利用分子标记辅助选择聚合水稻Pi-ta、Pi-b和Wx-mq基因
2017-10-17张亚东赵庆勇周丽慧赵春芳王才林
姚 姝 陈 涛 张亚东 朱 镇 赵庆勇 周丽慧 赵 凌 赵春芳 王才林
江苏省农业科学院粮食作物研究所 / 江苏省优质水稻工程技术研究中心 / 国家水稻改良中心南京分中心, 江苏南京 210014
利用分子标记辅助选择聚合水稻Pi-ta、Pi-b和Wx-mq基因
姚 姝 陈 涛 张亚东 朱 镇 赵庆勇 周丽慧 赵 凌 赵春芳 王才林*
江苏省农业科学院粮食作物研究所 / 江苏省优质水稻工程技术研究中心 / 国家水稻改良中心南京分中心, 江苏南京 210014
近年来, 优良食味粳稻品种南粳 46、南粳5055和南粳 9108在江苏等地大面积推广, 促进了优质稻米产业的发展。但这些品种均不抗稻瘟病, 且缺乏适合在淮北地区种植的中熟中粳型优良食味粳稻品种。本研究以同时携带稻瘟病抗性基因Pi-ta和Pi-b的江苏抗病、高产粳稻品种武粳15为母本, 携带低直链淀粉含量基因Wx-mq的优良食味粳稻品种南粳5055为父本配置杂交组合进行聚合育种。利用Pi-ta和Pi-b基因的分子标记多重PCR体系以及Wx-mq基因的四引物扩增受阻突变体系PCR检测技术, 分别在不同的分离世代对目标基因位点进行检测, 结合田间多代选育、抗性鉴定和籽粒胚乳外观鉴定, 成功地将Pi-ta、Pi-b和Wx-mq基因聚合于一体, 选育出稻瘟病抗性好、食味品质优、产量高的水稻新品系“南粳0051”, 适合在江苏省淮北地区种植。本研究将三套自主研发的PCR检测体系成功应用于分子标记辅助选择, 不仅为水稻多基因聚合育种提供了快捷、高效的选择方法, 也为水稻抗病、优质育种创制了新的种质资源。
水稻; 稻瘟病; 低直链淀粉含量; Pi-ta; Pi-b; Wx-mq; 分子标记辅助选择
Abstract:In recent years, japonica rice varieties Nanjing 46, Nanjing 5055, and Nanjing 9108 were planted in a large area in Jiangsu province because of their good eating quality. However, these varieties are not resistant to rice blast, and there is no japonica rice variety with good eating quality and suitable for planting in Huaibei area, Jiangsu province. The goal of modern rice breeding is to develop new varieties with good quality, high yield and multi-resistance. Marker-assisted selection combined with traditional breeding methods can greatly improve breeding efficiency. In this study, the high yield rice variety “Wujing 15” contained two blast resistance genes of Pi-ta and Pi-b was used as gene recipient to be crossed with the rice variety “Nanjing 5055”,which carries the low-amylose content gene Wx-mq. We developed two multiple PCR systems to identify resistance and susceptible alleles on Pi-ta and Pi-b loci and the Tetra-primer Amplification Refractory Mutation PCR System for Wx-mq gene. MAS technique was adopted in combination with field phenotyping of agronomic traits, blast resistance identification, and classification of the grain endosperm appearance. Finally, a new japonica line, tentatively designated “Nanjing 0051”, was bred, which showed excellent agronomic traits and adaptation in the Huaibei area. The results indicate that the three PCR systems developed in this study are applicable and effective in rice breeding to improve blast resistance and grain quality. The success of breeding “Nanjing 0551” is an example for using MAS technique to pyramid favorable genes and facilitate breeding process in rice.
Keywords:Rice; Blast; Low-amylose content; Pi-ta; Pi-b; Wx-mq; Marker-assisted selection
稻瘟病由子囊真菌(Magnaporthe oryzae)引起,是影响水稻高产稳产最为严重的病害之一, 发病严重时甚至颗粒无收[1]。稻瘟病在水稻的各生育期均能发生, 但以穗颈瘟对水稻产量影响最大。稻瘟病广泛分布于世界各稻区[2]。20世纪90年代以来, 稻瘟病在我国西南、长江中下游和东北等稻区持续大流行, 年均发生面积330~570万公顷[3], 大暴发年份可达800万公顷左右, 直接威胁国家粮食安全。江苏是我国水稻主产省, 水稻种植面积 227万公顷左右,其中粳稻的种植面积约占全省水稻种植面积的90%;稻瘟病特别是穗颈瘟是影响江苏粳稻生产的主要病害之一。稻瘟病的防治一直是影响全世界水稻生产的大问题, 长期的实践证明, 将多个具有不同抗谱的稻瘟病抗性基因聚合到同一个品种中, 是培育具有持久抗瘟性品种的有效措施之一[4]。目前已经克隆的稻瘟病抗性基因中, 其主效抗性基因 Pi-ta和Pi-b在我国很多稻区表现广谱持久的稻瘟病抗性,被广泛应用于我国的水稻育种和生产[2,5-6]。
近年来, 随着市场经济的发展和人民生活水平的改善, 消费者对稻米食味品质的要求也越来越高。研究表明, 直链淀粉含量是决定米饭质地和食味品质的重要因素之一, 由颗粒结合淀粉合酶(granule-bound starch synthase, GBSS)催化合成,GBSS由蜡质基因 Wx编码[7]。低直链淀粉含量(5%~15%)的稻米是介于普通粳米和糯米之间的中间类型, 不仅米饭质地柔软、富有弹性, 而且具有冷不回生, 米饭适口性及膨化性好的特点, 越来越受到人们的青睐[8]。因此, 培育低直链淀粉含量的水稻品种, 不仅可以满足多样化的消费需求, 而且有利于开拓国内外稻米市场。目前已知水稻中可供育种利用的低直链淀粉含量突变基因有 14个, 它们大多由1对隐性基因控制, 且主要分布于水稻第6、第9、第10和第12染色体上[7]。其中Wx-mq是通过化学诱变剂 N-甲基-N-亚硝基脲(N-methyl-N nitrosourea, MNU)处理日本水稻品种越光而获得的低直链淀粉含量变异基因[9]。育种家利用携带Wx-mq基因的水稻资源Milky Queen培育了大量直链淀粉含量偏低、食味品质优良的水稻品种, 如New-hikari、关东 194、南粳 46[10]、南粳 5055[11]、南粳 9108[12]等。由于南粳 46、南粳 5055和南粳9108优异的食味品质, 符合当前农业供给侧结构性改革的需求, 受到广大农户的青睐, 种植面积正在逐步扩大。
2014年以来, 稻瘟病在江苏大范围爆发。南粳46、南粳5055和南粳9108只携带稻瘟病抗性基因Pi-b, 已不能满足当前江苏水稻生产对稻瘟病抗性的需求。因此, 选育抗稻瘟病、优质、高产水稻品种是当前江苏粳稻育种的主要目标。本研究利用稻瘟病抗性基因Pi-ta、Pi-b与感病等位基因pi-ta、pi-b多重PCR检测体系, 以及Wx-mq基因的四引物扩增受阻突变体系PCR检测技术, 在不同分离世代对目标基因进行检测, 结合田间多代选育、抗性鉴定和籽粒胚乳外观鉴定, 成功地将抗稻瘟病基因Pi-ta和Pi-b与低直链淀粉含量基因Wx-mq聚合于一体, 选育出多抗、优质、高产水稻新品系“南粳0051”。
1 材料与方法
l.1 材料
包括武粳15、南粳5055及其所配组合武粳15/南粳5055衍生的后代单株或株系, 其中, 武粳15为江苏省武进稻麦育种场育成的抗病、高产早熟晚粳稻品种, 携带稻瘟病抗性基因 Pi-ta和 Pi-b[13]; 南粳5055为江苏省农业科学院粮食作物研究所选育的优良食味粳稻品种, 含有来自关东194的低直链淀粉含量基因Wx-mq[8-11]。
1.2 DNA提取
在水稻分蘖盛期取新鲜幼嫩叶片, 参照 CTAB方法[14]提取供试材料单株基因组总DNA。
1.3 稻瘟病抗性基因Pi-ta和Pi-b及感病等位基因pi-ta和pi-b多重PCR检测体系
Pi-ta和Pi-b是最早被克隆的2个稻瘟病抗性基因, 对稻瘟病菌生理小种的抗谱较广[5-6]。虽然王忠华等[15]报道了检测稻瘟病抗性基因 Pi-ta和感病等位基因pi-ta, Fjellstrom等[16]和刘洋等[17]分别报道了检测稻瘟病抗性基因Pi-b和感病等位基因pi-b的引物序列, 但单个基因检测繁琐、费时、成本高, 不利于高通量检测。为此, 我们构建了同时检测稻瘟病抗性基因Pi-ta和Pi-b (简称体系I)、感病等位基因pi-ta和pi-b (简称体系II)的多重PCR体系[13]。体系I由引物YL155/YL87和Pi-bdomF/Pi-bdomR构成;体系II由引物YL183/YL87和Lys145F/Lys145R构成(表1)。两套多重PCR反应体系均为20 μL, 包括l×PCR buffer (含 1.5 mmol μL–1MgCl2)、200 μmol μL–1dNTP、1 μL Taq DNA 聚合酶(5 U μL–1)、100~150 ng模板 DNA和适量引物。扩增反应在Tprofessional热循环仪上进行, 经95℃预变性3 min;然后94℃变性1 min, 55℃复性1 min, 72℃延伸1 min, 6个循环; 再94℃变性1 min, 55℃复性50 s, 72℃延伸30 s, 32个循环; 最后72℃延伸6 min。反应产物在1.5%琼脂糖凝胶上120 V电泳25 min, 经溴化乙锭(Ethidium Bromide, EB)染色后于凝胶成像系统下观察, 记录结果。
1.4 Wx-mq基因四引物扩增受阻突变体系
采用Chen等[7]报道的四引物扩增受阻突变体系检测Wx-mq基因。引物名称、序列及其扩增片段预期大小见表2。20 μL PCR反应体系包括DNA (10 ng μL–1) 2.0 μL, Primer (4 pmol μL–1) 2.0 μL, 10×buffer(含 MgCl2) 2.0 μL, dNTP (2.5 mmol L–1) 0.4 μL, Taq DNA 聚合酶(5 U μL–1) 0.5 μL 和 ddH2O 13.1 μL。反应程序为95℃预变性5 min; 然后95℃变性30 s、65℃复性30 s、72℃延伸1 min, 循环35次后, 72℃延伸7 min, 10℃冷却10 min后, 将扩增产物加指示剂备用。反应产物在1.5%琼脂糖凝胶上120 V电泳25 min, 经溴化乙锭(EB)染色后并于凝胶成像系统下观察, 记录结果。本研究所用引物均由上海英潍捷基公司合成。

表1 用于多重PCR反应的引物名称、序列及预期片段长度Table 1 Name, sequences, and expected fragment size of specific primers used for multiplex PCR

表2 用于检测Wx-mq基因的引物名称、序列及预期片段长度Table 2 Name, sequences, and expected fragment size of specific primers used for detecting Wx-mq gene
1.5 稻瘟病抗性的接种鉴定
为鉴定新品系的稻瘟病抗性, 采用江苏省水稻品种中间试验的人工接种方法。在水稻孕穗破口期前5~7 d, 用6个注射器分别吸取6个稻瘟病生理小种的菌丝碎片与薄壁分生孢子混合液1 mL, 稻瘟病菌的分生孢子悬浮液, 浓度约为 2×105孢子 mL–1。从侧面注入水稻孕穗的苞内直至溢出。选择气温25~28℃和相对湿度为 90%的阴天或傍晚(当天下午的 15:00—18:00), 每个品系接种 6个生理小种, 对每个小种接种 5穗, 并作好相应标记, 每穗不能重复接种不同的生理小种。参照江苏省农业科学院植物保护研究所稻瘟病鉴定标准, 0级为无病, 免疫; 1级为1/4以下枝梗发病或穗颈有斑点, 抗病; 2级为1/4以上枝梗发病或主轴中部发病, 或颈部有病, 但对产量影响不大, 中抗; 3级为主轴中部或颈部发病,对产量有显著影响, 感病; 4级为穗颈发病造成白穗,高感[18]。
稻瘟病抗性鉴定接种菌株为江苏省南京、徐州、盐城、太湖、苏州等不同生态地区分离鉴定的稻瘟病菌中选择的各个生理小种代表性菌株, 包括ZB29、ZC15、ZD1、ZE3、ZF1和 ZG1, 均由江苏省农业科学院植物保护研究所提供。
1.6 胚乳淀粉特性的表型鉴定
成熟后对各单株籽粒胚乳淀粉基因型目测鉴定,胚乳外观全部为透明的个体可确认为正常胚乳的单株, 基因型 WxbWxb; 胚乳外观全部呈云雾状、乳白色、透明度差的个体可确认为低直链淀粉含量的单株, 基因型为 Wx-mqWx-mq; 胚乳外观既有透明的个体也有非透明的个体可确认为杂合单株, 基因型为WxbWx-mq。
2 结果与分析
2.1 利用分子标记辅助选择聚合 Pi-ta、Pi-b和Wx-mq
2008年正季在南京以武粳15为母本, 南粳5055为父本配制杂交组合, 同年冬季在海南种植 F1, 并混收F2种子。次年正季在南京种植F2分离群体。在分蘖盛期, 分别利用Wx-mq基因四引物扩增受阻突变体系 PCR检测技术和稻瘟病抗性基因 Pi-ta和Pi-b, 感病等位基因pi-ta和pi-b多重PCR检测体系对F2群体的285个单株进行分子检测。Wx-mq基因的扩增产物呈现3种类型的条带, 即南粳5055的带型(439 bp和292 bp)、武粳15的带型(439 bp和200 bp)以及同时具有双亲带型的杂合带型(439、292和200 bp)。经统计, 在 F2群体中共检测到具有南粳5055带型的单株72个, 具有武粳15带型的单株75个, 而具有杂合带型的单株 138株。这与成熟后对胚乳外观特性鉴定的结果完全一致。对具有439 bp和292 bp两条谱带的72个单株进行稻瘟抗性基因Pi-ta和Pi-b的多重PCR检测, 发现同时具有1024 bp和365 bp谱带的单株21个。随后利用感病等位基因pi-ta与pi-b多重PCR体系对上述72个单株验证检测显示, 相同编号的 21个单株中均未检测出1042 bp和803 bp谱带。证明这21个单株即为同时含有Pi-ta、Pi-b基因(纯合体)和Wx-mq基因(纯合体)的单株。部分PCR扩增结果如图1、图2和图3所示。

图1 武粳15/南粳5055衍生的F2群体中部分单株Wx-mq基因的分子检测Fig. 1 Detection of Wx-mq genotype in F2population derived from Wujing 15/Nanjing 5055

图2 武粳15/南京5055衍生F2群体中部分Wx-mq显性纯合体Pi-ta、Pi-b基因的分子检测Fig. 2 Detection of Pi-ta and Pi-b genes in F2population derived from Wujing 15/Nanjing 5055

图3 武粳15/南粳5055衍生的F2群体中部分Wx-mq显性纯合体pi-ta、pi-b基因的分子检测Fig. 3 Detection of pi-ta and pi-b genes in F2population derived from Wujing 15/Nanjing 5055
2.2 含Pi-ta、Pi-b和Wx-mq三基因纯合单株的表型鉴定
2010年正季将筛选到的 21个单株分别种成 F3家系进行表型鉴定。稻瘟病的田间抗性鉴定分别以亲本武粳15和南粳5055作为抗、感对照品种。采用来自江苏不同地区的 6个稻瘟病生理小种对不同株系进行穗颈瘟接种鉴定。接种后40~50 d, 黄熟初期, 分小区和生理小种收获接种的穗子, 调查总穗数以及病穗数, 按照发病百分率记载病级。其中, 南粳5055和武粳15的穗颈瘟综合抗性分别为4级和1级, 说明试验区稻瘟病的发病较为充分, 能有效区别F3家系在田间的抗性水平。鉴定结果表明, 21个单株衍生的F3家系中有7个家系的抗病性未发生分离, 发病率均在0~1级之间, 其中5个株系抗性水平与抗病亲本武粳15相当, 表现为抗; 2个株系对穗颈瘟表现为免疫, 抗性水平超过抗病对照武粳15 (表3)。证明利用分子标记进行水稻稻瘟病抗性的辅助选择是行之有效的。

表3 7个F2:3家系的稻瘟病抗性评价Table 3 Resistance to rice blast in seven F2:3lines
成熟后, 对每个家系分别收取50个单株观察水稻胚乳淀粉特性显示, 21个F4家系的所有单株胚乳外观全部呈云雾状、乳白色的不透明, 可确认其基因型为Wx-mqWx-mq。这与Wx-mq基因分子标记的检测结果完全吻合, 进一步证明利用四引物扩增受阻突变体系对暗胚乳突变基因Wx-mq进行辅助选择的可行性和准确性。
2.3 三基因聚合后代的农艺性状系统选育
根据综合农艺性状、田间抗性表现和产量性状,在每个F3家系选择15个单株, 7个F3家系中共选择105个单株, 2011年正季在南京种植105个F4小区,每个小区40株。成熟后在产量和农艺性状表现优异的 45个小区进行单株选择, 从每个小区选择 5~10株, 共选315个单株。中选单株于2012年正季在南京种成F5小区, 每个小区40株。其中有11个小区表现农艺性状稳定一致, 丰产性较好。2013年进行小区鉴定, 选择5个品系于2014年进行品比试验。根据试验结果, 其中 JD0051表现优异, 暂定名为“南粳0051”, 其选育过程如图4所示。为防止Wx-mq和 Pi-ta、Pi-b基因在选育、收获过程中丢失, 2014年在水稻分蘖盛期, 从 JD0051小区中随机选取 30个单株进行分子检测。其中, Wx-mq基因四引物扩增受阻突变体系的检测结果证实该品系同时具有纯合的低直链淀粉含量基因型(图 5); 稻瘟病抗性基因Pi-ta和Pi-b多重PCR检测体系的鉴定结果显示所有单株样品均出现约1024 bp和365 bp的特征条带,感病等位基因pi-ta和pi-b多重PCR检测体系的鉴定结果显示所有单株样品均未出现 1042 bp和 803 bp谱带, 这表明“南粳 0051”为具有 Pi-taPi-ta和Pi-bPi-b抗性基因的纯合体(图6和图7)。这一检测结果与田间对稻瘟病抗性的调查以及成熟后对胚乳外观形态的观察结果完全吻合。
2.4 抗病、优质新品系“南粳0051”的农艺性状
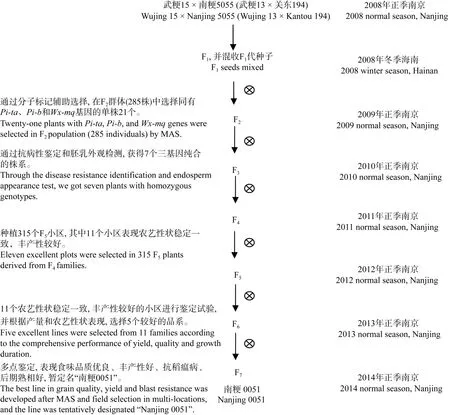
图4 分子标记辅助选择聚合3个基因的育种程序Fig. 4 Breeding procedure of pyramiding three genes by molecular marker-assisted selection (MAS)
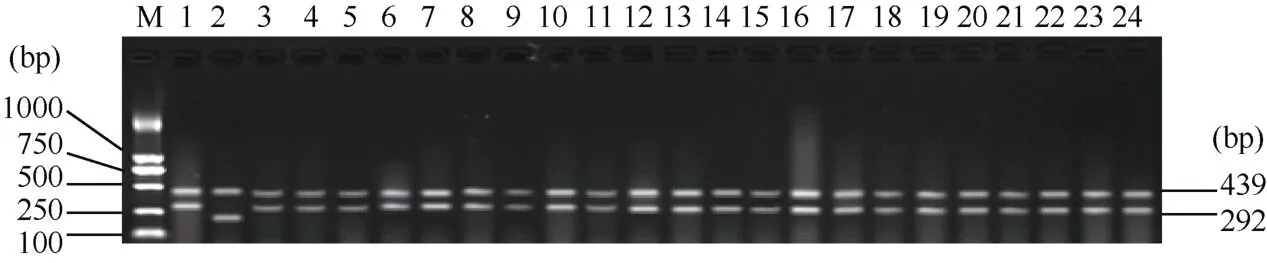
图5 稳定品系南粳0051 Wx-mq基因型的分子鉴定Fig. 5 Molecule identification of Wx-mq genotype in stable line Nanjing 0051

图6 稳定品系南粳0051抗病基因Pi-ta、Pi-b的分子鉴定Fig. 6 Molecule identification of Pi-ta, Pi-b genotype in stable line Nanjing 0051
在 2013—2014年的小区鉴定和多点试验中,“南粳0051”表现综合性状好, 全生育期150 d左右,与对照徐稻 3号相当, 属中熟中粳稻类型。株型集散适中, 株高 100 cm 左右; 分蘖性强, 单株成穗10~12个; 穗型较大, 每穗总粒数120~140粒, 结实率95%左右, 千粒重27~29 g, 米粒长宽比1.5左右。2015年和 2016年分别参加江苏省区域试验, 其中2015年区域试验产量10 238 kg hm–2, 分别比对照徐稻 3号(CK1)和连粳 7号(CK2)增产 9.9%和 4.5%,2016年区域试验产量10 173 kg hm–2, 比对照徐稻3号增产2.2% (表4)。江苏省农业科学院植物保护研究所统一进行抗性鉴定结果(表5), “南粳0051”苗瘟接种鉴定 6个生理小种 ZB7、ZC11、ZD5、ZE3、ZF1和ZG1均为0级, 穗颈瘟综合抗性指数3.5级左右, 抗性明显强于对照品种。农业部稻米及制品质量监督检验测试中心武汉分中心检测结果(表 6),“南粳 0051”的糙米率 85%左右, 精米率 74%左右,整精米率70%, 均高于对照品种, 垩白米率、垩白度则低于对照品种, 直链淀粉含量 10%左右, 胶稠度86~90 mm, 明显高于对照品种, 说明新品系的品质优于对照品种。“南粳 0051”由于含有低直链淀粉含量基因 Wx-mq, 与目前在江苏大面积生产上种植的南粳 46、南粳 5055、南粳 9108一样, 胚乳具半糯特性, 米饭晶莹剔透, 质地柔软, 口感润滑, 富有弹性, 食味品质较佳。

图7 稳定品系南粳0051感病基因pi-ta、pi-b的分子鉴定Fig. 7 Molecule identification of pi-ta, pi-b genotype in stable line Nanjing 0051

表4 新品系和对照的农艺性状和产量性状Table 4 Agronomic traits and yield traits of Nanjing 0051 and CK
3 讨论
3.1 分子标记辅助选择在基因聚合中的应用
优质、高产、多抗是水稻育种工作者长期追求的目标。而随着生产的发展、人民生活水平的提高以及气候与生态条件的变化, 对水稻育种提出了更新更高的要求。因此, 多基因聚合是育种工作者经常遇到的技术难题。近年来, 低直链淀粉含量的半糯粳稻南粳46、南粳5055、南粳9108等品种的优良食味品质得到江苏等地广大农户和居民的认可,种植面积正在逐步扩大。然而, 2014年以来, 由于稻瘟病在江苏等地的大范围爆发, 对这些品种的推广造成严重威胁。另一方面, 目前在江苏淮北地区还缺乏中熟中粳型优良食味粳稻品种。因此, 选育同时具备抗稻瘟病和低直链淀粉含量特性的中粳稻品种, 既能解决稻瘟病对当前水稻生产的严重威胁,又能满足人们对优质稻米日益增长的消费需求, 是江苏现代水稻育种的重要目标。在实际育种中, 如何在分离群体的大量个体植株中选择出目标单株是一项艰巨的工作, 利用分子标记辅助选择技术可在水稻生长的任何时期、取任何部位进行辅助选择,尤其是在抗病育种中人工接种病菌失败或没发病的情况下, 更能体现其优越性, 被育种家争相应用到水稻育种中。随着水稻基因组测序的完成, 越来越多的重要基因被定位或克隆, 分子标记辅助的基因聚合育种方法已经成为水稻分子育种的重要手段,是现代分子生物学与传统遗传育种的结合点。在多基因聚合过程中, 分子标记辅助选择具有非常高效的作用, 该技术能够快速准确地选到聚合 2个或 2个以上的目标基因的个体, 克服了常规表型鉴定的局限性。我们曾利用MAS技术聚合水稻抗条纹叶枯病基因 Stv-bi和低直链淀粉含量基因 Wx-mq[9];Huang等[19]和Hittalmani等[4]分别运用MAS技术聚合水稻抗白叶枯病和抗稻瘟病基因; Naoki等[20]将来自Modan的抗条纹叶枯病基因Stv-bi和抗稻瘟病基因 Pb1通过分子标记聚合, 选育出新的抗性品种爱知106; Narayanan等[21]聚合3个主效基因Pi-1、Piz-5和Xa21到Co39, 2个主效基因Piz-5和Xa21到IR50;Jiang等[22]聚合Xa21和Bt基因到恢复系明恢63中;倪大虎等[23]将抗稻瘟病基因 Pi9(t)及抗白叶枯病Xa21和 Xa23基因聚合到同一植株中, 获得了 4个三基因聚合且农艺性状优良的株系; 柳武革等[24]运用MAS技术聚合2个稻瘟病抗性基因, 培育广谱、持久抗稻瘟病两系不育系。

表6 南粳0051与对照品种的稻瘟病抗性比较Table 6 Comparison in grain quality traits between Nanjing 0051 and control varieties
3.2 抗稻瘟病基因Pi-ta、Pi-b和低直链淀粉含量基因Wx-mq的聚合
在抗性改良过程中, 回交选育可以有效地改善受体品种的个别性状, 但改良品种的综合性状与原品种性状相似, 很难在原有的基础上取得突破。为了进一步探讨多抗、优质基因聚合体创制及分子标记辅助选择的可行性, 本研究通过常规杂交与分子标记相结合, 利用抗稻瘟病基因Pi-ta和Pi-b, 感病等位基因pi-ta和pi-b多重PCR检测体系以及Wx-mq基因的四引物扩增受阻突变体系PCR检测技术, 为提高选择效率, 在整个基因聚合和转移过程中对每一代都实现了严格的筛选, 只选择优质和高抗稻瘟病的植株, 由于稻瘟病菌具有丰富的种群多样性,使所聚合的有限抗病基因选择压力逐年加大。因此利用分子标记辅助选择的同时必须利用人工接种稻瘟病菌检验。本研究利用田间抗性鉴定验证所选择标记的准确性, 更加注重农艺性状的选择, 从而实现了Pi-ta、Pi-b和Wx-mq三基因的聚合, 三基因聚合新品系“南粳 0051”高抗条纹叶枯病, 对稻瘟病的抗性明显强于目前生产上大面积种植的优良食味粳稻南粳46、南粳5055和南粳9108。
值得一提的是, 由于利用分子标记辅助选择进行多基因聚合, 需要利用与目的基因紧密连锁的分子标记对每个基因进行一次检测, 工作量很大; 此外, 还要对多基因聚合系进行抗性鉴定, 不同抗性基因在抗性鉴定上的差异, 会使工作量大增。种种限制因素, 造成目前多基因聚合育种还不如人意的现实。而本研究在检测稻瘟病抗性基因Pi-ta和Pi-b,感病等位基因pi-ta和pi-b上做了一些有益尝试, 并取得了良好的效果。我们通过不断优化PCR反应组分和反应条件, 构建了多重 PCR技术体系, 首次将2个稻瘟病抗性基因 Pi-ta和 Pi-b, 感病等位基因pi-ta和pi-b的检测在一次扩增体系中完成, 显著减少工作量, 节约成本, 提高育种效率, 特别适合育种单位对大批量材料进行分子标记辅助选择。
4 结论
利用分子标记辅助选择将稻瘟病抗性基因Pi-ta和Pi-b与低直链淀粉含量基因Wx-mq聚合于同一个体, 获得了稻瘟病抗性得到提高且食味品质优良的粳稻新品系“南粳 0051”, 说明本研究自主开发的三套分子标记选择体系对目标基因的辅助选择是有效可行的, 可用于优质抗病育种。而应用多重 PCR技术在同一反应体系中检测多个基因, 提高了检测和育种效率。
[1]王世维, 郑文静, 赵家铭, 魏松红, 王妍, 赵宝海, 刘志恒. 辽宁省稻瘟病菌无毒基因型鉴定及分析. 中国农业科学, 2014,47: 462–472 Wang S W, Zheng W J, Zhao J M, Wei S H, Wang Y, Zhao B H,Liu Z H. Identification and analysis of Magnaporthe oryzae avirulence genes in Liaoning province. Sci Agric Sin, 2014, 47:462–472 (in Chinese with English abstract)
[2]李祥晓, 王倩, 罗生香, 何云霞, 朱苓华, 周永力, 黎志康. 黑龙江省稻瘟病菌无毒基因分析及抗病种质资源筛选. 作物学报, 2012, 38: 2192–2197 Li X X, Wang Q, Luo S X, He Y X, Zhu L H, Zhou Y L, Li Z K.Analyzing avirulence genes of Magnaporthe oryzae from Heilongjiang province and screening rice germplasm with resistance to blast fungus. Acta Agron Sin, 2012, 38: 2192–2197 (in Chinese with English abstract)
[3]Shen M G, Lin J Y. The economic impact of rice blast disease in China. In: Zeigler R S, Leong S A, Teng P S. Rice Blast Disease.England, UK: CAB International, 1994. pp 321–331
[4]Hittalmani S, Parco A, Mew T W, Zeigler R S, Huang N. Fine mapping and DNA marker assisted pyramiding of the three major genes for blast resistance in rice. Theor Appl Genet, 2000, 100:1121–1128
[5]时克, 雷财林, 程治军, 许兴涛, 王久林, 万建民. 稻瘟病抗性基因Pi-ta和Pi-b在我国水稻主栽品种中的分布. 植物遗传资源学报, 2009, 10: 21–26 Shi K, Lei C L, Cheng Z J, Xu X T, Wang J L, Wan J M. Distribution of two blast resistance genes Pi-ta and Pi-b in major rice cultivars in China. J Plant Genet Resour, 2009, 10: 21–26 (in Chinese with English abstract)
[6]Bryan G T, Wu K S, Farrall L, Jia Y I, Hershey H P, McAdams S A, Fault K N, Donaldson G K, Renato T, Valent B. A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi-ta. Plant Cell, 2000, 12:2033–2045
[7]陈涛, 骆名瑞, 张亚东, 朱镇, 赵凌, 赵庆勇, 周丽慧, 姚姝,王才林. 利用四引物扩增受阻突变体系PCR技术检测水稻低直链淀粉含量基因Wx-mq. 中国水稻科学, 2013, 27: 529–534 Chen T, Luo M R, Zhang Y D, Zhu Z, Zhao L, Zhao Q Y, Zhou L H, Yao S, Wang C L. Detection of Wx-mq gene for low-amylose content by tetra-primer amplification refractory mutation system PCR in rice. Chin J Rice Sci, 2013, 27: 529–534(in Chinese with English abstract)
[8]朱昌兰, 沈文彪, 翟虎渠, 万建民. 水稻低直链淀粉含量基因育种利用的研究进展. 中国农业科学, 2004, 37: 81–88 Zhu C L, Shen W B, Zhai H Q, Wan J M. Progresses in researches of the application of low-amylose content rice gene for breeding. Sci Agric Sin, 2004, 37: 81–88 (in Chinese with English abstract)
[9]姚姝, 陈涛, 张亚东, 朱镇, 赵庆勇, 赵凌, 周丽慧, 王才林.利用分子标记辅助选择聚合水稻暗胚乳突变基因Wx-mq和抗条纹叶枯病基因Stv-bi. 中国水稻科学, 2010, 24: 341–347 Yao S, Chen T, Zhang Y D, Zhu Z, Zhao Q Y, Zhao L, Zhou L H,Wang C L. Pyramiding of translucent endosperm mutant Wx-mq gene and Stv-bifor resistance to rice stripe virus by Marker- assisted selection in rice (Oryza sativa L.). Chin J Rice Sci, 2010,24: 341–347 (in Chinese with English abstract)
[10]王才林, 张亚东, 朱镇, 陈涛, 赵凌, 林静. 通过分子标记辅助选择培育优良食味水稻新品种. 中国水稻科学, 2009, 23:25–30 Wang C L, Zhang Y D, Zhu Z, Chen T, Zhao L, Lin J. Breeding of a new rice variety with good eating quality by marker assisted selection. Chin J Rice Sci, 2009, 23: 25–30 (in Chinese with English abstract)
[11]王才林, 张亚东, 朱镇, 陈涛, 赵庆勇, 赵凌, 周丽慧, 姚姝.优良食味粳稻新品种南粳5055的选育及利用. 农业科技通讯,2012, (2): 84–87 Wang C L, Zhang Y D, Zhu Z, Chen T, Zhao Q Y, Zhao L, Zhou L H, Yao S. Breeding and application of a new rice variety Nanjing 5055 with good eating quality. Bull Agric Sci & Technol,2012, (2): 84–87 (in Chinese)
[12]王才林, 张亚东, 朱镇, 姚姝, 陈涛, 赵庆勇, 周丽慧, 赵凌.优良食味粳稻新品种南粳9108的选育与利用. 江苏农业科学,2013, 41(9): 86–88 Wang C L, Zhang Y D, Zhu Z, Yao S, Chen T, Zhao Q Y, Zhou L H, Zhao L. Breeding and application of a new rice variety Nanjing 9108 with good eating quality. Jiangsu Agric Sci, 2013,41(9): 86–88 (in Chinese)
[13]姚姝, 刘燕清, 张亚东, 朱镇, 陈涛, 赵庆勇, 周丽慧, 赵春芳,于新, 王才林. 水稻抗稻瘟病基因Pi-ta和Pi-b多重PCR体系的构建与应用. 作物学报, 2014, 40: 1565–1571 Yao S, Liu Y Q, Zhang Y D, Zhu Z, Chen T, Zhao Q Y, Zhao L,Zhou L H, Zhao C F, Yu X, Wang C L. Establishment and application of multiplex PCR system for two blast resistance genes Pi-ta and Pi-b in rice. Acta Agron Sin, 2014, 40: 1565–1571 (in Chinese with English abstract)
[14]Li W, Lei C L, Cheng Z J, Jia Y L, Huang D Y, Wang J L, Wang J K, Zhang X, Su N, Guo X P, Zhai H Q, Wan J M. Identification of SSR markers for a broad-spectrum blast resistance gene Pi20(t)for marker-assisted breeding. Mol Breed, 2008, 22: 141–149
[15]王忠华, 贾育林, 吴殿星, 夏英武. 水稻抗稻瘟病基因Pi-ta的分子标记辅助选择. 作物学报, 2004, 30: 1259–1265Wang Z H, Jia Y L, Wu D X, Xia Y W. Molecular markerassisted selection of the rice blast resistance gene Pi-ta. Acta Agron Sin, 2004, 30: 1259–1265 (in Chinese with English abstract)
[16]Fjellstrom R, Conaway-Bormans C A, McClung A M, Marchetti M A, Shank A R, Park W D. Development of DNA markers suitable for marker assisted selection of three Pi genes conferring resistance to multiple Pyricularia grlsea pathotypes. Crop Sci,2004, 44: 1790–1798
[17]刘洋, 徐培洲, 张红宇, 徐建第, 吴发强, 吴先军. 水稻抗稻瘟病 Pi-b基因的分子标记辅助选择与应用. 中国农业科学,2008, 41: 9–14 Liu Y, Xu P Z, Zhang H Y, Xu J D, Wu F Q, Wu X J.Marker-assisted selection and application of blast resistant gene Pi-b in rice. Sci Agric Sin, 2008, 41: 9–14 (in Chinese with English abstract)
[18]罗楚平, 倪磊, 陈志谊, 刘永锋, 刘邮洲, 聂亚锋. 水稻稻瘟病接种技术及2009年江苏省区试品种抗性鉴定. 江苏农业科学, 2009, (6): 178–179 Luo C P, Ni L, Chen Z Y, Liu Y F, Liu Y Z, Nie Y F. Inoculation technology and identification of resistance to rice blast in regional tests in 2009 of Jiangsu province. Jiangsu Agric Sci, 2009,(6): 178–179 (in Chinese)
[19]Huang N, Angeles E R, Domingo J, Magpantay G, Singh S,Zhang G, Kunaravdiel N, Bennett J, Khush G S. Pyramiding of bacterial blight resistance genes in rice: marker-assisted selection using RFLP and PCR. Theor Appl Genet, 1997, 95: 313–320
[20]Sugiura N, Tsuji T, Kato T, Saka N, Touyama T, Hayano S Y,Izawa T. Molecular marker assisted selection in a recurrent backcross breeding for the incorporation of resistance to rice stripe virus and panicle blast in rice (Oryza sativa L.). Breed Res,2004, 6: 143–148
[21]Narayanan N N, Baisakh N, Cruz C M V, Gnanamanickam S S,Datta K, Datta S K. Molecular breeding for the development of blast and bacterial blight resistance in rice cv. IR50. Crop Sci,2002, 42: 2072–2079
[22]Jiang G H, Xu C G, Tu J M, Li X H, He Y Q, Zhang Q F. Pyramiding of insect- and disease-resistance genes into an elite indica.cytoplasm male sterile restorer line of rice ‘Minghui 63’. Plant Breed, 2004, 123: 112–116
[23]倪大虎, 易成新, 李莉, 汪秀峰, 张毅, 赵开军, 王春连, 章琦,王文相, 杨剑波. 分子标记辅助培育水稻抗白叶枯病和稻瘟病三基因聚合系. 作物学报, 2008, 34: 100–105 Ni D H, Yi X C, Li L, Wang X F, Zhang Y, Zhao K J, Wang C L,Zhang Q, Wang W X, Yang J B. Developing rice lines resistant to bacterial blight and blast with Molecular marker-assisted selection. Acta Agron Sin, 2008, 34: 100–105 (in Chinese with English abstract)
[24]柳武革, 王丰, 金素娟, 朱小源, 李金华, 刘振荣, 廖亦龙, 朱满山, 黄慧君, 符福鸿, 刘宜柏. 利用分子标记辅助选择聚合Pi-1和 Pi-2基因改良两系不育瘟病. 作物学报, 2008, 34:1128–1136 Liu W G, Wang F, Jin S J, Zhu X Y, Li J H, Liu Z R, Liao Y L,Zhu M S, Huang H J, Fu F H, Liu Y B. Improvement of rice blast resistance in TGMS line by pyramiding of Pi-1 and Pi-2 through molecular marker-assisted selection. Acta Agron Sin, 2008, 34:1128–1136 (in Chinese with English abstract)
Pyramiding Pi-ta, Pi-b, and Wx-mq Genes by Marker-assisted Selection in Rice(Oryza sativa L.)
YAO Shu, CHEN Tao, ZHANG Ya-D*ong, ZHU Zhen, ZHAO Qing-Yong, ZHOU Li-Hui, ZHAO Ling,ZHAO Chun-Fang, and WANG Cai-Lin
Institute of Food Crops, Jiangsu Academy of Agricultural Sciences / Jiangsu High Quality Rice R&D Center / Nanjing Branch of China National Center for Rice Improvement, Nanjing 210014, China
10.3724/SP.J.1006.2017.01622
本研究由江苏省农业科技自主创新基金项目(CX[12]1003), 江苏省重点研发计划项目(BE2016370)和国家现代农业产业技术体系建设专项(CARS-01-62)资助。
This study was supported by the Agricultural Science and Technology Independent Innovation Fund of Jiangsu Province (CX [12]1003), the Key Research and Development Projects of Jiangsu Province (BE2016370), and China Agriculture Research System (CARS-01-62).
*通讯作者(Corresponding author): 王才林, E-mail: clwang@jaas.ac.cn, Tel: 025-84390307
): 2017-03-03; Accepted(接受日期): 2017-07-23; Published online(网络出版日期): 2017-08-02.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20170802.2059.002.html
