宝天曼自然保护区树附生苔藓空间格局分析
2017-09-25韦博良胡金涛贾宏汝叶永忠袁志良
许 宁,韦博良,胡金涛,贾宏汝,叶永忠,袁志良
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.河南财政金融学院,河南 郑州 450052)
宝天曼自然保护区树附生苔藓空间格局分析
许 宁1,韦博良1,胡金涛1,贾宏汝2,叶永忠1,袁志良1
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.河南财政金融学院,河南 郑州 450052)
采用单变量g(r)函数,选用同质性泊松、异质性泊松、同质性托马斯和异质性托马斯4个零模型,分析了宝天曼自然保护区内1 hm2样地中11种树附生苔藓物种的空间分布格局。结果显示,11个物种中,有6个物种在同质性泊松模型下表现出聚集分布,在同质性托马斯模型下的聚集程度有所减弱,但在异质性泊松模型下完全变为随机分布。与同质性泊松模型下的分布相比,在异质性泊松模型下,有4个物种在较大尺度上的聚集分布变为随机分布;而在同质性托马斯模型下,有2个物种在较小尺度上的聚集分布变为随机分布。研究表明: 1)生境异质性和扩散限制均对树附生苔藓空间分布格局的形成有影响,且生境异质性起主要作用; 2)生境异质性可在小尺度上影响树附生苔藓的空间分布,扩散限制也能在大尺度上发挥作用。
树附生苔藓;空间格局;零模型;生境异质性;扩散限制
苔藓植物是水生向陆生过渡的高等植物,在世界各地从热带雨林至寒温带荒漠包括南极洲在内均有分布[1]。在生态系统中,苔藓植物是独特而又不可缺少的一个类群。树附生苔藓植物(epiphytic bryophytes)是生活在乔木或者灌木树皮上的一类苔藓植物[2]。由于这类苔藓植物生活环境和附生基质的特殊性,其对环境变化的响应较其他植物更为敏感[3],因此有关苔藓的研究多用树附生苔藓作为研究对象。国内外对树附生苔藓植物的研究主要集中在区系、环境监测和多样性方面[4-5],对构建树附生苔藓植物种群空间格局驱动力的研究才刚刚起步。点格局分析方法能描述任意尺度下种群的空间分布特点,是有效的格局分析方法[6]。空间格局形成的主要驱动力是基于生态位理论的生境异质性[7]和基于中性理论的扩散限制[8]。生境异质性强烈影响着群落物种的空间分布[9-10],而传播能力通常与空间资源量的占有能力成正比,从而肯定了扩散限制对群落结构构建的重要贡献[11]。以往点格局的研究多以树木作为研究对象,有关苔藓的相关研究还较少,苔藓生长是否受到生境异质性和扩散限制影响还不得而知。因此,本研究以1 hm2样地内所有树附生苔藓植物为研究对象,应用点格局分析方法对树附生苔藓空间格局进行研究,进而揭示影响树附生苔藓格局形成的生态学过程。
1 研究区概况
河南省宝天曼自然保护区位于河南南阳市内乡县, 地理坐标为33°25′~ 33°33′ N, 111°53′~ 112°04′ E,属于典型的北亚热带向暖温带过渡地区[12],其植被属暖温带落叶阔叶林向亚热带常绿阔叶林的过渡类型。保护区总面积约为53.4 km2, 年平均气温15.1 ℃,1月份平均气温1.5 ℃,7月份平均气温27.8 ℃。高山区无霜期160 d,低山区无霜期227 d。年均降水量885.6 mm,年均蒸发量991.6 mm,年均相对湿度为68%[13]。
2 研究方法
2.1样地调查
2016年6月,在宝天曼保护区核心地带选取苔藓植物类型具有代表性的地段设立一个100 m×100 m样地,并对样地内所有胸径大于1 cm的目标树干上的附生苔藓植物进行普采,共采集树生苔藓植物标本230份,全部在实验室鉴定到种,凭证标本保存于河南农业大学标本馆。
2.2数据处理
本研究中树附生苔藓植物的位置信息来自被附生的木本或灌木植物的x和y坐标,也就是每个物种在1 hm2样地中出现的点位(以下简称为附生点位)。在单个物种格局分析中要求个体数不小于30,因此,本研究选取符合该条件的11个物种进行格局分析。
本研究采用单变量g(r)函数[14]来分析树附生苔藓植物空间分布格局,并量化各空间分布格局发生的尺度。g(r)函数由Ripley’sK函数衍生而来, 因其能有效地剔除Ripley’sK函数不断累积的小尺度效应[15-16],在目前的格局分析中得到了广泛的应用。
生态学研究的一个重要方法是通过物种空间分布格局来推导其形成过程,进而推测多样性的维持机制[17]。本研究采用的零模型包括同质性泊松模型(homogenous Poisson model)、异质性泊松模型(heterpgeneous Poisson model)、同质性托马斯模型(homogenous Thomas model)和异质性托马斯模型(heterpgeneous Thomas model)。由于空间格局形成过程的复杂性,单一的零模型无法准确论证繁复的生态过程。同质性泊松模型是点格局过程中最简单的一种,假设物种的空间格局分布不受任何过程影响,其实质为完全空间随机模型(CSR)[18]。异质性泊松模型主要通过空间异质强度函数表现物种个体密度与生境异质性的关联[19],可模拟物种与其生境之间的相互关系[20],作为检验生境异质性是否影响物种空间分布格局时的零假设[19]。同质性托马斯模型模拟的是物种子代个体受扩散限制而聚集分布于母株周围的分布格局,作为检验物种空间分布格局是否受扩散限制的影响的零假设。异质性托马斯模型同时模拟了生境异质性和扩散限制的生态学过程,主要用于估计生境异质性与扩散限制联合效应。
通过以上4种模型的比较,可以在一定程度上明确生境异质性和扩散限制对不同树附生苔藓植物物种空间格局形成的影响作用[21]。例如某物种在同质性泊松模型下有聚集分布,而在异质性泊松模型下的所有尺度上均为随机分布,说明生境异质性影响了该物种空间格局的形成;如果某物种在同质性泊松模型下有聚集分布,而在同质性托马斯模型下聚集程度有所减弱,可说明传播限制作用是物种空间格局构建过程中的驱动力。
格局分析通过R-3.3.0软件中的Spatstat软件包实现。通过99次Monte Carlo随机模拟计算99%的置信区间,g(r)值高于上包迹线为聚集分布, 在上下包迹线之间为随机分布,低于下包迹线为均匀分布。
3 结果与分析
3.1苔藓物种组成
根据标本鉴定结果,样地内树附生苔藓植物共有20科39属59种(表1),其中苔类4科4属5种,藓类16科35属54种。

表1 样地中分布的59种树附生苔藓植物名录Table 1 List of 59 epiphytic bryophyte species in sample area
附生点位数不小于30的树附生苔藓有羊角藓、短叶毛锦藓、瓦叶假细罗藓、长肋青藓、皱叶牛舌藓、皱叶绢藓、长叶纽藓、毛尖青藓、中华耳叶苔、黄边凤尾藓和圆叶匐灯藓等11个物种。它们的附生点位数依次为138、69、69、52、44、42、35、33、33、30和30,总计575,占样地内所有树附生苔藓点位总数的66.63%。
3.2空间分布格局
3.2.1 空间分布点图 样地中11个树附生苔藓空间点位分布见图1。从图1中也可以看出这11个物种在样地中的密度和空间分布位置有显著差异,其中羊角藓在样地北侧的密集程度最高,而且密度呈现出从南到北逐渐增大的趋势(图1-a);短叶毛锦藓在样地南侧有明显的聚集(图1-b);瓦叶假细罗藓和长肋青藓的空间点位均呈现出小聚块分布(图1-d、图1-f);皱叶牛舌藓和皱叶绢藓主要分布在样地北侧部分(图1-i、图1-k)。其他树附生苔藓物种在样地中的分布较广,无明显的分布趋势。
3.2.2 格局分析结果 不同树附生苔藓在4个模型下的空间分布格局有显著差异。如短叶毛锦藓在同质性泊松分布模型下表现为聚集分布,在同质性托马斯分布模型下聚集程度有所减弱,在异质性泊松分布模型和异质性托马斯分布模型下表现为随机分布(图2)。
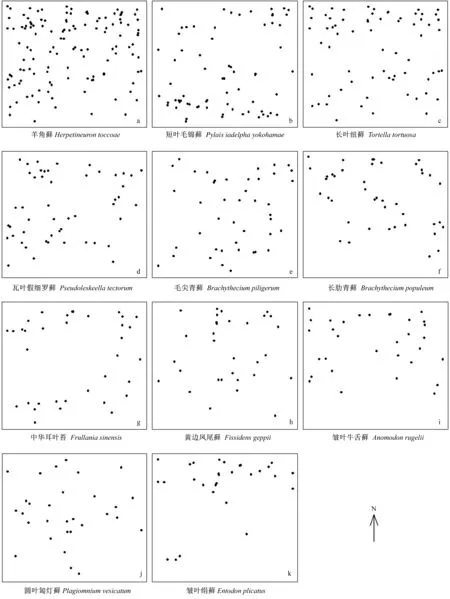
图1 11个苔藓物种在样地中的分布点图Fig.1 Mapped point pattern of 11 bryophyte species
从11个树附生苔藓在不同模型下的空间格局来看(表2),在同质性泊松分布模型下空间格局表现出聚集的树附生苔藓物种一共有6种,分别为羊角藓、短叶毛锦藓、瓦叶假细罗藓、长肋青藓、皱叶牛舌藓和皱叶绢藓。与同质性泊松分布模型相比,在同质性托马斯分布模型下空间格局表现出聚集的树附生苔藓物种减少为4个,分别为短叶毛锦藓、瓦叶假细罗藓、皱叶牛舌藓和皱叶绢藓。在异质性泊松分布模型和异质性托马斯分布模型下,所有11个树附生苔藓物种在所有尺度上均呈现出随机的分布格局。

图中黑色实线为用实际数值计算的g(r),虚线为理论值,灰色区域为置信区间,上下边界分别为拟合的上、下包迹线。
The black solid line in the figure is calculated with the actual value of theg(r); The dotted line is theoretical value; The gray area is confidence interval; The upper and lower boundaries are respectively fitting upper and lower envelope.

图2 短叶毛锦藓格局分析结果Fig.2 Point pattern analyses for Pylaisiadelpha yokohamae

续表Continued Table m
注:表中“—”表示物种在该模型下的所有尺度上均为随机分布格局;聚集尺度小于1 m的忽略不计。
Note: The “—” indicates that the specie are randomly distributed on all scales in the model; Ignore the aggregation scale that smaller than 1 m.
4 结论与讨论
空间分布格局是植物种群个体在群落内的分布形式,可反映出种群与生态因素之间的内在联系。聚集分布格局产生的主要原因是扩散限制、生境异质性以及二者的共同作用[22]。羊角藓和长肋青藓在模拟生境异质性的同质性泊松分布模型下均有聚集分布,在去除环境异质性效应的异质性泊松模型下全部尺度上都为随机分布,表明这两个物种的空间格局形成均受到生境异质性影响。且相对同质性泊松模型而言,这两个物种在剔除扩散限制的同质性托马斯模型下的聚集分布完全消失,说明生境异质性和扩散限制都是形成这两种苔藓植物整体空间分布格局的驱动力。
短叶毛锦藓、瓦叶假细罗藓、皱叶牛舌藓和皱叶绢藓这4个物种在同质性泊松分布模型有聚集分布,但在同质性托马斯分布模型下的聚集程度弱于同质性泊松分布模型,这表明扩散限制在一定程度上影响了这4个物种空间格局的形成。同时,这4个物种在异质性泊松模型下全部尺度上都为随机分布,这表明生境异质性对这几个物种的分布也有影响,并且所起的作用比扩散限制的作用更大。
近年来的研究表明,扩散限制和生境关联通常共同作用于植物群落的组配[23-24]。从总体上来看,在研究的11个树附生苔藓物种中,有6个树附生苔藓物种空间格局形成均受两种驱动力的影响,且生境异质性对树附生苔藓空间分布的影响较大。其余的5个物种:长叶纽藓、毛尖青藓、中华耳叶苔、黄边凤尾藓和圆叶匐灯藓的空间格局在4个模型下均无聚集分布。其原因可能是由于本研究中所设立样地位置没有涵盖到适合这几个物种的最适生境[25],或者是样地面积有限,而这几种树附生苔藓物种孢子的传播距离超过了样地的最大尺度,使得这几个树附生苔藓在1 hm2样地内没有呈现出显著的聚集分布。因此也无法检测扩散限制和生境异质性是否影响这几个树附生苔藓的格局形成。
有研究表明,扩散限制和生境异质性对格局形成的影响与研究尺度有关。不同物种空间格局的形成受到不同生态过程的影响,而且不同生态过程发生显著作用的空间尺度也存在差异。对乔木而言,生境异质性对空间格局的影响通常发生在较大尺度上(大于10 m)[26]。本研究中短叶毛锦藓在同质性泊松模型与异质性泊松模型下的分布格局表明,生境异质性不仅在大尺度上起作用,也在小尺度上(小于10 m)对树附生苔藓空间格局形成过程中起作用。与乔木不同的原因可能是树附生苔藓本身形体微小,而且对环境变化敏感,因此,树附生苔藓受小尺度上环境异质性的影响较乔木更为剧烈。STOKES等[27]认为扩散限制是解释物种在较大尺度上聚集的原因之一。瓦叶假细罗藓、皱叶牛舌藓和皱叶绢藓在同质性泊松模型与同质性托马斯模型下的聚集尺度的变化说明,扩散限制可以在较大尺度上影响树附生苔藓植物的空间分布格局,这表明苔藓植物孢子可以在媒介作用下长距离传播。
探究环境因子和扩散限制在群落空间格局构建中的相对重要性是植物学研究的重要课题。苔藓植物种群空间分布格局的形成是物种本身、物种之间的生态关系以及环境因子共同作用的结果。扩散限制和生境异质性在树附生苔藓植物群落的形成过程中都起着重要作用。树附生苔藓能够扩散到较大尺度上,而且在较小尺度上也受到生境异质性的影响。影响物种空间分布的原因有很多,本研究仅分析了环境因子和扩散限制对树附生苔藓的影响,未来将进一步研究苔藓的种间关系,并分析其他生活型物种对苔藓的生长和孢子传播的影响。
[1] 吴鹏程. 苔藓植物生物学[M]. 北京: 科学出版社, 1998.
[2] SMITH A J E. Bryophyte ecology[M]. London: Chapman and Hall, 1982.
[3] 徐晟翀, 曹同, 于晶,等. 上海市树附生苔藓植物生态位[J]. 生态学杂志, 2006, 25(11): 1338-1343.
[4] MITCHELL R J, SUTTON M A, TRUSCOTT A M, et al. Growth and tissue nitrogen of epiphytic Atlantic bryophytes: effects of increased and decreased atmospheric N deposition[J]. Functional Ecology, 2004, 18(3): 322-329.
[5] DAVIS D D, MCCLENAHEN J R, HUTNIK R J. Use of an epiphytic moss to biomonitor pollutant levels in southwestern Pennsylvania[J]. Northeastern Naturalist, 2001, 8(4): 379-392.
[6] 张金屯. 植物种群空间分布的格局分析[J]. 植物生态学报, 1998, 22(4): 344-349.
[7] KYLE E H, CONDIT R, HUBBELL S P, et al. Habitat associations of trees and shrubs in a 50 hm2neotropical forest plot[J]. Journal of Ecology, 2001, 89(6): 947-959.
[8] HUBBELL S P, BORDA-DE-GUA L. The unified neutral theory of biodiversity and biogeography [M]. New Jersey: Princeton University Press, 2001.
[9] CHEN L, MI X, COMITA L S, et al. Community-level consequences of density dependence and habitat association in a subtropical broad-leaved forest[J]. Ecology Letters, 2010, 13(6): 695-704.
[10] BARTELS S F, CHEN H Y H. Is understory plant species diversity driven by resource quantity or resource heterogeneity?[J]. Ecology, 2010, 91(7): 1931-1938.
[11] SEIDLER T G, PLOTKIN J B. Seed dispersal and spatial pattern in tropical trees[J]. PLoS Biol, 2006, 4(11): e344.
[12] 尚富德, 王正德. 伏牛山南北自然过渡地带植物多样性的特征及其成因分析[J]. 河南大学学报(自然科学版), 1998, 28(1): 54-60.
[13] 陈云, 袁志良, 任思远,等. 宝天曼自然保护区不同生活型物种与土壤相关性分析[J]. 科学通报, 2014, 59(24): 2367-2376.
[14] RIPLEY B D. Spatial Statistics[M]. New Jersey: John Wiley & Sons Inc, 2005.
[15] WIEGAND T, GUNATILLEKE S, GUNATILLEKE N. Species associations in a heterogeneous Sri Lankan dipterocarp forest[J]. American Naturalist, 2007, 170(4):77-95.
[16] STOYAN D, PENTTINEN A. Recent applications of point process methods in forestry statistics[J]. Statistical Science, 2000, 15(1): 61-78.
[17] LAW R, ILLIAN J, BURSLEM D F R P, et al. Ecological information from spatial patterns of plants: insights from point process theory[J]. Journal of Ecology, 2009, 97(4):616-628.
[18] 王鑫厅, 侯亚丽, 梁存柱,等. 基于不同零模型的格局分析[J]. 生物多样性, 2012, 20(2): 151-158.
[19] ILLIAN J, PENTTINEN A, STOYAN H, et al. Spatial analysis and modelling of spatial point patterns[J]. Technometrics, 2008, 47(4): 516-517.
[20] WIEGAND T A. MOLONEY K. Rings, circles, and null-models for point pattern analysis in ecology[J]. Oikos, 2004, 104(2): 209-229.
[21] DIGGLE P J. Statistical analysis of spatial point patterns[J]. Biometrics, 1983, 32(3): 659-667.
[22] LIN Y C, CHANG L W, YANG K C, et al. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation[J]. Oecologia, 2011, 165(1): 175-184.
[23] SHEN G, YU M, HU X S, et al. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity [J]. Ecology, 2009, 90(11): 3033-3041.
[24] WANG X, WIEGAND T, WOLF A, et al. Spatial patterns of tree species richness in two temperate forests[J]. Journal of Ecology, 2011, 99(6): 1382-1393.
[25] 黄运峰, 丁易, 臧润国,等. 海南岛霸王岭热带低地雨林树木的空间格局[J]. 植物生态学报, 2012, 36(4): 269-280.
[26] VALENCIA R, FOSTER R B, VILLA G, et al. Tree species distributions and local habitat variation in the Amazon: large forest plot in eastern Ecuador[J]. Journal of Ecology, 2004, 92(2): 214-229.
[27] STOKES C J, ARCHER S R. Niche differentiation and neutral theory: an integrated perspective on shrub assemblages in a parkland savanna[J]. Ecology, 2010, 91(4):1152-1162.
(责任编辑:朱秀英)
SpatialpatternanalysisforepiphyticbryophytesinBaotianmanNatureReserve
XU Ning1, WEI Boliang1, HU Jintao1, JIA Hongru2, YE Yongzhong1, YUAN Zhiliang1
(1.College of Life Sciences, Henan Agricultural University, Zhengzhou 450002, China; 2.Henan College of Finance and Taxation, Zhengzhou 450002, China)
In this study, we analysed the spatial pattern of 11 epiphytic bryophytes species by using univariate ofg(r) function, along with homogeneous Poisson model,heterogeneous Poisson model, homogeneous Thomas model and heterogeneous Thomas model, in the 100 m×100 m plot area of Baotianman Nature Reserve. The results showed that: 1) 6 out of 11 species showed aggregation distribution in the homogeneous Poisson model, and the aggregation decreased under the homogeneous Thomas model. But all of them were randomly distributed in the heterogeneous Poisson model; 2) Compared with the distribution under the homogeneous Poisson model, 4 species’ aggregation distribution turned into random distribution on the large scale of the heterogeneous Poisson model; And 2 species’ aggregation distribution developed into random distribution on a small scale of homogeneous Thomas model.The research suggests that: 1) Both habitat heterogeneity and dispersal limitation have effects on the spatial distribution pattern of epiphytic bryophytes, and the habitat heterogeneity plays a major role; 2) Habitat heterogeneity can affect the spatial distribution of epiphytic bryophytes on a small scale, and dispersal limitation can also work on a large scale.
epiphytic bryophytes; spatial pattern; null model; habitat heterogeneity; dispersal limitation
S 718
:A
2016-11-08
河南省科技攻关项目(132102110133);河南省教育厅科学技术研究重点项目(14A180013)
许 宁(1991-),男,山西运城人,硕士研究生,主要从事植物生态研究。
袁志良(1976 -),男,河南信阳人,副教授,博士。
1000-2340(2017)02-0230-07
