宝天曼不同生长基质上苔藓植物的多样性
2017-09-25李培坤胡金涛杜晓军叶永忠袁志良
张 旭,李培坤,胡金涛,杜晓军,叶永忠,袁志良
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.中国科学院植物研究所植被数量生态学重点实验室,北京 100093)
宝天曼不同生长基质上苔藓植物的多样性
张 旭1,李培坤1,胡金涛1,杜晓军2,叶永忠1,袁志良1
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.中国科学院植物研究所植被数量生态学重点实验室,北京 100093)
基于宝天曼暖温带-北亚热带落叶阔叶林1 hm2样地,通过重要值、多样性指数和相似性指数等指标对样地内不同附生基质上苔藓多样性进行研究。结果表明,样地内苔藓植物共有29科57属98种,羊角藓为优势种。样地内石生苔藓物种数最多,共有77种;石生苔藓、土生苔藓和树附生苔藓的多样性指数依次减小。石生苔藓与土生苔藓的相似性系数最大,为0.722,而土生与腐木生苔藓物种相似性最低,为0.523。苔藓生活型以耐旱能力最弱的平铺型为主,反映出样地内水分状况良好,但在不同生长基质中苔藓生活型的比例存在差异。由此可见,不同苔藓对不同生长基质的适应存在一定差异,生境的多样性有助于提高苔藓的多样性。
苔藓; 生活型;生物多样性;生长基质; 宝天曼
苔藓是由水生向陆生过渡的高等植物,具有独特的生理生态适应机制[1],常常成为各种生长基质上的先锋拓荒种[2]。在森林生态系统中,苔藓作为重要组成成分,对森林物种多样性的维系发挥着重要的作用[3],但不同生长基质上的苔藓却在物种组成和丰富度上不尽相同[4]。曹同等[5]在对长白山地区地面生苔藓、树附生苔藓和腐木生苔藓的多样性研究中发现生境类型的异质性影响苔藓分布和组成;马文章等[6]对哀牢山中山湿性常绿阔叶林木质残体、林地、林冠和岩石上的苔藓进行了研究,结果显示不同基质上苔藓物种组成差异较大,优势苔藓植物间的关联程度影响苔藓植物的多样性。因此,针对不同生长基质上的苔藓植物展开调查和分析,深入研究山地森林生态系统苔藓植物物种多样性维持机制至关重要。苔藓的生活型是指在其生长型的基础上,包含环境条件对植物形态塑造的信息[7]。生活型涵盖苔藓生长点的位置和分枝类型等形态特征,而且还包括植株对生境的拓展策略等生态信息。相同类别生活型的苔藓具有相似的生态习性和竞争策略,对生活型的划分有助于物种鉴定,易于分析苔藓的分布及苔藓群丛的构建成因[6]。以往虽然有大量学者对苔藓的物种组成以及对生活型进行了分析[8-10],但很少有比较研究不同生长基质间苔藓的物种组成的相似性以及多样性差异,更少有研究检验不同生活型的苔藓与其附生基质的关系。为此,本研究以宝天曼自然保护区1 hm2样地内的苔藓为研究对象,对苔藓的物种组成和物种多样性进行调查研究,以期深入理解暖温带-亚热带过渡区物种多样性的维持机制,并对宝天曼自然保护区内物种的多样性保护提供参考。
1 研究样地与研究方法
1.1研究区域概况及样地设置
宝天曼国家级自然保护区位于河南省西南部,为暖温带-亚热带气候过渡区,年均温度15.2 ℃,年均降雨量885.6 mm,年均相对湿度68%,土壤pH值为6.5。优越的气候环境条件孕育了本区茂密的森林、丰富的野生动、植物资源,为永久性大型综合监测研究提供了优越条件[11-12]。
参照BCI 50 hm2样地的建立方法[13],在宝天曼自然保护区建立了1个1 hm2(100 m×100 m)的落叶阔叶林样地(森林类型主要为锐齿栎林森林群落),将样地划分为25个20 m×20 m的样方并编号,对每个样方内胸径(DBH)≥1 cm的个体进行每木调查并编号。
1.2样方调查
在苔藓生态学研究中,对苔藓进行调查时多根据一定的研究目的设置一定的样线和样方,统计20 cm×20 cm样方内物种盖度[8-10]。但对于苔藓在大面积和大尺度上的研究较少。为获得苔藓在较大尺度上的生态学特性,本研究对样地内的苔藓进行全面普查,参考文献[6]对苔藓生长基质的划分,将苔藓生长基质划分为树干、土壤、岩石和木质残体(Coarse Woody Debris, CWD),并将样地内各生长基质上的每一块苔藓群丛作为一个样点,用小方格为1 cm×1 cm的金属网调查样点内的苔藓的面积,采集样点内所有苔藓物种,并在标本袋上记录各物种在金属丝框内的面积、生长基质、与所附生的树干编号和样地编号。采集的苔藓标本依据《苔藓植物志》和《中国苔纲和角苔纲植物属志》在实验室显微镜下鉴定。
1.3数据分析
1.3.1 重要值计算 由于本研究是大尺度的调查取样,计算重要值时参照草本重要值的计算方法,将苔藓在20 m×20 m样方上出现的次数除以总的样方数作为频度;将苔藓在所有样点内出现的次数作为该物种的多度;将某种苔藓在所有样点中的面积除以所有样点的总面积作为该物种的显著度。苔藓的重要值计算公式如下:
样地内某物种重要值=1/3(相对频度+相对多度+相对显著度)
相对频度=某种苔藓的频度/所有苔藓的频度之和
相对多度=某种苔藓的多度/所有苔藓的多度之和
相对显著度=某种苔藓的显著度/所有苔藓的显著度之和
1.3.2 多样性指数 为定量比较不同生态系统苔藓的多样性和物种丰富度,用重要值代替“物种个体数量”计算物种多样性指数[14],对树附生苔藓、土生苔藓、石生苔藓的多样性指标进行计算,以反映生态群落中物种的丰富度和多样性。公式如下:
Shannon-wiener多样性指数H′=-∑PilnPi
(1)
(2)
式中:Pi为物种i的相对重要值,Pi=Ni/N,Ni为物种i的绝对重要值,N为重要值之和。
1.3.3 相似性系数 相似性系数的研究对分析不同生长基质间苔藓物种的组成分布和了解苔藓对环境的适应能力具有重要意义。本研究采用Sφrensen指数计算不同生长基质上苔藓物种的相似性[15]:
Cs=2a/(2a+b+c)
(3)
式中:Cs是Sφrensen相似性系数,a为甲乙2个群落所共有的物种数量,b为甲群落有而乙群落没有的物种数,c为乙群落有而甲群落没有的物种数。
1.3.4 苔藓生活型划分 依据 BATES对苔藓生活型的定义[7],参考马文章等[8]针对哀牢山苔藓生活型的划分,本研究按照表1的标准对宝天曼国家自然保护区的苔藓进行了划分,共划分为5个生活型。

表1 附生苔藓生活型的划分依据Table 1 Classification of life forms in epiphytic bryophytes
2 结果与分析
2.1不同生长基质苔藓概况
2016年6月对宝天曼1 hm2样地中的苔藓进行调查,共采集苔藓样本764份,其中树生苔藓254份,土生苔藓113份,石生苔藓319份,腐木生苔藓78份。鉴定结果如表2,经鉴定发现共采集苔藓29科57属98种,树附生苔藓21科40属62种,土生苔藓21科36属56种,石生苔藓23科45属77种,腐木生苔藓14科25属32种。石生苔藓物种丰富度最高,其次为树附生苔藓,再次为土生苔藓,腐木生苔藓物种最少。经过观察,保护区内树附生苔藓大多生长位置不高且和腐木生苔藓一样多聚集或交织生长在一块,而土生苔藓和石生苔藓也多成斑块状聚集生长。
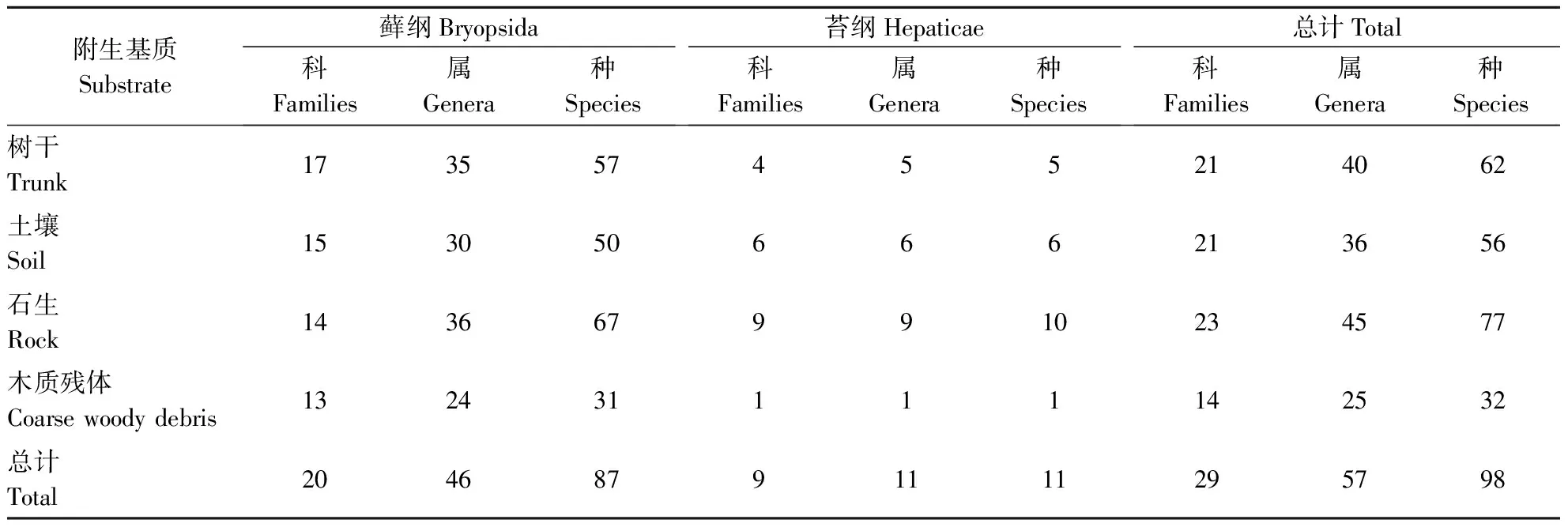
表2 不同基质上附生苔藓物种数Table 2 Epiphytic bryophytes species on different substrates
2.2不同生长基质苔藓的重要值研究
由于调查时部分木质残体上苔藓的面积缺失,暂不计算腐木生苔藓的重要值和多样性,仅对树附生苔藓、土生苔藓、石生苔藓的重要值和多样性进行计算(表3)。总的样点数(除木质残体外)共686个,其中羊角藓(Herpetineurontoccoae)、长肋青藓(Brachytheciumpopuleum)、圆叶匐灯藓(Plagiomniumvesicatum)、长叶纽藓(Tortellatortuosa)、长柄绢藓(Entodonmacropodus)、短叶毛锦藓(Pylaisiadelphayokohamae)、毛尖青藓(Brachytheciumpiligerum)、硬叶对齿藓(Didymodonrigidulus)、皱叶绢藓(Entodonplicatus)、大羽藓(Thuidiumcymbifolium)分别出现214、223、173、161、92、105、91、73、80、90次,其重要值在2.79~8.86,位于前10位,是1 hm2样地中苔藓的优势种。在不同生长基质中苔藓的重要值排序有所差异,在树附生苔藓中羊角藓占主导地位,土生苔藓中金发藓科的小仙鹤藓是优势种,石生苔藓中青藓科的长肋青藓是优势种。
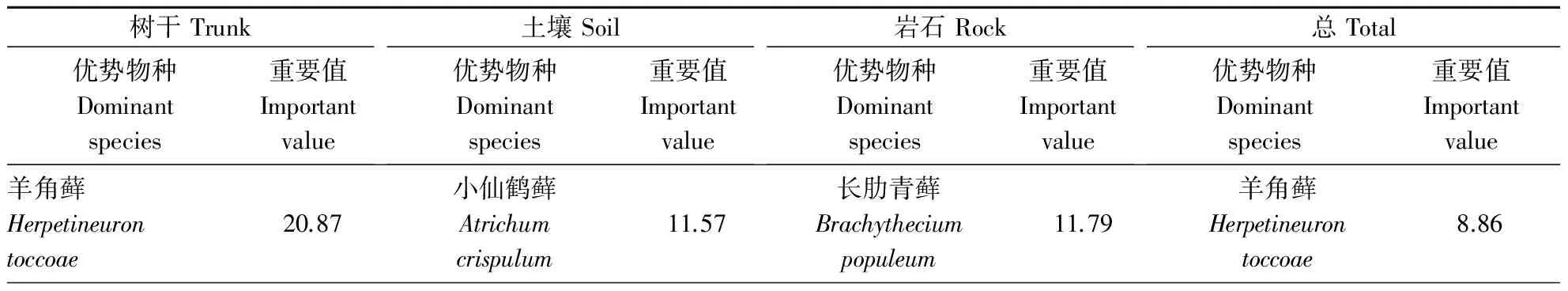
表3 不同附生基质上重要值前10位的苔藓Table 3 Top 10 dominant epiphytic bryophytes ranked by important value on different substrates

续表Continued table
2.3不同附生基质苔藓的多样性指数研究
通过重要值计算的物种多样性指数如表4。宝天曼1 hm2样地内所有苔藓的Shannon-Wiener多样性指数和Simpson优势度指数分别为3.868和0.96,石生苔藓的Shannon-Wiener多样性指数和Simpson多样性指数均大于土生苔藓,土生苔藓的多样性指数大于树附生苔藓,说明石生苔藓的多样性是最丰富的,土生苔藓次之,树附生苔藓的多样性最小。
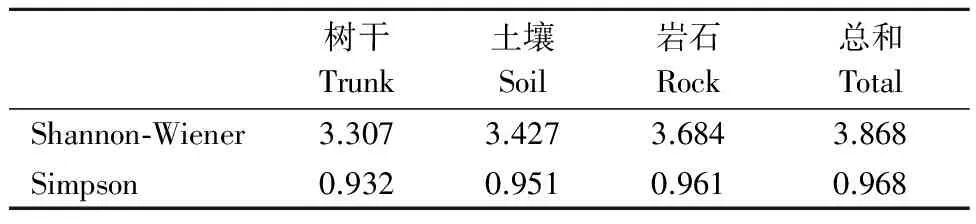
表4 不同生长基质上苔藓的多样性Table 4 Species diversity of bryophytes on different substrates
2.4不同生长基质上苔藓物种的相似性研究
本研究中一些苔藓对不同生长基质均有较强的适应性,土壤与岩石上附生苔藓的共有种数最多,达48种,林地与腐木表面苔藓的共有物种数最少,有23种。采用Sφrensen指数相似性系数计算苔藓在不同附生基质间的物种相似性系数(表5),通过比较发现,石生与土生苔藓物种相似性指数最高,土生和腐木生苔藓的物种相似性指数最低。不同生长基质苔藓相似性系数与各生长基质间共有种的数量变化趋势相同。
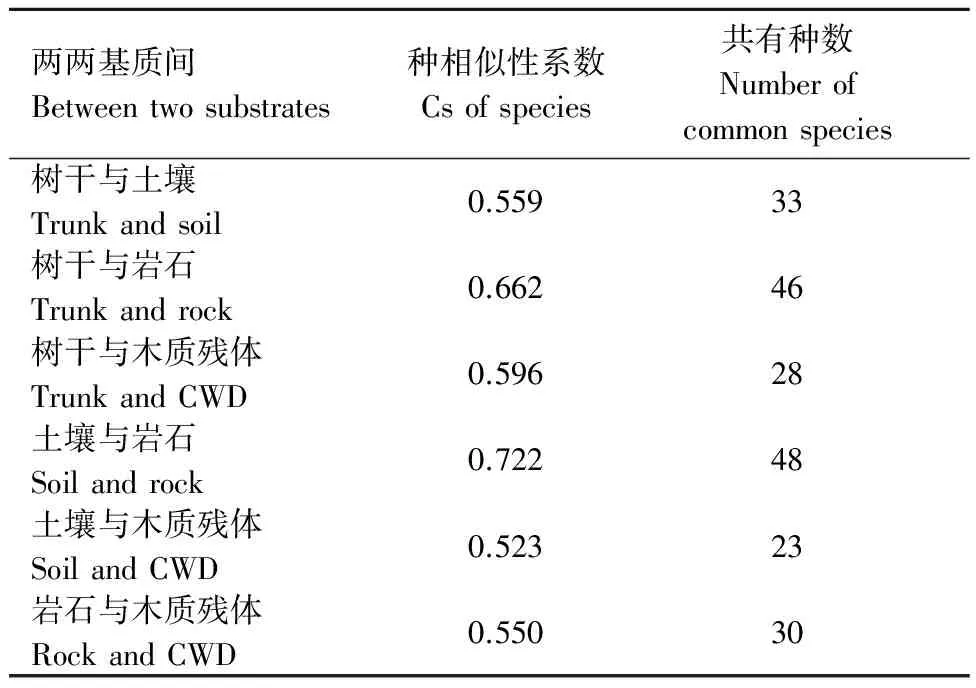
表5 附生基质间共有种与相似性系数Table 5 The number of common species and Sφrensen’s similarity coefficient between substrates
2.5不同生长基质苔藓生活型研究
宝天曼1 hm2样地内生长的苔藓涵盖了平铺型、交织型、丛集型、垫状型和扇形5种苔藓生活型。不同生长基质中各生活型的苔藓重要值的比值见表6。从表6可以看出,5种生活型在各附生长基质中均有出现,样地内苔藓以平铺型为主,垫状最少。树生苔藓中交织型和垫状型苔藓所占比例明显高于其他生长基质,土生苔藓除了以平铺型苔藓为主外,扇形和丛集型苔藓所占比例明显高于其他基质。
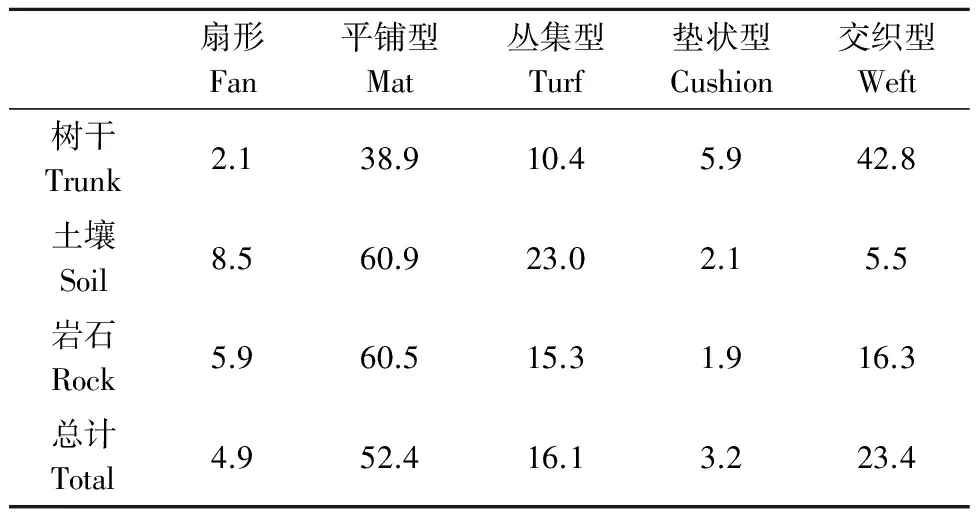
表6 不同生长基质中各生长型苔藓的比例Table 6 Ratio of bryophyte life-forms in different substrates %
3 结论与讨论
3.1不同基质上苔藓物种多样性分析
宝天曼样地内分布着丰富的苔藓植物,共有苔藓29科57属98种,藓纲植物20科46属87种,苔纲植物9科11属11种。不同生长基质上苔藓物种组成上存在差异,石生的物种数最多,树生和土生次之,腐木生物种最少。通过对各基质上苔藓重要值计算发现,树附生苔藓中牛舌藓科的羊角藓是优势种,同时青藓科(毛尖青藓和长肋青藓)较多;土生苔藓中金发藓科的小仙鹤藓是其优势种,较其他基质棉藓科(圆条棉藓和垂蒴棉藓)显著增多;石生苔藓中青藓科的长肋青藓是其优势种,较其他基质丛藓科(硬叶对齿藓和长叶纽藓)增多,同时树干上的中华耳叶苔是整个样地中仅有的重要值排进前10位的苔类。
不同生长基质上生长的苔藓的Shannon-Wiener多样性指数和Wimpson优势度指数比较结果显示,总体>石生>土生>树附生苔藓植物,多样性指数的差异可能与不同生长基质为苔藓提供的水分、温度、光照、矿质营养等条件以及环境稳定性和优势种种间相互关联有关[6],苔藓对环境变化敏感,林下小生境越丰富,苔藓多样性越高[16]。岩石附近树木较少,且枯枝落叶较少覆盖于岩石表面,表面光照条件较好,且得益于林内良好的水分环境,长时间的演替使得岩石上苔藓最为丰富[6];土壤能够为苔藓提供较好的营养条件,但易受到枯落物覆盖的影响,光照条件较岩石和树干较差,因此土生苔藓生物多样性次于石生苔藓;树附生苔藓的多样性指数最小,这可能是由于树附生苔藓植物中羊角藓重要值远高于其他优势种,羊角藓较强的竞争力,抑制了其他苔藓的生长所致[17]。
3.2不同附生基质苔藓物种相似性分析
样地内不同生长基质间苔藓组成相似性指数最小为0.523,最大为0.722,并且有19种苔藓在4种生长基质上均有分布,这可能与苔藓独特的生理生态适应机制有关。苔藓会随着环境变化适应不同的生长基质[18],随着湿度等环境条件的改善,苔藓会降低对生长基质的专一要求,向多种生长基质拓展生境[19]。宝天曼自然保护区年均降雨量885.6 mm,空气湿度大,水分条件不再是限制苔藓分布的重要因素,导致物种向多种基质拓展,因此各生长基质间苔藓植物相似性高。
在不同生长基质间,土生和石生苔藓间的相似性指数最高。这可能是由于岩石在风化和生物的作用下转变为土壤,石生的苔藓有一部分继续在新生的薄土层中生存,其提供的环境条件与林下土壤相似所致[20]。同时,树生与石生苔藓的相似性指数较高,与土生苔藓的相似性指数较低。这可能是由于树生苔藓和石生苔藓的养分条件较为相似,与土壤中的养分相比都较为贫瘠,都无法像土生苔藓一样获取较为丰富的营养元素所致。
与树生苔藓和其他生长基质上苔藓的相似性类似,腐生苔藓与石生苔藓的相似性大于与土生苔藓的相似性。这可能是由于腐生苔藓大多来自树干上原有的树生苔藓,因此腐生苔藓与树生苔藓的性质较为相似。而且腐生苔藓物种数最少,本身与其他几种基质的相似性均较低,因此本研究中土生与腐木生苔藓物种相似性最低。这与郭水良等[21]发现的腐木生更接近于地面生苔藓的结果不同,可能是因为宝天曼地处暖温带-亚热带过渡区,较长白山气候湿润,气温较高,样内木质残体腐烂较快,没有足够时间使地面生苔藓向木质残体拓展所致。
3.3不同生长基质苔藓生活型分析
生活型是植物对环境的一种适应,各基质上附生苔藓的生活型组成比例有一定的差异,这可能与不同基质提供的水分、光照条件有关[22]。附生苔藓对外界环境的要求在不同的生活型之间存在着差异:从垫状型、丛集型到平铺型,其耐旱能力逐渐减弱[22],而扇形则对大气湿度具有苛刻的要求[7]。总体来看,划分的5种生活型在各生长基质中均有出现,并以平铺型为主,这也表明样地内具有良好的水分条件。但不同生长基质上,各种生活型的比例有所差异。其中石生和土生苔藓也均以平铺型为主,但是树生苔藓以耐旱较强的交织型为主,并且耐旱性较差的平铺型也占较大比例。这可能是由于样地中树干本身距地面较高,水分条件比石生和土生环境的差;而且由于树干呈圆柱形,存在阴面和阳面,阳面光照强湿度低,阴面光照弱湿度相对高,同时存在的2种环境导致树干上既存在耐旱较强的交织型苔藓,又存在耐旱性较差的平铺型苔藓,并且交织型苔藓略微占优势。
本研究对宝天曼国家自然保护区苔藓研究能较好地反映亚热带—暖温带过渡区的苔藓多样性,样地内石生苔藓最为丰富,土生苔藓次之,树附生苔藓多样性最低,石生苔藓和土生苔藓的物种组成相似性指数最高,土生苔藓与腐木生苔藓相似性指数最低。苔藓的生活型能较好地反映苔藓对不同基质的适应性,同时平铺型占优势也说明了样地良好的水分状况。但不同生长基质上苔藓组成和影响其生长的环境因素间的详细关系还需进一步研究。
[1] 汪庆, 吴鹏程. 苔藓植物的多样性研究[J]. 生物多样性, 1999, 7(4): 332-339.
[2] 吴鹏程. 苔藓植物生物学[M]. 北京: 科学出版社, 1998.
[3] 刘文耀, 马文章, 杨礼攀. 林冠附生植物生态学研究进展[J]. 植物生态学报, 2006, 30(3): 522-533.
[4] HUMPHREY J W, DAVEY S, PEACE A J, et al. Lichens and bryophyte communities of planted and semi-natural forests in Britain: the influence of site type, stand structure and deadwood[J]. Biological Conservation, 2002, 107(2): 165-180.
[5] 曹同, 郭水良. 长白山主要生态系统苔藓植物的多样性研究[J]. 生物多样性2000,8(1): 50-59
[6] 马文章, 刘文耀, 宋亮. 哀牢山中山湿性常绿阔叶林不同生长基质苔藓植物的组成与分布特征[J]. 植物分类与资源学报,2011, 33(4) : 443 - 450.
[7] BATES J W. Is′ life-form′a useful concept in bryophyte ecology?[J]. Oikos, 1998, 82(2): 223-237.
[8] 马文章, 刘文耀, 杨礼攀, 等. 边缘效应对山地湿性常绿阔叶林附生植物的影响[J]. 生物多样性, 2008, 16(3): 245 - 254.
[9] MA W Z, LIU W Y, LI X J. Species composition and life forms of epiphytic bryophytes in old-growth and secondary forests in Mt. Ailao, SW China[J]. Cryptogamie Bryologie, 2009, 30(4): 477-500.
[10] 谢斐, 杨再超, 左经会, 等. 贵州山地森林公园地面生苔藓植物多样性[J]. 北方园艺, 2015(9): 71-75.
[11] 袁志良, 陈云, 韦博良, 等. 暖温带-北亚热带生态过渡区物种生境相关性分析[J]. 生态学报, 2013, 33(24): 7819-7826.
[12] 贾宏汝. 环境空间异质性对暖温带-北亚热带过渡区植物群落构建的影响[D]. 郑州: 郑州大学, 2015.
[13] CONDIT R. Research in large, long-term tropical forest plots[J]. Trends in Ecology & Evolution, 1995, 10(1): 18-22.
[14] 马克平, 钱迎倩. 生物群落多样性的测度方法[J]. 生物多样性, 1994, 2(3): 162-168.
[15] KREBS C J. Ecological methodology[M]. New York: Harper & Row Press, 1989.
[16] RAMBO T R, MUIR P S. Forest floor bryophytes ofPseudotsugamenziesii-Tsugaheterophyllastands in Oregon: influences of substrate and overstory[J]. Bryologist, 1998, 101(1): 116-130.
[17] HOFFMAN G R, KAZMIERSKI R G. An ecologic study of epiphytic bryophytes and lichens onPseudotsugamenziesiion the Olympic Peninsula, Washington: I. A description of the vegetation[J]. Bryologist, 1969,72(1): 1-19
[18] PIIPPO S. Epiphytic bryophytes as climatic indicators in Eastern Fennoscandia[M]. Helsinki:Finnish Botanical Pub. Board, 1982.
[19] TNG D Y P, DALTON P J, JORDAN G J. Does moisture affect the partitioning of bryophytes between terrestrial and epiphytic substrates within cool temperate rain forests?[J]. The Bryologist, 2009, 112(3): 506-519.
[20] 姜炎彬, 邵小明. 苔藓植物分布及其物种多样性的研究评述[J]. 植物科学学报, 2010, 28(3): 385-390.
[21] 郭水良, 曹同. 长白山森林生态系统腐木生苔藓植物生态分布的 DCA 排序研究[J]. 应用生态学报, 1999, 10(4): 399-403.
[22] BIRSE E M. Ecological studies on growth-form in bryophytes: II. Experimental studies on growth-form in mosses[J]. The Journal of Ecology, 1957, 45(3): 721-733.
(责任编辑:朱秀英)
BryophytediversityamongdifferentsubstratesinBaotianman
ZHANG Xu1, LI Peikun1, HU Jintao1, DU Xiaojun2, YE Yongzhong1, YUAN Zhiliang1
(1.College of Life Sciences,Henan Agricultural University, Zhengzhou 450002,China; 2.State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093,China)
The diversity of moss on different substrates were studied by important value, species diversity index and similarity index in Baotianman, Henan. The results showed this region boasted a rich bryophyte flora which yielded 98 species belonging to 29 families and 57 genera,Herpetineurontoccoaeas the dominant species, and there were 77 species growing on rock with the largest number; The diversity index decreased in turn by hosting rock, soil and trunk; The similarity index of soil and rock was the 0.722 largest of all indexes; The mat was a dominant life forms of bryophytes, which reflected good water conditions.Thus, there existed certain differences for bryophytes to adapt to different substrates, and habitat diversity was beneficial for the diversity of bryophytes.
bryophyte; life form; species diversity; substrates; Baotianman
Q948
:A
2016-11-14
河南省教育厅科学技术研究重点项目(14A180013)
张 旭(1993-),男,河南洛阳人,硕士研究生,主要从事植物生态学研究。
袁志良(1976-),男,河南信阳人,副教授,博士。
1000-2340(2017)03-0377-06
