大豆lox2基因表达方式分析及其启动子预测
2017-10-20刘晓鑫
赵 艳,刘晓鑫,翟 莹,曹 慧, 吴 琼
(1. 齐齐哈尔大学生命科学与农林学院,黑龙江 齐齐哈尔 161006;2.中种国际种子有限公司长春分公司,吉林 长春 130113)
大豆lox2基因表达方式分析及其启动子预测
赵 艳1,刘晓鑫2,翟 莹1,曹 慧1, 吴 琼1
(1. 齐齐哈尔大学生命科学与农林学院,黑龙江 齐齐哈尔 161006;2.中种国际种子有限公司长春分公司,吉林 长春 130113)
在不同逆境胁迫、激素条件下,检测大豆各器官中lox2基因表达方式,并对其启动子的序列及功能进行预测。利用qPT-PCR方法,检测大豆lox2基因表达方式;此外,通过PCR方法,克隆获得大豆lox2基因5’端上游序列,利用启动子在线软件进行预测分析。结果表明,该基因受干旱诱导,诱导表达量随处理时间增强;在盐、低温、ABA条件处理下,诱导表达下降,下降的表达量不随处理时间而变化。大豆lox2基因在大豆根、茎、叶、花中的相对表达量低,而在种子中的相对表达量高。克隆获得大豆lox2基因5’端上游2016 bp序列,命名为LP。在线软件预测分析,表明LP序列中含有多种典型的种子特异表达元件,如SEF3motif、SEF4 motif、E-box、AACACA、ACGT、Skn-1 motif、AATAAA等;及与逆境、激素诱导相关的元件,如LTR、MBS、W-box、AAAG-motif、ABRE等。推测大豆LP启动子具有调控下游基因大量表达在种子中的特性,同时具有受脱落酸及其它逆境诱导表达的特性。
大豆;lox2基因;启动子;克隆
大豆是我国重要的经济作物,籽粒中含有丰富的蛋白质、脂类和多种营养元素,磷、铁、钙矿质比其他作物高几十倍,并含有多种维生素, 特别是大豆中含有人体不能合成的8 种必需氨基酸,是其他谷类作物不能比拟的。自大豆基因组测序后,现已进入大豆功能基因组研究,主要集中在对基因及调控元件如启动子等的研究中,对大豆中基因表达方式的研究有助于基因功能的研究;对启动子的研究能够从本质上揭示基因表达方式的原因,有助于启动子功能的研究,为大豆基因工程研究提供有利工具。
脂肪氧化酶(Lipoxygenase,简称脂氧酶,Lox) 在动植物界中广泛存在,在真核生物中参与不饱和脂肪酸的代谢,且在动植物不同发育阶段存在着不同的类型[1]。最早在大豆种子中获得,大豆种子中含有丰富的Lox(占种子蛋白质含量的1 %~2 %),可催化不饱和脂肪酸的加氧反应[2]。现已从大豆种子中分离出Lox1、Lox2和Lox3(包含Lox3a和Lox3b) 3种不同性质的同工酶[3]。现今对Lox的研究主要集中在粮食作物、谷类作物种子研究中,如大豆种子Lox能直接与蛋白质和氨基酸结合,产生豆腥味、苦涩味,降低食品风味和营养品质[4];在水稻种子中,Lox是稻谷陈化变质的关键酶,进而导致种子衰老和陈化变质[5];在小麦中,过高的Lox活性会加快脂质的氧化反应,使籽粒氧化变质,从而使其储存期变短,造成粮食损失,及麦类食品的营养价值下降[6]。为避免lox基因的表达,提高作物种子应用品质及利于贮藏,现已从遗传育种的角度开展了一些缺失体品种的选育研究[7]。但lox基因在不同环境条件下或大豆器官中的表达情况如何,颜克亮等[8],利用抑制性消减杂交技术和半定量PCR,发现lox2基因在种子胚中高表达,但lox基因在不同环境条件下的表达情况,未有相关报道。为更明确大豆lox2基因在逆境胁迫、激素、大豆各器官中的表达方式,本研究中根据NCBI中大豆lox2基因序列,利用qRT-PCR方法,检测lox2基因在盐、干旱、低温、脱落酸(ABA)及在大豆根、茎、叶、花和种子中的相对表达量;并克隆lox2基因5’端上游序列,利用启动子在线预测软件对lox2基因启动子的表达方式进行分析。
1 材料与方法
1.1 供试材料
大豆品种吉豆2号为齐齐哈尔大学分子生物学实验室保存;大肠杆菌DH 5α感受态购自北京索莱宝公司;TRIzol reagent 购自Invitrogen公司;Oligo(dT)18、M-MLV反转录酶、Recombinant RNase Inhibitor、ExTaq、pMD18-T克隆载体、SYBP Premix Ex TaqⅡ荧光定量试剂盒、限制性内切酶PstI 和NcoI均购自Takara公司;DNA凝胶回收试剂盒购自维特洁公司;引物合成、测序由北京六合华大基因公司完成。
1.2 大豆材料处理
逆境及激素条件下材料处理:将大豆水培幼苗分别置于含有NaCl (200 mmol·L-1) 和PEG8000(20 %)的Hoaglands营养液中进行盐和干旱胁迫处理;大豆幼苗置于4 ℃培养箱中进行低温处理;对大豆水培幼苗喷施脱落酸(200 μmol·L-1,ABA)[9]。在处理0、1、2、5、10、24 h分别取样,迅速放于液氮中,置-80 ℃中保存备用。
各器官的取材:分别取大豆根、茎、叶、花、未成熟种子(开花后30 d,30DAF)和成熟种子(90DAF),迅速放于液氮中,置-80 ℃中保存备用。
1.3 QRT-PCR
根据RNA提取试剂盒步骤,提取大豆各种备用材料的总RNA;以总RNA为模板,按照逆转录的步骤分别合成cDNA的第一条链,进行qRT-PCR反应。以大豆持家基因β-tubulin(GMU12286)为内参,内参引物分别为(T1: 5’-GGAAGGCTTTCTTGCATTG GTA-3’; T2: 5’- AGTGGCATCCTGGTACTGC-3’)[10]。根据NCBI中,大豆lox2基因的核苷酸序列(L06038.1),利用Primer Premier 5.0 软件设计检测该基因的引物,上游引物(QF: 5’-AAGAGGAGGAAAGCCGTGAA-3’)和下游引物(QR:5’-GAATCAAAGTGGCGAGGAGAC-3’)。利用BIO-RAD CFX96实时定量PCR仪,反应循环参数为:95 ℃ 10 s,58 ℃ 20 s,72 ℃ 30 s。每次取样点设3次技术重复,实验共设3次生物学重复[11-12]。
1.4 大豆lox2 基因启动子序列的克隆及预测分析
取大豆幼苗叶片0.1 g为材料,采用CTAB法提取大豆的基因组DNA[13]。根据大豆基因组序列(http://www.phytozome.net/soybean)和NCBI中大豆lox2基因序列,设计特异引物扩增大豆lox2基因ATG的5’ 端上游序列。PCR扩增中的上游、下游引物分别带有PstI与NcoI酶切位点,上游引物为LF:5’-GGGCTGCAGTTATGGCTTCTTGTTTGTCTTA C-3’,下游引物为LR:5’-GGGCCATGGCTTTGCCAAAACCTACCAATAAC-3’。PCR反应条件为预变性95 ℃ 5 min;94 ℃ 30 s,54 ℃ 40 s,72 ℃ 1 min,共30个循环;72 ℃后延伸7 min。将PCR扩增片段切胶回收并连接到pMD18-T克隆载体上,获得重组质粒,鉴定后进行测序。利用DNAMAN软件进行同源性比对。在线启动子分析软件PlantCARE(http://www.dna.affrc.go.jp/PLACE/signalscan.htmL)分析启动子序列中存在的顺式作用元件。
2 结果与分析
2.1 大豆lox2基因的诱导表达分析
利用qRT-PCR方法,在干旱(PEG)、盐(NaCl)、低温(4 ℃)和激素(ABA)不同条件时间处理下,对大豆lox2基因的相对表达量进行分析。结果显示:大豆lox2基因在干旱胁迫下,相对于对照,表达量先降低,5 h后,随着处理时间的延长,表达量逐渐升高,当处理24 h,相对表达量为7.47,说明大豆lox2基因受干旱诱导,且诱导表达量随处理时间增强(图1)。
大豆lox2基因在盐条件处理下,相对于对照,当处理为1 h,相对表达量下降30 倍左右,以后的2、5、10 和24 h 处理时间下,相对表达量与处理1 h的表达量变化不大(图2)。大豆lox2基因在低温、ABA条件处理下,表现出与盐处理条件下相似的规律(图3~4)。说明大豆lox2基因受盐、低温、ABA诱导,诱导表达下降,下降的表达量不随处理时间而变化。
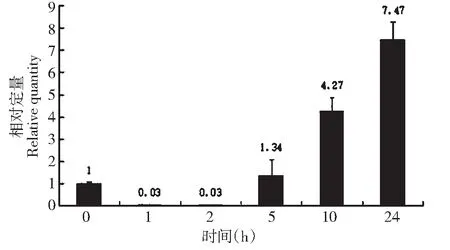
图1 干旱胁迫下lox2基因的表达Fig.1 Expression of lox2 gene under drought stress
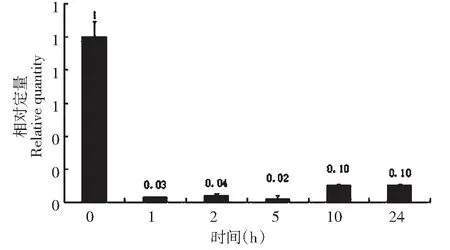
图2 盐胁迫下lox2基因的表达Fig.2 Expression of lox2 gene under salt stress

图3 低温胁迫下lox2基因的表达Fig.3 Expression of lox2 gene under cold stress

图4 ABA诱导lox2基因的表达Fig.4 Expression of lox2 gene under ABA induction
2.2 大豆lox2基因的器官表达分析
利用qRT-PCR方法,大豆持家基因β-tubulin为内参,对大豆根、茎、叶、花、未成熟种子和成熟种子中lox2基因的相对表达量进行分析。结果显示(图5):大豆lox2基因在大豆根、茎、叶、花中的表达低,相对于叶中的表达量,大豆lox2基因在根、茎、花中表达活性分别为5.48,0.12,0.46;而在大豆种子中,尤其是未成熟种子中的相对表达活性高,相对于叶中的表达量,大豆lox2基因在未成熟种子和成熟种子中的表达活性分别为125.54 和26.84。

图5 大豆lox2基因在大豆各器官中的表达Fig.5 Expression of soybean lox2gene in different soybean organs

M.2000 bp DNA marker; 1.PCR产物 M. 2000 bp DNA marker; 1.PCR amplification of LP图6 LP 的扩增产物Fig.6 PCR amplification of LP

M.2000 bp DNA marker;1~2. Pst I和Nco I双酶切片段 M.2000 bp DNA marker;1-2.Pst I and Nco I digestion of pMD18-T-LP图7 pMD18-T-LP酶切鉴定Fig.7 Restriction enzyme digestion identification of pMD18-T-LP

图8 大豆lox2基因启动子序列中的顺式作用元件Fig.8 Cis-elements of lox2 gene promoter
2.3 大豆lox2基因启动子序列的克隆
根据NCBI中大豆lox2基因的序列,及大豆基因组序列,设计引物,PCR扩增大豆lox2基因ATG上游序列,获得长度为2016 bp的序列,命名为LP(图6)。将该片段连接到pMD18-T载体上,获得重组质粒pMD18-T-LP。从大肠杆菌菌液中提取质粒,用PstI和NcoI双酶切鉴定,结果得到与预期相符的片段(图7)。利用DNAMAN软件,将测序结果与大豆基因组中该段序列进行同源性比对,同源性达到100 %,说明该段序列在不同大豆品种间无差别。
2.4 大豆LP启动子的序列分析
启动子在线预测软件分析结果表明,LP启动子中含有多种典型的种子特异表达元件(图8),与胚高表达相关的顺式作用元件(SEF3 motif和SEF4 motif)[14],常出现在参与三酰基甘油合成和植物种子特异表达基因的启动子中的元件(E-box)[15],种子特异表达的顺式作用元件(AACACA、ACGT、Skn-1 motif、AATAAA)[16-19];及与激素、逆境诱导相关元件,如ABRE(脱落素响应元件)[20],LTR(低温应答元件)[21],MBS(参与干旱反应,是MYB的结合位点)[22],W-box(WRKY转录因子的结合位点)[23],AAAG-motif(DOF转录因子的结合位点)[24]。
3 讨 论
本研究中对大豆lox2基因在不同逆境胁迫和激素处理下的表达情况进行分析。大豆lox2基因在干旱处理下,表达呈上调趋势,表达量在24 h达到最大值,是对照表达量的7.47倍,说明lox2基因受干旱上调诱导明显。在盐、低温和ABA处理条件下,lox2基因的表达量均下降,说明lox2基因受盐、低温和ABA的下调诱导。因此推测,大豆种子在盐、低温和ABA处理条件下,可降低豆腥味、苦涩味产生。此外,lox2基因在不同逆境胁迫和激素处理下的表达情况,很可能与大豆lox2基因启动子中存在逆境、激素诱导相关顺式元件,及其位置有关。
本研究对大豆lox2基因在各器官中的差异表达情况进行分析。lox2基因在大豆种子中,尤其是未成熟种子(30DAF)的相对表达活性很高,是根、茎、叶、花中表达量的23~1000倍,这与大豆种子中含有丰富的Lox的密切相关。根据大豆lox2基因在各器官中的表达情况看,该基因为种子特异表达基因。进一步克隆lox2基因上游的启动子序列,基因的5’端上游调控序列中包含大量的顺式调控元件,往往可能位于距离ATG5’端上游更远端,所以本研究中在不明确有效启动子序列时,克隆了ATG上游2016 bp的序列,能够尽可能的包含了较多的调控元件(或是增强子)。克隆的LP启动子序列中序列中包含多种与种子特异表达相关的顺式元件,因此,该基因的启动子很可能具有调控下游基因在大豆种子中大量表达的活性,仍需进一步实验验证。
4 结 论
利用qPT-PCR方法检测表明大豆lox2基因受干旱、盐、低温、ABA诱导,且该基因在大豆种子中的相对表达量高;其启动子序列LP中含有多种典型逆境、激素诱导相关元件及种子特异表达元件,推测大豆LP启动子具有调控下游基因大量表达在种子中的特性,同时具有受脱落酸及其它逆境诱导表达的特性。
[1]阎彩霞,张彩英,马峙英,等. 大豆种子脂肪氧化酶研究进展[J]. 河北农业科学,2005,9(2):82-87.
[2]Patel D D, Patel R R, Thakkar V R. Purification, characterization and application of lipoxygenase isoenzymes from Lasiodiplodia theobromae[J]. Appl Biochem Biotech, 2015, 175(1):513-525.
[3]Zhao Y, Zhou J, Xing D. Phytochrome B-mediated activation of lipoxygenase modulates an excess red light-induced defence response in Arabidopsis[J]. J Exp Bot, 2014, 65(17):4907-4018.
[4]麻 浩,官春云,何小玲,等. 大豆种子脂肪氧化酶缺失基因控制豆腥味效果的研究[J]. 中国农业科学,2001,34(4):367-372.
[5]蒋家月,吴跃进,张从合,等. 水稻种胚脂肪氧化酶Lox1基因的遗传分析及定位[J]. 核农学报,2012,26(4): 629-633.
[6]吴培培,宋 双,张福彦,等. 黄淮麦区部分小麦种质脂肪氧化酶活性分析及等位基因检测[J]. 中国农业科学,2015,48(2):207-214.
[7]Lee K J, Hwang J E, Velsam V, et al. Selection and molecular characterization of a poxygenase-free soybean mutant line induced by gamma irradiation[J]. Theor Appl Genent(TAG), 2014,127(11):2405-2413.
[8]颜克亮,陈 波,姚家玲,等. 大豆种子形态建成期与成熟期正反抑制消减文库构建及差异表达基因分析[J]. 武汉大学学报(理学版),2008,54(2):202-208.
[9]Zhai Y, Wang Y, Li Y J, et al. Isolation and molecular characterization of GmERF7, a soybean ethylene-response factor that increases salt stress tolerance in tobacco[J]. Gene, 2013(2):174-183.
[10]Zhao Y, Sha W, Wang Q Y, et al. Molecular cloning and activity analysis of a seed-specific FAD2-1B gene promoter fromGlycinemax[J]. Cell Mol Biol, 2015, 61(4):85-89.
[11]Acharya S, Ranjan R, Pattanaik S, et al. Efficient chimeric plant promoters derived from plant infecting viral promoter sequences[J]. Planta, 2014, 239:381-396.
[12]Pan Y L, Ma X, LiangI H W, et al. Spatial and temporal activity of the foxtail millet (Setariaitalica) seed-specific promoterpF128 [J]. Planta, 2015, 241:57-67.
[13]Xiao Y H, Luo M, Fang W G, et al. PCR walking in cotton genome using YADE method [J]. Act Genet Sin, 2002, 29:62-66.
[14]Chung K J, Hwang S K, Hahn B S, et al. Authentic seed-specific activity of the Perilla oleosin 19 gene promoter in transgenic Arabidopsis[J]. Plant Cell Rep, 2008, 27:29-37.
[15]Kim M J, Kim J K, Shin J S. The SebHLH transcription factor mediates trans-activation of the SeFAD2 gene promoter through binding to E- and G-box elementsv [J]. Plant Mol Biol, 2007, 64 (4): 453-466.
[16]Luo K, Zhang G, Dwng W, et al. Functional characterization of a cotton late embryogenesis-abundant D113 gene promoter in transgenic tobacco[J]. Plant Cell Rep, 2008, 27:707-717.
[17]Gao M J, Lydiate D J, Li X, et al. Repression of Seed Maturation Genes by a Trihelix Transcriptional Repressor in Arabidopsis Seedlings[J]. Plant Cell, 2009, 21(1):54-71.
[18]Fauteux F, Stromvik M V. Seed storage protein gene promoters contain conserved DNA motifs in Brassicaceae, Fabaceae and Poaceae[J]. BMC Plant Biol, 2009(9):126.
[19]Xie C, Zhang B, Wang D, et al. Molecular cloning and Characterization of anachene-seed-specific promoter from motherwort(LeonurusjaponicusHoutt)[J]. Biotech Let, 2011, 33:167-172.
[20]Zhao Y, Shao S L, Li X W, et al. Isolation and activity analysis of a seed abundant soyAP1 gene promoter from soybean[J]. Plant Mol Biol Rep, 2012, 30:1400-1407.
[21]Xu W R, Yu Y H, Ding J H, et al. Characterization of a novel stilbene synthase promoter involved in pathogen-and Stress-inducible expression from Chinese wild Vitis pseudoreticulata[J]. Planta, 2010, 231(2):475- 487.
[22]刘晓敏,张莉弘,刘金亮,等. 玉米逆境诱导型启动子克隆及其植物表达载体构建[J].生物技术通报, 2011(3):86-90.
[23]Zhou Q Y, Tian A G, Zou H F, et al. Soybean WRKY-type transcription factor genes, GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants[J]. Plant Biotechnol J, 2008(6):486-503.
[24]Nalbandu K, Kohnehrouz B B, Saeed K A, et al. Isolating Barley (HordeumvulgareL.) B1 Hordein Gene Promoter and Using Sequencing Analaysis for the Identification of Conserved Regulatory Elements By Bioinformatic Tools[J]. Afr J Biotechnol, 2012, 11(29): 7378-7387.
(责任编辑 李山云)
ExpressionPatternAnalysisofSoybeanlox2GeneandPredictionofPromoter
ZHAO Yan1, LIU Xiao-xin2, ZHAI Ying1, CAO Hui1, WU Qiong1
(1.College of Life Science and Agroforestry, Qiqihaer University, Heilongjiang Qiqihaer 161006, China; 2.China Seed International Seed Co., LTD.Changchun Branch, Jilin Changchun 130113, China)
The expression patterns of soybeanlox2 gene in soybean organs under different adversity stress and hormone were detected, and the sequence and function oflox2 gene promoter were predicted. The expression pattern of soybeanlox2 gene was detected by qRT-PCR. Moreover, the 5’-flanking upstream sequence of soybeanlox2 gene by PCR method was isolated, which was analyzed by online software. The result showed that the activity oflox2 gene was increased by drought with time; the activity oflox2gene was decreased under salt, low temperature and ABA treatment, which were not related to the time. There was a little activity in roots, stems, leaves and flowers, but there was highest activity in seeds. Moreover, the 5’-flanking upstream sequence of soybeanlox2 gene, named LP, was isolated by PCR method, and the length was 2016 bp. Sequence analysis revealed that this fragment contained a series of motifs related to seed-specific promoters, hormone and stress-inducible promoters, such as SEF3 motif, SEF4 motif, E-box, AACACA, ACGT, Skn-1 motif, AATAAA, LTR, MBS, W-box, AAAG-motif, ABRE and so on.It was inferred that LP promoter possess the function of driving downstream gene expression abundantly in seeds and inducing by abscisic acid and other adversity.
Soybean;lox2 gene; Promoter; Cloning
1001-4829(2017)3-0505-06
10.16213/j.cnki.scjas.2017.3.004
S643
A
2016-03-15
黑龙江省自然科学基金项目(C201458);黑龙江省普通本科高等学校青年创新人才培养计划(UNPYSCT_2016090)
赵 艳(1981-),女,吉林吉林人,博士,副教授,研究方向植物分子生物学,E-mail:zhaoyan3053877@163.com。
