长链非编码RNA- p21诱导细胞凋亡在动脉粥样硬化中的作用
2017-11-01尹来波刘瑞英侯量胡思远朱志军朱佳龙
尹来波,刘瑞英,侯量,胡思远,朱志军,朱佳龙
(1.石河子大学医学院第一附属医院 心胸外一科,新疆 石河子 832008;2.山东省潍坊市市直机关医院 超声科,山东 潍坊 261061)
·论著·
长链非编码RNA-p21诱导细胞凋亡在动脉粥样硬化中的作用
尹来波1,刘瑞英2,侯量1,胡思远1,朱志军1,朱佳龙
(1.石河子大学医学院第一附属医院 心胸外一科,新疆 石河子 832008;2.山东省潍坊市市直机关医院 超声科,山东 潍坊 261061)
目的探讨lincRNA- p21通过调控细胞凋亡参与冠心病的发生发展过程,及其对动脉粥样硬化的影响。方法选取我院50例冠心病患者,用ElISA方法检测患者血清中lincRNA- p21含量和凋亡蛋白Bcl- 2含量。提取大鼠心肌内皮细胞制备大鼠动脉硬化模型,过表达或抑制lincRNA- p21后,CCK- 8法检测细胞存活率;构建lincRNA- p21质粒注射进入小鼠颈动脉损伤模型,HE染色观察小鼠颈动脉内膜新生。Caspase- 3活性检测试剂盒检测内皮细胞凋亡蛋白Caspase - 3活性;Western blot法检测凋亡蛋白Bcl- 2和BAX蛋白表达水平。结果冠心病患者血清内lincRNA- p21表达显著下降,差异有统计学意义(P<0.05)。大鼠原代内皮细胞中过表达lincRNA- p21显著降低细胞存活率,显著上升Caspase - 3活性,显著增加BAX表达,显著下降BCL- 2表达;lincRNA- p21抑制组细胞存活率明显增高,Caspase 3活性显著下降,促凋亡蛋白BAX水平显著降低,抗凋亡蛋白BCL- 2水平显著增加,差异有统计学意义(P<0.05)。小鼠体内局部沉默lincRNA- p21后,血管内/中膜厚度比明显增加,Caspase - 3活性明显降低,Bcl- 2蛋白表达增加且BAX蛋白表达降低,差异有统计学意义(P<0.05)。结论lincRNA- p21可通过调控细胞凋亡参与冠心病的发生发展过程。
lincRNA- p21;冠心病;细胞凋亡;BCL- 2
冠状动脉粥样硬化性心脏病(简称冠心病),是指冠状动脉血管发生动脉粥样硬化病变而引起血管腔狭窄或阻塞,造成心肌缺血、缺氧或坏死而导致的心脏病,临床常见病的多发病,严重威胁人群健康[1- 3]。长链非编码RNA(long non- codingRNAs,lncRNAs)是一类长度超过200 nt的ncRNA, 近年来lncRNAs被证明在干细胞的发育,多能性,细胞生长和细胞凋亡等中发挥重要的生物学功能,然而lncRNAs在心血管系统疾病中的作用尚不十分清楚[4- 6]。间长链非编码RNA- p21 (long intergenic non- coding RNA- p21, lincRNA- p21)做为转录因子P53的下游靶基因,其转录受P53正性调控并参与P53的调控过程[7,8]。研究显示,lincRNA- p21可以直接与靶基因mRNA序列结合发挥基因调控的功能,参与调控动脉粥样硬化的过程、调节血管平滑肌和巨噬细胞的细胞增殖分化,及刺激受损血管增生等作用,但lincRNA- p21的生物学功能并未完全明确[9,10]。在本研究中,我们旨在探讨lincRNA- p21调控细胞凋亡参与冠心病的发生发展过程,为阐明lncRNA在心血管系统疾病中的生理病理作用及调控机制提供有力基础。
1 材料与方法
1.1 实验材料
20只SPF级雄性Wistar大鼠(180- 200 g)和清洁型C57BL/6J小鼠(雄性18- 20 g)购买于北京维通利华实验动物技术有限公司。293T,人胚肾细胞购自中国科学院细胞库。重组穿梭质粒和包装质粒 pGag/Pol、pRev、pVSV- G 由上海吉玛构建制备。一抗多克隆抗体Bcl- 2,BAX,ACTB购于美国SantaCruz生物技术有限公司;二抗碱性磷酸酶标记山羊抗兔IgG购于北京中杉金桥生物技术有限公司;CCK- 8试剂盒购自日本同仁化学研究所;Caspase 3活性检测试剂盒购于上海碧云天生物工程有限公司;MAPK和PKC Elisa试剂盒均购于美国Abcam生物技术有限公司。IncRNA p21质粒及RealTime- PCR配套引物,均购自广州RiboBio公司;Real- Time PCR试剂盒购自日本TaKaRa公司。
1.2 患者材料
选取2016 年3月~2017年1月石河子大学医学院第一附属医院在心内科住院50例为冠心病患者,均行冠脉造影明确诊断。冠心病确诊为冠脉造影时左主干、前降支、回旋支、右冠脉或其主要分支的血管直径狭窄(至少 1 支冠状动脉直径狭窄>50%)。平均发病年龄为25~72岁,性别不限,为冠心病组。同时选取体检健康、且性别年龄均匹配的 30例健康体检者,为对照组。研究前均对所有患者进行完整的病史采集及体格检查。
1.3 大鼠内皮细胞动脉粥样硬化模型相关实验
1.3.1 大鼠原代内皮细胞制备 大鼠饲养在SPF级动物实验室,12 h/12 h昼夜循环光照,温度为(22±2) ℃。适应性喂养一周后,大鼠提取原代心肌内皮细胞。10%水合氯醛 (350 mg·kg-1),腹腔注射后麻醉大鼠,迅速开胸剖取心脏和主动脉,保留左心室心肌组织,研磨后加入含20%胎牛血清的DMEM高塘培养基37 ℃、5% CO2条件下培养。细胞贴壁后换液,加入高脂血清(胆固醇含量50 mg·dl-1)培养24 h,制备动脉粥样硬化模型,取对数生长期细胞用于后续实验[11,12]。
1.3.2 细胞慢病毒载体构建及细胞转染 lincRNA- p21质粒接种293T细胞培养过夜,加入穿梭质粒和包装质粒共转染293T细胞,6 h后弃去转染液,加入细胞培养液继续培养。收集转染后72 h的293T细胞上清液,用PBS液重悬病毒沉淀,- 80 ℃保存。将大鼠原代内皮细胞接种于24孔板中,待细胞融合率达到40%时,加入慢病毒原液转染,并加入Polybrene试剂增强感染效率,37%、5%CO2培养箱中培养。24 h后换液,加入新完全培养液继续培养,待细胞贴满培养皿底即传代,实验分阴性对照组、lincRNA- p21过表达组和IncRNA p21抑制组。
1.3.3 CCK- 8法检测细胞存活率 取转染后对数生长期细胞制备成细胞悬液,接种到96孔板中(100 μl/孔),37 ℃、5% CO2条件下过夜培养。各组细胞分别给予阿霉素和紫杉醇联合培养48 h后,更换培养基后加入10 μl CCK8,37 ℃、5% CO2条件下培养2 h,使用酶标仪在450 nm波长处测定OD值,计算细胞存活率。
1.4 小鼠动脉粥样硬化疾病模型相关实验
1.4.1 lincRNA- p21体内局部沉默的小鼠颈动脉线损伤动脉粥样硬化疾病模型的建立 小鼠颈动脉线损伤这一动脉粥样硬化疾病小鼠模型是研究动脉粥样硬化的经典动物模型[13,14]。模型建立手术操作如下[15]:10%水合氯醛 (350 mg·kg-1)腹腔注射麻醉小鼠,常规备皮消毒,钝性分离皮下组织及肌肉,暴露并分离左侧颈动脉,持粗糙金属导丝管持续抽插2- 3次损伤动脉内膜。30 μl的lincRNA- p21沉默重组病毒原液处理动脉内膜损伤部位,室温孵育30 min。缝合创口,待小鼠苏醒后以 60%高脂饲料诱导动脉粥样硬化。
1.4.2 HE染色 小鼠腹腔注射麻醉后,取颈动脉标本常规脱水透明,并用石蜡包埋连续切片6张(5μm/张)。苏木素染色,二甲苯透明,采用HE染色观察颈动脉内膜新生状态。
1.5 大小鼠内皮细胞相关检测
1.5.1 Caspase - 3活性检测试剂盒检测Caspase- 3活性 取心肌内皮细胞制备成细胞悬液,收集细胞于冰上裂解1 h,4 ℃离心后收集蛋白上清液。96孔板中加入用缓冲液稀释的蛋白样本50 μl反应缓冲液(reaction buffer含10 mM DTT)及5 μl 4 mM LEHD- pNA底物,37 ℃孵育1 h,在酶标仪405 nm测定其吸光值。Caspase- 3活性单位(U)=(各组A值- 凋零组A值)/标准曲线斜率,每次重复3孔。
1.5.2 Western Blot检测Bcl- 2,BAX蛋白表达水平 取心肌内皮细胞制备成细胞悬液,收集细胞于冰上裂解2 h,4 ℃离心后收集蛋白上清液。测定蛋白浓度后,将各组蛋白浓度总量调整为30 μg·μl- 1,加入到10%变性聚丙烯酰胺凝胶中电泳,转膜后,用牛血清白蛋白封闭1 h,加入一抗4 ℃过夜。隔天用TBST洗膜3次,并加入二抗孵育1 h。TBST洗膜两次,TBS洗膜1次。用ECL显色液进行显色,并对各泳道条带进行灰度扫描,得出相应的蛋白表达量。
1.6 患者血清lincRNA- p21和Bcl- 2含量检测
清晨用非抗凝管采集患者空腹抽静脉5 ml,血样静置后3000 rpm·min-1离心10 mim,分离血浆取血清待检。Trizol法提取细胞内总RNA。用含有gDNA Eraser的PrimeScriptRT试剂盒逆转录合成cDNA,用SYBR Premix Ex TaqII试剂盒进行实时荧光定量Real- time PCR分析。每个实验至少重复3次。分析:通过计算循环阈值CT值来确定基因的表达量。公式:ΔΔCT=(CT.实验组的基因-CT.实验组管家基因)-(CT.对照组目的基因-CT.对照组管家基因)。目的mRNA的量=2-ΔΔCT。
1.7 统计学处理
2 结 果
2.1 转染效率及CCK8法检测细胞的存活率
CCK- 8法检测细胞存活率显示,与对照组的细胞存活率相比,lincRNA- p21过表达组细胞存活率显著降低,细胞发生凋亡;lincRNA- p21抑制组细胞存活率明显增高,差异有统计学意义(P<0.01),如图2。
2.2 HE染色结果
结果表明,与对照组相比,体内局部沉默lincRNA- p21后小鼠血管内/中膜厚度比明显增加,说明lincRNA- p21的沉默可以明显加速损伤部位动脉内膜的新生,如图3。
2.3 Caspase - 3活性检测试剂盒检测结果
Caspase- 3活性检测结果可知,与对照组相比,lincRNA- p21过表达组Caspase - 3活性显著上升,细胞发生凋亡;lincRNA- p21抑制组Caspase- 3活性显著下降,促进细胞凋亡的发生,差异有统计学意义(P<0.01),如图4A。体内局部沉默lincRNA- p21后小鼠血管内皮细胞Caspase - 3活性显著下降,差异有统计与对照组相比,差异有统计学意义(*,P<0.05; **,P<0.01)
A.转染效率;B.细胞存活率
图2转染效率及CCK-8法检测细胞存活率

A- C.颈动脉HE染色;D.动脉内/中膜厚度比
与对照组相比,差异有统计学意义(*,P<0.05);与小鼠siNC组相比,差异有统计学意义(#,P<0.05)
图3HE染色检测结果(200X)
学意义(P<0.01),说明lincRNA- p21 的沉默可以明显减弱细胞凋亡的发生,如图4B。
2.4 Bcl- 2,BAX蛋白表达水平
用WesternBlot法测得结果可知,原代内皮细胞lincRNA- p21过表达组促凋亡蛋白BAX水平显著上升,抗凋亡蛋白Bcl- 2水平显著下降;原代内皮细胞
lincRNA- p21抑制组促凋亡蛋白BAX水平显著降低,抗凋亡蛋白BCL- 2水平显著增加,抑制细胞凋亡的发生,差异有统计学意义(P<0.05),如图5A和5B。体内局部沉默lincRNA- p21后小鼠内皮细胞后抗凋亡蛋白Bcl- 2水平显著上升,促凋亡蛋白BAX水平显著下降,差异有统计学意义(P<0.05),如图5C和5D。
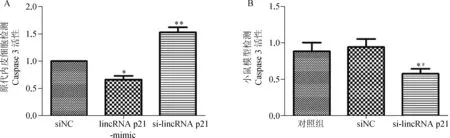
A.原代内皮细胞Caspase 3活性检测结果;B.小鼠模型Caspase - 3活性检测
与对照组相比,差异有统计学意义(*,P<0.05; **,P<0.01);与小鼠siNC组相比,差异有统计学意义(#,P<0.05)
图3Caspase-3活性检测试剂盒检测结果

A.原代内皮细胞各组蛋白表达含量测定;B.原代内皮细胞BAX和Bcl- 2蛋白含量表达;C.小鼠模型各组蛋白表达含量测定;D.小鼠模型BAX和Bcl- 2蛋白含量表达
与对照组相比,差异有统计学意义(*,P<0.05; **,P<0.01)与小鼠siNC组相比,差异有统计学意义(#,P<0.05)
图5WesternBlot蛋白表达含量
2.5 患者血清lincRNA- p21和Bcl- 2含量检测
患者血清检测显示,与对照组相比,冠心病患者血清内lincRNA- p21和Bcl- 2水平显著下降,差异有统计学意义(P<0.01);如图1。

与对照组相比,差异有统计学意义(*,P<0.05; **,P<0.01)
图1Real-timePCR检测lincRNA-p21和BCL-2含量检测
3 讨 论
lncRNAs是一类长度超过200 nt的ncRNA, 近年来lncRNAs被证明在干细胞的发育,多能性,细胞生长和细胞凋亡等中发挥重要的生物学功能,然而lncRNAs在心血管系统疾病中的作用尚不十分清楚[16,17]。lincRNA- p21做为转录因子P53的下游靶基因,可以直接与靶基因mRNA序列结合发挥基因调控的功能,并参与P53的调控过程[18,19]。冠心病在心肌缺血早期,心肌细胞死亡的主要形式是细胞凋亡,伴随心肌损伤的病理生理全过程[20]。研究表明,lincRNA- p21 在 ApoE- /- 小鼠动脉粥样硬化斑块组织中的表达量显著降低,提示 lincRNA- p21 可能与 AS 的发生发展存在一定的相关性[15]。但lincRNA- p21是否参与调解冠心病的发生发展过程并未明确。
本研究首先采集冠心病患者血清,对血清内lincRNA- p21的表达进行检测。结果表明,与对照组相比,冠心病患者血清内lincRNA- p21表达显著下降,说明lincRNA- p21在冠心病的发展过程中具有重要的作用。通过对临床病人标本中lincRNA- p21表达的检测,进一步确认了lincRNA- p21与动脉粥样硬化的相关性。从而将长链非编码RNA与动脉粥样硬化的发生发展相联系,为动脉粥样硬化这一疾病的治疗提供了全新的理论依据和潜在的分子靶标。
经大鼠原代培养内皮细胞,并对其进行高血脂动脉硬化造模后,观察lincRNA- p21在其中发挥的生物学功能。与对照组的细胞存活率相比,lincRNA- p21过表达组细胞存活率显著降低,Caspase 3活性显著上升;lincRNA- p21抑制组细胞存活率明显增高,Caspase 3活性显著下降,说明过表达lincRNA- p21可显著促进细胞凋亡的发生,影响细胞的增殖过程。同时,结果表明,lincRNA- p21过表达组促凋亡蛋白BAX水平显著上升,抗凋亡蛋白BCL- 2水平显著下降;lincRNA- p21抑制组促凋亡蛋白BAX水平显著降低,抗凋亡蛋白BCL- 2水平显著增加,说明沉默lincRNA- p21后,细胞凋亡发生受到抑制,细胞凋亡相关蛋白收到有效调控。
为了检测lincRNA- p21在体内对动脉硬化内膜新生及内皮细胞凋亡的影响,本研究采用动脉粥样硬化疾病小鼠模型构建方法[13,14],建立了lincRNA- p21体内局部沉默的小鼠动脉粥样硬化模型。结果表明,体内局部沉默lincRNA- p21后,血管内/中膜厚度比明显增加,说明 lincRNA- p21 的沉默可以明显加速损伤部位动脉内膜的新生,并能降低凋亡蛋白Caspase 3活性,促进Bcl- 2表达和抑制BAX表达,说明沉默lincRNA- p21后,细胞凋亡发生受到抑制,lincRNA- p21可有效参与动脉粥样硬化的发展过程。这一发现说明lincRNA- p21在体内体外均可以对动脉粥样硬化冠心病内皮细胞的增殖凋亡产生影响,进一步增强了其通过对细胞增殖和凋亡的调节进而影响动脉粥样硬化这一设想的可能性,但lincRNA- p21是通过何种方式参与了细胞增殖和凋亡的调控,受其调控的信号分子如何仍需进一步研究探讨。
综上所述,lincRNA- p21调控细胞凋亡参与冠心病的发生发展过程,可通过基因调控改善细胞增殖与凋亡,促进冠心病的发生发展。了解lincRNA的调节过程,为阐明lncRNA在心血管系统疾病中的生理病理作用及调控机制提供有力基础,具有重要的临床意义。
[1] 王朋,杨明会,李绍,等.冠心病心绞痛寒凝血瘀证大鼠心肌细胞凋亡及BAX、BCL- 2蛋白表达[J].中华中医药学刊,2014,32(3):506- 508.
[2] 高阅春,何继强,姜腾勇,等.冠心病患者冠状动脉病变严重程度与冠心病危险因素的相关分析[J].中国循环杂志,2012,27(3):178- 181.
[3] HU Y W,ZHAO J Y,LI S F,et al.RP5- 833A20.1 /miR- 382- 5p / NFIA- dependent signal transduction pathway contributes to the regulation of cholesterol homeostasis and inflammatory reaction[J].Arterioscler Thromb Vasc Biol,2015,35(1):87- 101.
[4] 卢耀星,杨建安,刘银河,等.心血管疾病相关IncRNA调控机制的研究进展[J].中国动脉硬化杂志,2016,24(5):537- 540.
[5] WAPINSKI O,CHANG H Y.Long noncoding RNAs and human disease[J].Trends Cell Biol,2011,21(6):354- 361.
[6] 胡春松,吴清华,胡大一.中国心血管现状:挑战与对策[J].中华高血压杂志,2015,23(7):625- 626.
[7] HUARTE M,GUTTMAN M,FELDSER D,et al.A large intergenic noncoding rna induced by p53 mediates global gene repression in the p53 response[J].Cell,2010,142(3):409- 419.
[8] YOON J H,ABDELMOHSEN K,SRIKANTAN S,et al.LincRNA- p21 suppresses target mrna translation[J].Mol Cell,2012;47:648- 655.
[9] WU G Z, CAI Y ,YANG Y J ,et al.Lincrna- p21 regulates neointima formation and atherosclerosis by enhancing p53 activity[J].Atherosclerosis ,2014 ,235 (2) :e24- e24.
[10] 郗瑞席,白瑞娜,李立志,等.IncRNA与心血管疾病[J].医学研究杂志,2015,44(8):9- 191.
[11] 韩学杰,张立石,沈绍功,等.痰瘀同治方对实验性动脉粥样硬化家兔主动脉、心肌及内皮细胞形态学的影响[J].中国动脉硬化杂志,2004,12(5):515- 518.
[12] 宁艳霞.类固醇激素合成急性调节蛋白(StAR)在血管内皮细胞和小鼠肝脏内的表达及意义[D].复旦大学,2006.
[13] OHNO T,GORDON D,SAN H,et al.Gene therapy for vascular smooth muscle cell proliferation after arterial injury[J].Science,1994,265(5173):781- 784.
[14] VARENNE O,PISLARU S,GILLIJNS H,et al.Local adenovirus- mediated transfer of human endothelial nitricoxide synthase reduces luminal narrowing after coronary angioplasty in pigs[J].Circulation,1998,98(9):919- 926.
[15] 吴庚泽.长链非编码RNA- p21(lincRNA- p21)在动脉粥样硬化中的作用和分子机制研究[D].第三军医大学,2014.
[16] GUTTMAN M,RINN J L.Modular regulatory principles of large non- coding rnas[J].Nature,2012,482:339- 346.
[17] WILUSZ J E,SUNWOO H,SPECTOR D L.Long noncoding rnas:Functional surprises from the rna world[J].Genes Dev,2009,23(13):1494- 1504.
[18] JACKSON S P,BARTEK J.The DNA- damage response in human biology and disease[J].Nature,2009,461(7267):1071- 1078.
[19] WU G,CAI J,HAN Y et al.LincRNA- p21 regulates neointima formation,vascular smooth muscle cell proliferation,apoptosis,and atherosclerosis by enhancing p53 activity[J].Circulation,2014,130(17):1452- 1465.
[20] 吕风华,高建芝,崔佳佳,等.叶酸对冠心病大鼠血清同型半胱氨酸和血管内皮生长因子的影响[J].西安交通大学学报,2011,32(4):449- 452.
Theeffectoflongnon-codingRNA-p21-inducedapoptosisinatherosclerosis
YINLai-bo1,LIURui-ying2,HOULiang1,HUSi-yuan1,ZHUZhi-jun1,ZHUJia-long
(1.FirstAffiliatedHospital,SchoolofMedicine,ShiheziUniversity,shihezi832008,China;2.DepartmentofUltrasound,WeifangMunicipalOfficialHospital,Weifang261061,China)
Objective: To investigate the effect of lincRNA- p21 on the development of coronary heart disease (CHD) via regulating apoptosis.MethodsFifty patients with coronary heart disease were randomly divided into two groups. The content of lincRNA- p21 and the content of apoptotic protein BCL- 2 were detected through ELISA method. Rat cardiac endothelial cells were extracted to prepare for rat arteriosclerosis model with overexpression or inhibition of lincRNA- p21, followed by the detection of cell viability using CCK- 8 method. LincRNA- p21 plasmid was constructed and implanted into the mice carotid artery injury model, Carotid artery neointima in mice was detected by HE staining. Caspase 3 activity assay kit was used to detect the expression of Caspase 3 in endothelial cells. The expression of BCL- 2 and BAX protein was detected by Westernblot.ResultThe expression of lincRNA- p21 in serum of patients with coronary heart disease was significantly decreased(P<0.05). The cell viability of lincRNA- p21 overexpression in rat primary endothelial cells was significantly reduced, Caspase 3 activity was significantly increased,BAX protein expression was significantly increased,and BCL - 2 protein expression was significantly decreased. LincRNA- p21 inhibation was significantly increased the cell survival, Caspase 3 activity was significantly decreased, the level of pro- apoptotic protein BAX was significantly decreased, anti- apoptotic protein BCL- 2 levels increased significantly, the difference was statistically significant(P<0.05). Intravascular/medial thickness ratio was increased significantly after mice localized with lincRNA- p21 inhibition, Caspase 3 activity was significantly decreased, Bcl- 2 protein expression was increased and BAX protein expression was decreased, the difference was statistically significant (P<0.05).ConclusionlincRNA- p21 could be involved in the development of coronary heart disease by regulating apoptosis.
lincRNA- p21; coronary heart disease; apoptosis; BCL- 2
2017- 02- 26
2017- 09- 11
尹来波(1983-),男,山东潍坊人,硕士,主治医师 ,主要研究方向:心胸外科疾病。E- mail:yinlaibo7297@163.com
朱佳龙 yinxiangyu6666@sohu.com
尹来波,刘瑞英,侯量,等.长链非编码RNA- p21诱导细胞凋亡在动脉粥样硬化中的作用[J].东南大学学报:医学版,2017,36(5):816- 821.
R541.4
A
1671- 6264(2017)05- 0816- 06
10.3969/j.issn.1671- 6264.2017.05.027
(本文编辑:孙茂民)
