G, Guan D, Zhang Q, et al. Distribution of dissolved organic carbon and KMnO4-oxidizable carbon along the low- to-high intertidal gradient in a mangrove forest [J]. Journal of Soils and Sediments, 2015,15(11):2199-2209.
[58] 肖 烨,黄志刚,武海涛,等.三江平原不同湿地类型土壤活性有机碳组分及含量差异[J]. 生态学报, 2015,35(23):7625-7633.
[59] 盛 浩,周 萍,袁 红,等.亚热带不同稻田土壤微生物生物量碳的剖面分布特征[J]. 环境科学, 2013,34(4):1576-1582.
[60] Lützow M V, Kögel-Knabner I, Ekschmitt K, et al. Stabilization of organic matter in temperate soils: mechanisms and their relevance under different soil conditions–a review [J]. European Journal of Soil Science, 2006,57(4):426-445.
[61] Song X Y, Spaccini R, Pan G, et al. Stabilization by hydrophobic protection as a molecular mechanism for organic carbon sequestration in maize-amended rice paddy soils [J]. Science of the Total Environment, 2013,458:319-330.
[62] Jiménez J J, Villar L. Mineral controls on soil organic C stabilization in alpine and subalpine soils in the Central Pyrenees: Insights from wet oxidation methods, mineral dissolution treatment and radiocarbon dating [J]. Catena, 2017,149(1):363- 373.
[63] Schmidt M W, Torn M S, Abiven S, et al. Persistence of soil organic matter as an ecosystem property [J]. Nature, 2011, 478(7367):49-56.
[64] 潘根兴,陆海飞,李恋卿,等.土壤碳固定与生物活性:面向可持续土壤管理的新前沿 [J]. 地球科学进展, 2015,30(8):940-951.
[65] 吴金水,肖和艾.土壤微生物生物量碳的表观周转时间测定方法[J]. 土壤学报, 2004,41(3):401-407.
致谢:在野外采样以及室内样品处理过程中,得到了福建师范大学地理科学学院王维奇老师的大力帮助;在样品分析过程中,得到了福建师范大学湿润亚热带生态地理过程教育部重点实验室的林燕语和彭园珍老师的帮助,在绘制采样点图时得到了福建师范大学地理科学学院谭立山同学的帮助,在此一并表示感谢!
Effects of hydrologic and salinity gradients on soil organic carbon composition in Min RiverEstuarine wetland.
WANG Chun1, LIU Xing-tu1*, TONG Chuan2,3, CHEN Xiao-xuan2,3, CHEN You-yang2,3, MOU Xiao-jie1, WAN Si-ang1
(1.Key Laboratory of Wetland Ecology and Environment, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130120, China;2.Institute of Geography, Fujian Normal University, Fuzhou 350007, China;3.Key Laboratory of Humid Subtropical Eco-geographical Process, Ministry of Education, Fujian Normal University, Fuzhou 350007, China)., 2017,37(10):3919~3928
In order to reveal the effects of hydrologic and salinity gradients on soil organic carbon composition in estuarine wetlands, the contents of soil organic carbon fractions along the hydrologic gradient within a freshwatermarsh and a brackishmarshes in the Min River Estuary were measured. Soil microibial biomass carbon (MBC) contents raised with increasing flooding frequency both in brackish-water marsh (67.8%) and freshwater marsh (38.8%), respectively. For both high tidal flat and low tidal flat, the MBC content in brackish-marsh was lower than that in freshwater marsh, and declining ranges were 52.9% and 43.1% high tidal flat via low tidal flat, respectively. Soil dissolved organic carbon (DOC) content in brackish marsh was higher than that in freshwater marsh, and increasing range were 56.7% and 105.6% high tidal flat via low tidal flat. Soil EOC content declined with increasing flooding frequency, and declining ranges were 18.0% and 50.1% brackish marsh via freshwater marsh, respectively. Soil EOC content in brackish marsh was higher than that in freshwater marsh, increased by 20.2% in high tidal flat and 97.4% in low tidal flat, respectively. The percentages of soil microbial entropy,DOC and EOCin SOC were 0.42% ~ 1.76%, 0.39% ~ 0.85% and 20.14% ~ 36.49%, respectively. Microbial entropy declined with increasing flooding frequency, while the proportions of soil DOC and EOCin SOC increased with increasing flooding frequency. Compared with the varied flooding environment, soil TN content and conductivity had a greater contribution on the SOC contents and its active components. Soil DOC and EOC contents were positively correlated with SOC content, and soil MBC content was negatively correlated with SOC, EOC and DOC contents, which implied substrate availability and soil MBC turnover rate exerted important impacts on controlling soil microbial activity and soil carbon pool accumulation in estuarine tidal marsh. Soil microbes increased with increasing flooding frequency, but they had a certain adaptation mechanism to the flooding environment. Elevated salinity increased soil DOC and EOC contents, but decreased soil MBC content. Soil nitrogen content and salinity which were important restrictive parameters demonstrated obviously effects on controlling soil carbon pool evolution in the tidal marsh ecosystemof the Min River estuary.
salinity;flooding environment;soil organic carbon;active fractions;Min River Estuary
X53,P95
A
1000-6923(2017)10-3919-10
王 纯(1982-),女,湖南省益阳人,博士后,主要从事湿地生物地球化学循环研究.发表论文20余篇.
2017-04-10
国家“973”项目(2013CB430401);中国博士后科学基金(2017M611337);国家自然科学基金资助(41371127)
*责任作者, 研究员, lxtmxh@163.com
