氮素水平对不同氮效率苎麻纤维发育关键酶的影响
2017-10-18陈继康谭龙涛喻春明朱爱国陈平高钢陈坤梅熊和平
陈继康,谭龙涛,喻春明,朱爱国,陈平,高钢,陈坤梅,熊和平
(中国农业科学院麻类研究所,长沙410205)
氮素水平对不同氮效率苎麻纤维发育关键酶的影响
陈继康,谭龙涛#,喻春明,朱爱国,陈平,高钢,陈坤梅,熊和平*
(中国农业科学院麻类研究所,长沙410205)
以氮高效(H2000-03)和氮低效(册亨家麻)2个基因型苎麻品种为材料,设置氮素水平(0、6、9、12、15 mmol/L)开展温室盆栽试验,研究氮素调控苎麻纤维发育关键酶的生理机制。结果表明,苎麻蔗糖合成酶(SS)、β-1,3-葡聚糖酶和吲哚乙酸氧化酶(IAAO)活性随氮素水平提高呈现逐渐增加的趋势,并在超过9mmol/L后增幅显著下降。各酶活性随生长发育时期均呈单峰变化趋势,表现为旺长期>幼苗期>成熟期。氮高效基因型的相关酶活性均高于氮低效基因型,且SS和β-1,3-葡聚糖酶活性在幼苗期和旺长期达到显著水平,IAAO活性在旺长期和成熟期达到显著水平(p<0.05)。相关酶对苎麻原麻产量具有重要影响,不同基因型对其响应特征不同,其中氮高效基因型的响应较敏感。苎麻纤维发育存在适氮水平,氮高效基因型可耐受更高氮素水平,并表现出显著较高的生产力和稳产性。
苎麻;氮效率;纤维;酶;氮素水平
苎麻(Boehmeria nivea L.)是我国重要的韧皮纤维作物。苎麻纤维产量和品质的形成是品种遗传性和环境因子共同作用的结果。氮素是作物生长发育的肥料三要素之一,其对苎麻纤维产量及品质的影响备受关注。研究显示,适宜的氮素可以提高纤维品质,而土壤氮素过剩或亏缺均会导致纤维品质下降。李元沅等[1]、王春桃等[2]认为,土壤全氮、水解氮与苎麻纤维细度呈显著或极显著正相关。周兆德等[3]认为,合理施用氮肥可以提高肥料利用率,随着氮肥用量增加,苎麻的株高、茎粗、皮厚等都相应增长。但欧阳铎声等[4]认为氮素含量过高,则单细胞的细胞壁较薄,出麻率、纤维品质都有所降低,纤维支数降低的程度比较明显。可见,以往研究由于施氮量、苎麻品种等因子的差异而没有得出统一的结论。
为获得高产,苎麻生产长期以来提倡“重施肥”。据测算,每生产50 kg原麻,需吸收氮5.05~7.80 kg,氮农学效率不足10,远低于主要粮食作物。氮效率的基因型差异在作物间、品种间普遍存在,明确氮高效基因型的生理机制、筛选氮高效基因型、进行品种改良是提高氮素利用效率最有效的措施之一。β-1,3-葡萄糖酶[5,6]、蔗糖合成酶[7]是影响纤维发育的关键酶,吲哚乙酸(IAA)是影响作物生长发育的关键酶[8],并且在纤维细胞分化中具有重要作用[9],其酶活的高低表征着作物纤维发育与品质形成的特征。申素芳[10]对5个品种苎麻纤维素合成关键酶随植株生长发育的动态变化特征进行了分析,认为蔗糖合成酶与纤维素含量、单纤维长径比正相关,β-葡聚糖酶活力与单纤维断裂强度正相关,但没有涉及相关酶对氮素的响应。陈平等[11]、蒋杰等[12]、刘昱翔等[13]、田志坚等[14]分别以不同的苎麻材料开展了苎麻纤维素合成酶基因的克隆和表达,但均没有上升到作物生理响应水平,也没有探讨不同氮效率基因型的差异。因此,基于以往研究的不足,本文设置不同氮素水平处理,对两个具有显著差异氮效率的苎麻基因型纤维发育关键酶进行对比分析,明确不同氮效率苎麻对氮素的响应特征,旨在提高苎麻氮素利用效率,为苎麻纤维产量与品质的提升提供理论依据。
1 材料与方法
1.1 试验材料
采用盆栽试验,于2014年在湖南省长沙市岳麓区的中国农业科学院麻类研究所温室进行。供试的苎麻基因型为前期水培试验,从30个苎麻资源中筛选出的氮低效基因型册亨家麻和氮高效基因型H2000-03,其中册亨家麻在9 mmol/L氮素营养液中的氮素利用效率(根据地上部生物量计算)为15.81%,H2000-03为46.01%,两个品种的鲜茎出麻率无差异。
苎麻采用嫩梢扦插繁殖。选取生长状况一致的扦插苗栽于盆中,每盆一株,土壤最大持水量为63.8%,每盆装干土30 kg,试验塑料盆直径60 cm,高55 cm,土壤自然风干、过筛去杂后装盆,用水沉实,供试土壤养分含量为:全氮 1.01 g/kg,速效氮 75.1 mg/kg,速效磷 16.25 mg/kg,速效钾37.41 mg/kg,有机质含量 2.35 mg/kg。
1.2 试验设计
试验设5个处理,分别用氮素水平为0、6、9、12、15mmol/L的营养液浇灌,代号分别为N0、N6、N9、N12和N15,每个处理3个重复,每个重复15株。为避免水分胁迫影响,每3天浇300~500mL营养液,采用营养液培养60 d,共浇灌20次,总灌水量为8 L,其他时期根据土壤水分状况浇灌清水。大量元素营养液配方参考Hoagland营养液配方,微量元素营养液配方参考文献[15],具体见表1。

表1 大量元素培养液配方Tab.1 Components ofmass elements in nutrient solution
试验从5月10日栽入盆中开始,分别在幼苗期(第15 d)、旺长期(第40 d)、成熟期(第54 d)取样,取样时间为上午10点,取样部位为从顶部数第3~8片功能叶和植株中部韧皮组织,等量混合,先用液氮固定30 s,然后放入-80℃冰箱中保存,用以测定样品相关酶活性,每个样品3个重复。收取所有苎麻植株,采用4BM-260型苎麻剥麻机剥制原麻,立即称重,得到单株鲜麻重。
1.3 测定项目与方法
苎麻样品蔗糖合成酶、β-1,3-葡聚糖酶和吲哚乙酸氧化酶活性参照《现代植物生理学实验指南》测定[16]。数据采用Microsoft Excel 2016进行初步整理和制图;采用IBM SPSS Statistics19.0软件进行处理,ANOVA进行方差分析,用Duncan法在0.05水平上进行多重比较;采用Pearson法进行一般线性相关分析。
2 结果与分析
2.1 不同氮效率苎麻蔗糖合成酶活性对氮素的响应

图1 氮素水平对不同时期苎麻蔗糖合成酶活性的影响Fig.1 Effects of nitrogen levels on SSactivity of different growth stages
苎麻植株蔗糖合成酶活性在三个时期整体上均表现为随着氮素水平升高逐渐增大(见图1)。随氮素水平升高,蔗糖合成酶活性的增长较平稳。苎麻植株蔗糖合成酶活性随生长发育时期呈单峰变化,但在前期保持较高水平而在后期有大幅下降。在同一氮素水平下,各生长发育时期氮高效基因型H2000-03的蔗糖合成酶活性均高于氮低效基因型册亨家麻,并且除幼苗期N0处理和N15处理均达到显著水平(p<0.05)。
2.2 不同氮效率苎麻β-1,3-葡聚糖酶活性对氮素的响应
苎麻植株β-1,3-葡聚糖酶活性在幼苗期、旺长期和成熟期随氮素水平升高逐渐增大(见图2)。相邻氮素水平间,N9处理与N6处理下苎麻植株β-1,3-葡聚糖酶活性的变化最大。其中,N9处理下H2000-03在三个生长发育时期分别较N6处理高71.8%、34.0%和38.3%;册亨家麻在N9处理下分别较N6处理高63.6%、63.7%和29.9%。氮素水平超过或低于N9处理,均表现出处理间差异变小的趋势。
苎麻植株β-1,3-葡聚糖酶活性随生长发育阶段呈单峰变化,表现为旺长期>幼苗期>成熟期。氮高效基因型苎麻H2000-03在各个时期和所有氮素水平下,均高于氮低效基因型苎麻册亨家麻,并且在幼苗期、旺长期的所有氮素水平下及成熟期的高氮水平下达到显著水平(p<0.05)。

图2 氮素水平对不同时期苎麻β-1,3-葡聚糖酶活性的影响Fig.2 Effects of nitrogen levels onβ-1,3-glucanade activity of ramie at different growth periods
2.3 不同氮效率苎麻吲哚乙酸氧化酶活性对氮素的响应
吲哚乙酸氧化酶活性在三个时期整体上均表现为随着氮素水平升高逐渐增大,而增长比率则逐渐下降(见图3)。其中幼苗期、旺长期和成熟期H2000-03吲哚乙酸氧化酶活性,N9处理较N0处理分别高70.0%、50.5%和68.7%;N15处理较N9处理分别高9.5%、13.3%和0.4%。吲哚乙酸氧化酶活性随生长发育时期呈先上升后下降的趋势,表现为旺长期>幼苗期>成熟期。同一氮素水平下,氮高效基因型H2000-03的吲哚乙酸氧化酶活性整体高于氮低效基因型册亨家麻,其中旺长期各施氮处理均显著较高(p<0.05),并在成熟期维持了显著较高的优势。
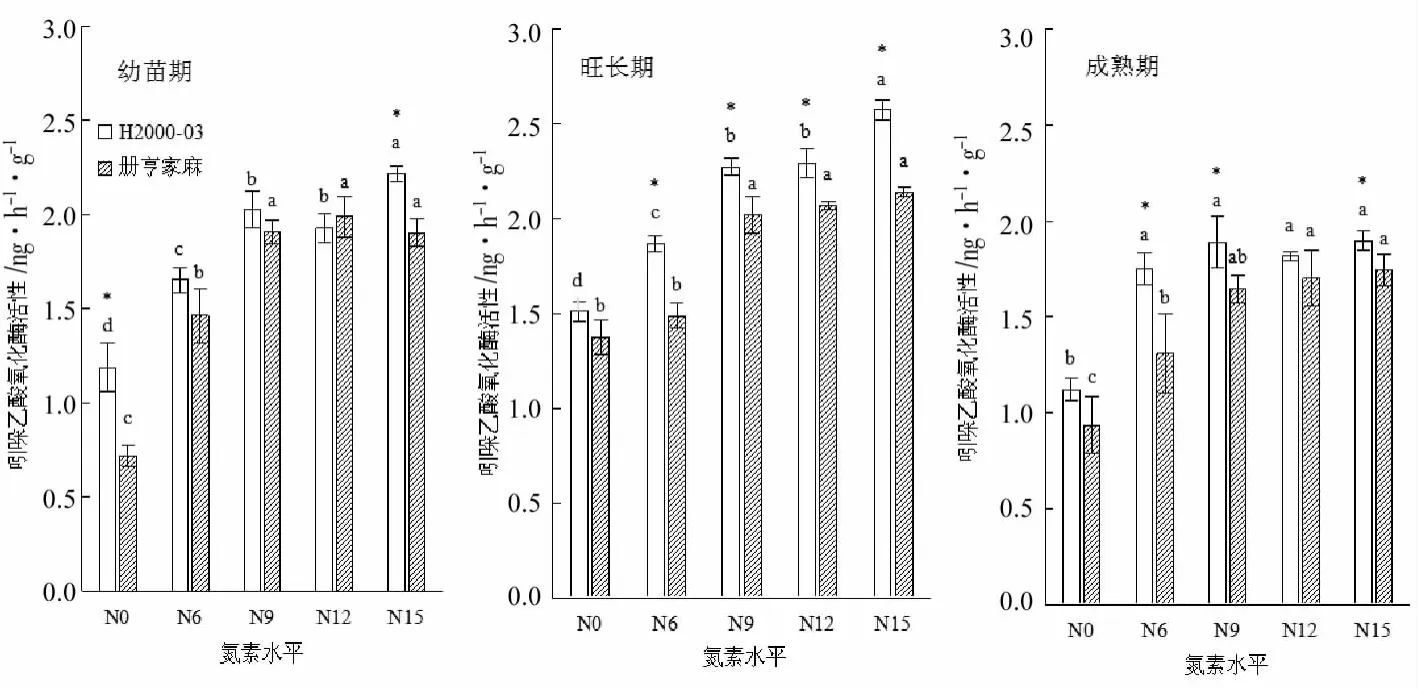
图3 氮素水平对不同时期苎麻吲哚乙酸氧化酶活性的影响Fig.3 Effects of nitrogen levels on IAAO activity of ramie at different growth periods
2.4 不同氮效率苎麻原麻产量与相关酶活性的关系
表2为随着氮素水平的变化,两个苎麻基因型各生长发育时期纤维发育关键酶活性与最终原麻产量的一般线性相关分析。氮高效基因型苎麻H2000-03的原麻产量与各生长发育时期的3种纤维发育关键酶活性均呈显著(p<0.05)或极显著(p<0.01)相关。氮低效基因型苎麻册亨家麻则没有达到显著水平。同一基因型、同一生长发育时期的不同酶之间整体呈显著或极显著正相关关系,表现出较高的协调性和一致性。
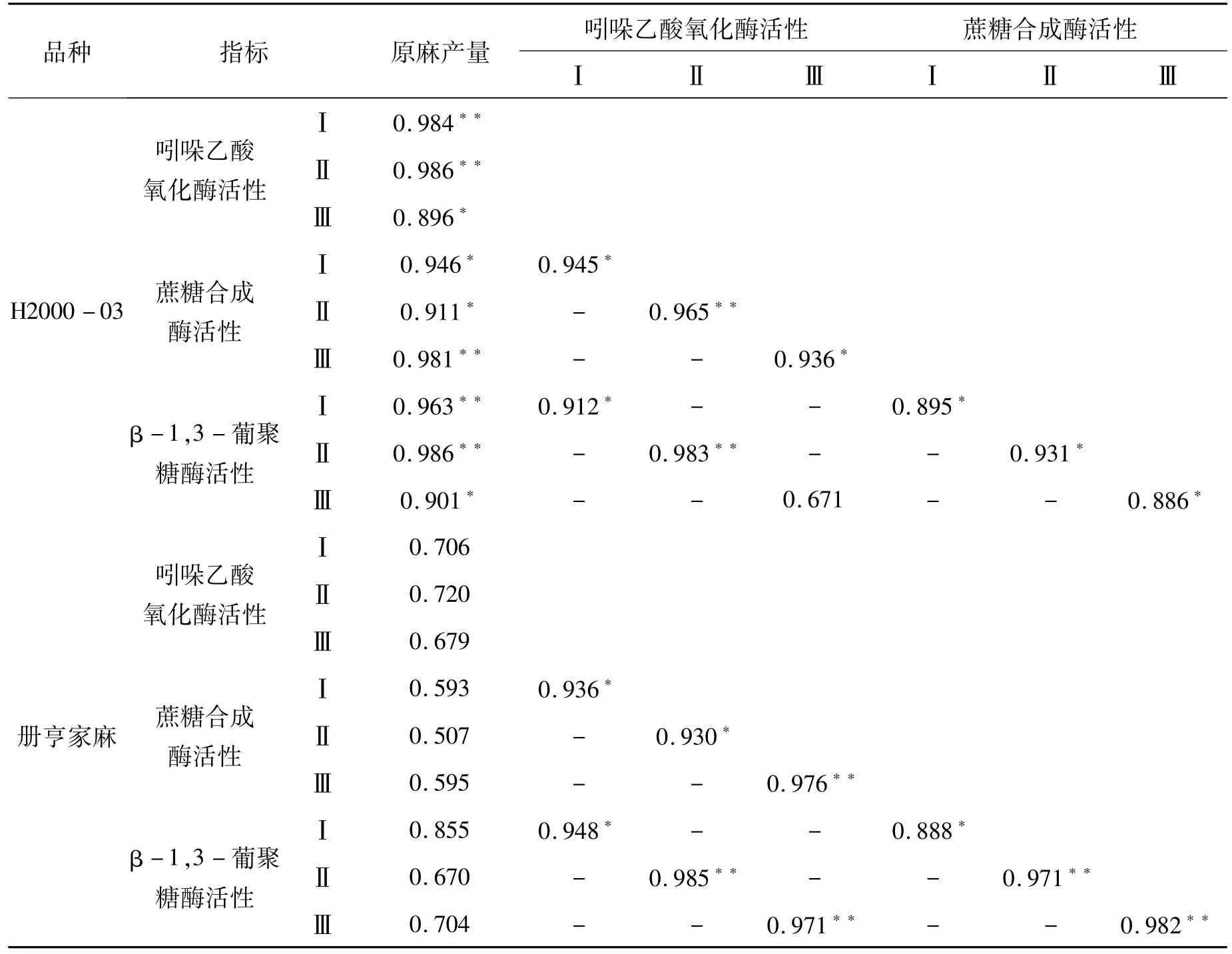
表2 不同氮效率苎麻纤维发育关键酶活性与原麻产量的相关分析Tab.2 Correlation analysis of enzyme activity related to fiber development and raw fiber yield of ramie with different nitrogen efficiency
3 讨论
目前对纤维发育关键酶的研究主要集中在棉花上面,对苎麻的报道较少。蔗糖合成酶(sucrose synthetase,SS)和 β-1,3-葡聚糖酶(β-1,3-Glucanase)被普遍认为是调控纤维素生物合成的关键酶,其活性的高低在很大程度上影响到纤维素的累积特性[17,18]。蔗糖合成酶活性的变化直接影响纤维素的合成速度和累积质量[19],而且蔗糖合成酶对氮素变化的响应较为敏感[20]。β-1,3-葡聚糖酶可内切β-1,3-葡聚糖糖苷键水解β-1,3-葡聚糖,在增加细胞可塑性的同时为糖代谢提供底物[21]。不同于棉花的是,苎麻为韧皮纤维作物,其纤维与茎秆的伸长密切相关,株高一直是衡量苎麻产量的重要农艺性状之一。苎麻纤维发育受IAA的调控作用较棉花可能更强。吲哚乙酸氧化酶(IAAO)可氧化分解IAA,调节细胞伸长,并对次生壁加厚发育等生理过程具有一定的调控作用[22]。因此,本文将上述三种酶作为影响苎麻纤维发育的关键酶进行分析,更符合苎麻生长发育的特点。
高相彬等[23]研究棉花纤维发育相关酶对棉铃对位叶氮浓度变化的响应发现,其活性均随棉铃对位叶氮浓度的上升呈先升后降的趋势。张文静等[24]则发现β-1,3-葡聚糖酶活性在次生壁加厚发育过程中呈下降趋势,SS和IAAO活性变化均呈单峰曲线,基因型间差异主要表现在酶活性的大小和峰值出现的时间。冯营等[25]发现适氮水平下,棉纤维中后期β-1,3-葡聚糖酶活性的提高有利于纤维素延长快速积累期及平缓积累;高氮水平纤维发育相关酶活性没有提高,虽然高于缺氮水平,但低于适氮水平。本研究发现,在试验涉及的范围内,SS、β-1,3-葡聚糖酶、IAAO随氮素水平提高整体表现为上升趋势,但产量呈单峰曲线,且相关酶活性在高氮条件下随氮素水平提高增幅显著变缓。苎麻相关酶活性随生长发育时期呈单峰变化。这与已有研究关于纤维发育存在适氮水平的结论是一致的,但相关酶活性的变化特征与棉花不同,显示出两种作物不同的纤维发育特征。
在棉纤维快速伸长期,POD和IAAO活性较低,有利于细胞快速伸长;而进入次生壁加厚期时,活性迅速增加[26],通过调节内源ABA/IAA,对纤维素的生物合成和次生壁加厚发育进行调控[27]。本研究中IAAO活性在各时期均随氮素水平的升高逐渐提高,总体表现为旺长期酶活性最高,幼苗期次之,成熟期最低,且氮高效基因型高于氮低效基因型。表明在苎麻旺长期,IAAO活性较高,韧皮部进入次生壁加厚期,且氮高效基因型一直具有较高的IAAO活性,能促进苎麻韧皮部的加厚。从三种酶活性较好的协调性和一致性推断,苎麻纤维从植株顶端至基部逐渐由以伸长为主变为以加厚为主,这与棉花种子纤维的伸长和加厚规律不同。相关酶在植株各部位的分布和作用可能存在较大的差异,需要进一步的研究验证。
本研究发现,氮高效基因型的相关酶活性均高于氮低效基因型,且SS和β-1,3-葡聚糖酶活性在幼苗期和旺长期达到显著水平,IAAO活性在旺长期和成熟期达到显著水平。可见,氮高效基因型较氮低效基因型在生长中前期具有显著较高的纤维伸长和累积能力,而中后期具有显著较高的纤维加厚能力。育种和栽培中可以对这些特性加强利用和改良。
氮高效基因型苎麻的原麻产量在各氮素水平下均高于氮低效基因型,而且在高氮胁迫下两者表现出较大的差异。但氮素水平高于N9处理时,氮高效基因型随氮素水平提高保持了较高的原麻产量,而氮低效基因型原麻产量显著下降。一方面说明不同基因型苎麻纤维发育对氮素水平的响应特征不一致,另一方面说明不同基因型苎麻纤维发育对其生理状况的响应特征也不一致,生产和育种中需要注意与其他因素的协调。另外,相关酶作用持续时间及峰值出现的时间还需要进一步观察分析。
4 结论
在0~15 mmol/L施氮范围内,苎麻蔗糖合成酶、β-1,3-葡聚糖酶和吲哚乙酸氧化酶活性随氮素水平提高呈现逐渐增加的趋势,并在超过9mmol/L后增幅显著下降。各酶活性在生长发育时期均呈单峰变化趋势,表现为旺长期>幼苗期>成熟期。氮高效基因型的相关酶活性均高于氮低效基因型,且SS和β-1,3-葡聚糖酶活性在幼苗期和旺长期达到显著水平,IAAO活性在旺长期和成熟期达到显著水平。相关酶对苎麻原麻产量具有重要影响,不同基因型对其响应特征不同,其中氮高效基因型的响应较敏感。苎麻纤维发育存在适氮水平,氮高效基因型可耐受更高氮素水平,并表现出显著较高的生产力和稳产性。
[1]李元沅,刘觉民.苎麻土壤生态系统的类型和组成[J].湖南农学院学报,1989,15(S1):60-69.
[2]王春桃,王良臣,潘昌立,等.氮素化肥对苎麻纤维产量和品质影响的研究[J].中国麻业科学,1982,4(4):1-5.
[3]周兆德,李天贵,欧阳铎声,等.苎麻肥料效应的研究[J].湖南农学院学报,1989,15(S1):76-82.
[4]欧阳铎声,周兆德,崔国贤,等.钾对苎麻碳氮代谢及纤维产量和品质的影响[J].中国麻业科学,1989,12(2):25-29.
[5]Meier H,Buchs L,Buchala A J,etal.(1,3)-β-D-Glucan(callose)is a probable intermediate in biosynthesis of cellulose of cotton fibres[J].Nature,1981,289(2):821-822.
[6]Brown R M,Saxena IM,Kudlicka K.Cellulose biosynthesis in higher[J].Trends in Plant Science,1996,5(1):149-155.
[7]Ruan Y,Chourey P S,Delmer D P,etal.The Differential Expression of Sucrose Synthase in Relation to Diverse Patterns of Carbon Partitioning in Developing Cotton Seed[J].Plant Physiology,1997,115(2):375-385.
[8]Delker C,Raschke A,QuintM.Auxin dynamics:the dazzling complexity of a smallmolecule’smessage[J].Planta,2008,227(5):929-941.
[9]Ranocha P,Dima O,Nagy R,et al.ArabidopsisWAT1 is a vacuolar auxin transport facilitator required for auxin homoeostasis.[J].Nature Communications,2013,4(4):2625.
[10]申素芳.苎麻纤维发育过程中纤维素合成关键酶与纤维品质的相关性研究[D].长沙:湖南农业大学,2014.
[11]陈平,喻春明,王延周,等.苎麻与大麻CesA1基因的生物信息学分析[J].中国麻业科学,2013,35(3):118-121.
[12]蒋杰,揭雨成,周清明,等.苎麻纤维素合酶基因BnCesA1全长cDNA的克隆与表达分析[J].植物遗传资源学报,2012,13(5):851-857.
[13]刘昱翔,陈建荣,彭彦,等.两种苎麻纤维素合酶基因cDNA序列的克隆及表达[J].作物学报,2014,40(11):1925-1935.
[14]田志坚,易蓉,陈建荣,等.苎麻纤维素合成酶基因cDNA的克隆及表达分析[J].作物学报,2008,34(1):76-83.
[15]汤涤洛.苎麻不同品种生根特性及氮素营养初步研究[D].武汉:华中农业大学,2012.
[16]中国科学院上海植物生理研究所.现代植物生理学实验指南[M].上海:科学出版社,1999.
[17]Salnikov V V,Grimson M J,Seagull RW,etal.Localization of sucrose synthase and callose in freeze-substituted secondarywall-stage cotton fibers[J].Protoplasma,2003,211(3):175-184.
[18]Amor Y,Haigler CH,Johnson S,et al.A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants[J].Proceedings of the National Academy of Sciences of USA,1995,92(20):9353-9357.
[19]Salnikov V V,Grimson M J,Seagull RW,etal.Localization of sucrose synthase and callose in freeze-substituted secondarywall-stage cotton fibers[J].Protoplasma,2003,211(3):175-184.
[20]马溶慧,许乃银,张传喜,等.氮素调控棉花纤维蔗糖代谢及纤维比强度的生理机制[J].作物学报,2008,34(12):2143-2151.
[21]蒋光华,孟亚利,陈兵林,等.低温对棉纤维比强度形成的生理机制影响[J].植物生态学报,2006,30(2):335-343.
[22]Arpat A,Waugh M,Sullivan JP,etal.Functional genomics of cell elongation in developing cotton fibers[J].PlantMolecular Biology,2004,54(6):911-929.
[23]高相彬,王友华,陈兵林,等.棉(Gossypium hirsutum L.)纤维发育相关酶和纤维比强度对棉铃对位叶氮浓度变化的响应研究[J].植物营养与肥料学报,2011,17(5):1227-1236.
[24]张文静,胡宏标,王友华,等.棉纤维发育相关酶活性的基因型差异与纤维比强度的关系[J].中国农业科学,2007,40(10):2177-2184.
[25]冯营,赵新华,王友华,等.棉纤维发育过程中糖代谢生理特征对氮素的响应及其与纤维比强度形成的关系[J].中国农业科学,2009,42(1):93-102.
[26]Jasdanwala R T,Sing Y D,Chinoy J J.Auxin Metabolism in Developing Cotton Hairs[J].Journal of Experimental Botany,1977,28(5):1111-1116.
[27]Yang Y M,Xu CN,Wang BM,etal.Effects of plantgrowth regulators on secondary wall thickening of cotton fibres[J].Plant Growth Regulation,2001,35(3):233-237.
Effects of Nitrogen Levels on Enzyme Activity Related to Fiber Development of Ramie Varieties with Different Nitrogen Utilization Efficiency
CHEN Jikang,TAN Longtao#,YU Chunming,ZHU Aiguo,CHEN Ping,GAO Gang,CHEN Kunmei,XIONG Heping*
(Institute of Bast Fiber Crops,Chinese Academy of Agricultural Sciences,Changsha 410205,China)
Two ramie varieties,H2000-03 with average nitrogen utilization efficiency is 46.01%and Ceheng Jiama,with average nitrogen utilization efficiency 15.81%,were used to perform potted experiment,with 5 nitrogen application levels(0 mmol/L,6 mmol/L,9 mmol/L,12 mmol/L,15 mmol/L),under glass in Changsha,Hunan Province.Enzyme activities related to fiber development were analyzed for understanding their physiologicalmechanism under differentnitrogen levels.The results showed that:Sucrose synthetase(SS),β-1,3-Glucanase and IAAO activities increased along with the increasing of nitrogen application.The increasing rate slowed down when nitrogen amount exceeded 9 mmol/L.Every enzyme activity showed a single-peak curve along with the growth periods of ramie and the sequence from high to low was fastgrowing period,seedling period andmaturing period.Enzymeactivities of H2000-03,which had a higher nitrogen utilization efficiency,was higher than Ceheng Jiama,especially for SS andβ-1,3-Glucanase at early period(seedling and fast growing period),for IAAO at later period(fast growing and maturing period)(p<0.05).Raw fiber yield of ramie was significantly affected by enzyme activities and showed different responding characteristics between varieties.Ramie variety with high nitrogen utilization efficiency wasmore sensitive to enzyme.There was an optimum range of nitrogen for fiber development of ramie.Ramie variety with high nitrogen utilization was better tolerated to higher nitrogen levels and represented higher productivity and yield stability.
ramie;nitrogen utilization efficiency;fiber development;enzyme;nitrogen levels
S563.1
A
1671-3532(2017)05-0257-07
2017-03-06
中国农业科学院基本科研业务费专项资助(1610242016027);国家麻类产业技术体系建设专项资助(CARS-19)
陈继康(1985-),男,助理研究员,主要从事苎麻多用途与农田生态研究。E-mail:chenjikang@caas.cn
#共同第一作者:谭龙涛(1984-),男,博士研究生,主要从事作物遗传育种研究。E-mail:3828134@163.com
*通讯作者:熊和平(1955-),男,研究员,主要从事苎麻育种研究。E-mail:ramiexhp@vip.163.com
