新型的以eEF2激酶-调控自噬信号通路为靶点的抗肿瘤药物网络分析△
2017-09-22柳欢张珂王航宇钟益宁
柳欢,张珂,王航宇*,钟益宁*
(1.广西中医药大学,广西 南宁 530299;2.石河子大学 药学院,新疆 石河子 832002)
·基础研究·
新型的以eEF2激酶-调控自噬信号通路为靶点的抗肿瘤药物网络分析△
柳欢1,张珂2,王航宇2*,钟益宁1*
(1.广西中医药大学,广西 南宁 530299;2.石河子大学 药学院,新疆 石河子 832002)
目的:eEF-2激酶有可能是哺乳动物大自噬的重要组分,研究寻求在电脑中识别以eEF-2K调控的细胞自噬途径为靶点的新的抗肿瘤药物。方法:使用一系列的生物信息学方法和实验步骤,如网络构造(network construction)、枢纽蛋白识别(hub protein identification)、微阵列芯片分析(microarray analyses)、靶向microRNA预测(targeted microRNA prediction)和分子锚定(molecular docking),确定新抗肿瘤药物。结果:通过几个在线数据库的数据,并进一步将它们修饰进入基础的eEF-2K相关联蛋白质之间的反应网络,利用电脑构建了一个全球人类网络工作。以自噬基因的不同表达为基础,确定了eEF-2K调控的自噬中的枢纽蛋白。随后,确认了miRNA能够以eEF-2K调控的自噬为靶向。最后,在DrugBank和ZINC数据库中扫描了一系列能够在乳腺瘤细胞中以eEF-2K为靶点的分子合成物。结论:系统地确认了以eEF-2K为靶点的新的小分子产物,并预测了靶点抗肿瘤药物。这些发现可能为揭示eEF-2K和自噬之间的复杂关系、并为发现可能的肿瘤药物提供了进一步的线索。
eEF-2K;细胞自噬;肿瘤;AKT
细胞自噬是通过自噬泡和自噬溶酶体降解细胞质和细胞器,并将细胞组分回收用于能量利用的一个高度保守的过程。在饥饿或生长因子失效时,细胞自噬可能通过提供不同的能量资源作为一个暂时的存活机制[1-2]。当面对低氧或新陈代谢压力时,细胞自噬可以完善快速生长细胞的营养利用,也因此对肿瘤细胞的存活有贡献作用[3-5]。eEF-2K是钙调蛋白依赖型蛋白激酶,并通过磷酸化eEF-2负调节蛋白质合成。eEF-2是100kDa蛋白,其通过在核糖体的A和P位促进肽链和tRNA间的转换过程,从而调节肽链延长部分的转移。在Thr56位被eEF-2K催化磷酸化后的eEF-2将降低核糖体与蛋白质延长因子之间的吸引力[6]。我们前期的研究描述了eEF-2K可能通过响应营养缺失而作为哺乳动物大自噬途径的中心组分[7-8]。在压力促进的自噬中,eEF-2K担任的角色已经被其他研究确认[9]。因为蛋白质合成是主要的能量消耗过程,通过eEF-2K活动而导致的蛋白质合成停止和细胞自噬促进可大量保存能量,并在新陈代谢压力下支持细胞生存。此外,相较于周围的其他正常组织,eEF-2K已被证实在多种乳腺癌细胞族和人类乳腺癌组织中有过度表达和活性增加的情况。通过观察,eEF-2K在癌症中的作用可正调节多种类型的肿瘤组织,其中包括恶性神经胶质瘤或者乳腺瘤[10-13];抑制此激酶会导致多种肿瘤细胞的生存能力下降[14]。近期又发现了eEF-2K可能在包括营养缺乏、生长因子抑制、由糖分解抑制剂2-脱氧葡糖导致的能量压力等的环境或者新陈代谢的压力下,对细胞自噬起到正调节(positive regulator)作用[15]。
在此研究中,我们寻求在电脑中识别以eEF-2K调控的细胞自噬途径为靶点的新的抗肿瘤药物。我们的研究显示,eEF-2K调控的细胞自噬在肿瘤中起到了细胞保护作用,且抑制细胞自噬性生存,可调高对治疗药物的敏感性。
1 材料和方法
1.1 数据处理与网络构建
为构建全球人类蛋白质交互作用(PPI)网络,我们从6个不同的人类PPI数据库收集了多种蛋白质交互作用的资料,这6个不同的数据库有:Human Protein Reference Data(HPRD)[16],Bio molecular Object Network Databank(BOND)[17],IntAct[18],HomoMINT[19],BioG RID[20]和Database of Interacting Protein(DIP)[21]。接着,依据Gene Ontology(GO)在一对蛋白中至少有一个为自噬相关蛋白,我们构建了细胞自噬的PPI网络。然后,依据在肿瘤细胞自噬性死亡的前期报道,我们进一步提取出eEF-2K调控的细胞自噬的枢纽蛋白和相关的途径[22]。据此,以上所提及的枢纽蛋白可以与eEF-2K结构组成一个经修饰的eEF-2K自噬网络,在此网络中,每个蛋白在癌症环境下都可以与eEF2K相反应。
1.2 枢纽蛋白的多重分析
我们通过以下3个黄金准则决定自噬相关蛋白是不是自噬枢纽蛋白:第一,在功能联系网络中,每个蛋白等级以它与其他蛋白相联系的数目计算[23]。基于高等级的蛋白倾向于在蛋白网络中担任着更重要作用的假设,因此,高等级蛋白会被认为是枢纽蛋白。第二,在凝集素促进的肿瘤细胞中,枢纽蛋白应该与已知的自噬相关蛋白有联系作用,这样在开发自噬相关的新靶点时这些枢纽蛋白就更值得关注。而这个方法与之前关于需要新的与肿瘤相关的基因研究相似[24]。第三,一个以自噬为模型的网络组件对确定枢纽蛋白也是必要的,因为这些枢纽蛋白在“密集区”会有所增加[25]。在此研究中,我们致力于在植物凝集素促进的肿瘤细胞中明确自噬枢纽蛋白,因此,结合已知的枢纽蛋白和通过之前提及的3个黄金准则,可以确定自噬枢纽蛋白。
1.3 基于微阵列芯片的动态eEF-2K调控途径的分析
可以与同一个蛋白相互反应的蛋白往往具有相似的基因表达模式,所以基因可以一起表达的蛋白比不能一起表达的更可能发生反应。为了确定某种特定的基因是否可以共同表达,我们使用了微阵列数据(NO.E-GEOD-26459)在三苯氧胺(tamoxifen)处理的MCF-7细胞中,为确定在自噬中成对合作表达的基因,用2.5 mmol·L-1DTT处理[26]。在控制的实验组中,微阵列重复使用了3次。E-GEO数据库提供了正确的和标准的前期数据。因此,数据前期处理需达到微阵列的必要分析(SAM)的标准,且相应的探针来自于Uniport数据库[27]。鉴于原始实验数据未进行log2过程,我们在数据模型汇总加入了log2指令和T-test,利用在SAM中的“Two Class”(未成对)模型来分析不同的基因表达。通过以上这些测试,我们运用SAM的Δ变量来确定0.13并列出所有表达不同(正调节或者负调节)的基因。首先我们使所有的数据进行log2变换标准化,再使用平均连锁来聚连这些数据(在此项操作之前,我们已提取这些基因的表达数据)。当这些表达数据的数值是2时,我们采用其平均值,如果是3或者更大的数值,我们取其中位数。最后,我们把微阵列数据绘制到SAM图表中,并评估它。我们同时也使用Cluster和TreeView[28]来搜集所有不同的基因表达,为下一步的分析做准备。此外,我们使用一个相似的方法来确定协同表达的miRNA。我们选择的微阵列数据(NO.E-GEOD-26459)是来自于在三苯氧胺(tamoxifen)处理的MCF-7细胞[29]。
1.4 miRNA靶点预测
因为可用的预测模型有不同的敏感性和特异性,这些方法的综合应当可以大大降低正和负的误差,且只有通过3种不同的算法才能够认可所预测出的相互之间的反应。我们利用从Target(适度的配对、编号、编号类型、编号内容、除编号内容外最优先保存的选择排名)、MiRanda(适度的配对、编号、最配对的miRNA)[30]和Diana-MicroT(杂交的能量阈值规则)[31]中的人类特异性数据把miRNA和mRNA之间的关系总结起来。
1.5 序列对比
在序列对比当中,搜集了NCBI数据库中所有被报道过的不同种类生物中的eEF2K序列[32]。这些序列都通过ClustalW方法(version2.1进行了分析,蛋白质分析所用的是Clustal W方法)进行分析。
1.6 模型和分子锚定
eEF-2K最初的X射线晶体构型协调的三维几何学是来自于Protein Data Bank(http://www.pdb.org/)。我们下载了最新版本的DrugBank(http://www.drugbank.ca/)中的FDA批准的小分子化合物,并从最新的ZINC数据库(http://zinc.docking.org/)中的可商用的化合物来构建eEF-2K的扫描资料。之后,所有的候选化合物都通过Open Babel工具箱和ZINC数据库来进行所有的结构编辑。
此外,我们使用UCSF DOCK6.3软件[33]和AMBER力场参数来进行分子锚定计算。首先,基于网格的分数算法来排列所有的小分子药物,在分数算法中所有的药物足够灵活地对接受者(eEF-2K)进行回应。然后,运用DOCK6.3中的AMBER分数功能来重新排列之前基于网格运算分数前100的药物。在amber分数计算期间,PDB2PQR服务器[34]是利用自动化PDB文件准备和eEF2K的质子化作用状态作业。在我们的锚定过程中,方向的最大数量设置为500。
1.7 分子动力学(MD)模拟
MD模拟使用了GROMACS(4.0.5版本)软件包[35]来监测eEF2K和选中化合物的结合状态。两个选中的小分子化合物分别是FDA批准药物和ZINC数据库的分值最高的,用PRODRG2服务器[36]来生成配体拓扑。eEF2K的拓扑结构由使用GROMOS96 43al力场[37]的pdb2gmx服务器编辑而成。在这个MD过程中,5000 PS(1PS=735.498 75 W)的刺激以2 fs的时间步骤在执行,然后用VMD软件来观察产生的轨迹文件。
2 结果和讨论
2.1 人类PPI自噬网络的构成
在本研究中,我们基于IntAct、HPRD、HomoMINT、BOND、BioGRID和DIP计算构造了人类的总体PPI网络(大约85 806个蛋白偶对),包含了几乎全部PPI(见图1A)。为了构造一套真阳性基因对,实体PPI来自包括BioGRID的37 710个蛋白偶对(8982个蛋白质),BOND的8044个蛋白偶对(4073个蛋白质),HomoMINT的14 892个蛋白偶对(6240个蛋白质),HPRD的39 044个蛋白偶对(9614个蛋白质),IntAct的34 935个蛋白偶对(8849个蛋白质)以及DIP的12 809个蛋白偶对(4818个蛋白质)。这些结果表明,作为自噬PPI网络,总数为2633的独特的蛋白偶对已准备好(见图1B)。
2.2 癌症eEF2K-调节的自噬子网的识别
众所周知,PPI网络通常有复杂的性质,在众多信号通路有多个连接;因此,有必要在一个特定的生物背景下,通过进一步整合和分析高通量数据来代表这个网络,比如癌症。根据Table S1的数据,我们提取了所有PPI,包括致癌基因(MARK2、MK03、PAK1、SIK1、ULK1、ULK2、WNK2、PI3K)、肿瘤抑制(AAPK1、CDK1、EGFR、ERBB2、FAK2、JAK2、JAK3)和包括自噬PPI网络的mTOR在内的其他类型。随后,我们发现mTOR eEF2K AAPK1可能与其他参与自噬途径的蛋白质相互作用。Akt已经是众所周知涉及eEF2K-调节的自噬细胞死亡,我们手动添加Akt及其相关自噬通路到这个自噬通路中。因此,结合3个黄金标准和表1的数据,表明自噬通路是由15个自噬相关蛋白和Akt(总数为21)的相互作用形成的(见图1C)。例如,eEF2K的程度是37,其链接到从GO注释的自噬蛋白质的程度是7且位于密集区域(网络模块区域)。此外,Akt的程度是8,其链接是2且位于密集区域(模块)。同时,mTOR的程度是6;其链接是3且位于稀疏区。
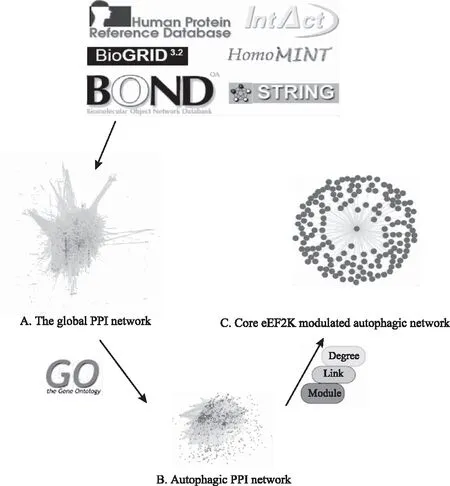
注:A.总体PPI网络;B.自噬PPI网络;C.eEF2K子网络。图1 乳腺癌eEF2K-调控的自噬通路的识别(204 mm×237 mm)
2.3 MCF-7细胞的eEF2K-调控通路的靶点miRNA的预测
在上述的15个自噬相关蛋白质中,我们分别通过TargetScan来预测了654个靶点miRNA,通过MiRanda来预测了2929个靶点miRNA,以及通过Diana-MicroH来预测了391个靶点miRNA。随后,我们综合了这些预测的相同结果,得到52个miRNA,其中hsa-miR-96、hsa-miR-100、hsa-miR-144、hsa-miR-17、hsa-miR-20、hsa-miR-20b、hsa-miR-106b、hsa-miR-93、hsa-miR-302a、hsa-miR-302b、hsa-miR-302c、hsa-miR-302d、hsa-miR-135a、hsa-miR-135b和hsa-miR-216a显示为枢纽靶点蛋白质(FAK2、JAK2、JAK3、MARK2、PAK1、ULK1、ULK2、WNK2、PI3K和MTOR)(见图2B)。在三苯氧胺处理过的自噬应激下的人类乳腺癌MCF-7细胞中,我们用微阵列显著性(SAM)分析处理了不同微阵列表达的数据,来识别正常和恶性细胞之间不同的基因表达(见图2B)。我们指明,这些蛋白质被确认为不同表达的蛋白质,提取功能枢纽蛋白质,依赖于基因共表达谱,从而在肿瘤的自噬应激下起关键的调控作用。由于这些微阵列数据,我们鉴别出了一些潜在的eEF2K-调控途径,比如乳腺癌中的AKT-eEF2K-mTOR途径。
值得注意的是,miRNA是高度保守的非编码内源性RNA,长度为22个核苷酸(nt),众所周知能调节癌症的凋亡和自噬的通路,表明了miRNA的致癌和肿瘤抑制功能[38-40]。此外,我们确认了新的致癌和肿瘤抑制的miRNA-调节和eEF2K-调节的自噬途径涉及了MCF-7细胞(见图2C)。结论是,一个自噬的AKT-eEF2K-mTOR途径可以被预测受hsa-miR-96、hsa-miR-100、hsa-miR-144(mTOR)、hsa-miR-561、hsa-miR-588、hsa-miR-661(Akt靶点)和hsa-miR-459、hsa-miR-22(eEF2K靶点)调控。我们先前的研究已报道了基于Bayesian模型的凋亡网络可以鉴别一些可能作为枢纽蛋白质的靶点的miRNA,这些蛋白有TP53、SRC、细胞周期-依赖激酶2/6、TNFR16/19和TGF-b受体1/2(39)。另外,我们最近的研究证明,在人类乳腺癌 MCF-7细胞中,自噬PPI网络可以确认9个自噬枢纽蛋白质和13个相关的致癌或肿瘤抑制miRNA。与以前的癌细胞凋亡或自噬的研究相比,我们首次报道了一些预测的乳腺癌细胞内eEF2K-调节的自噬途径的靶点miRNA。
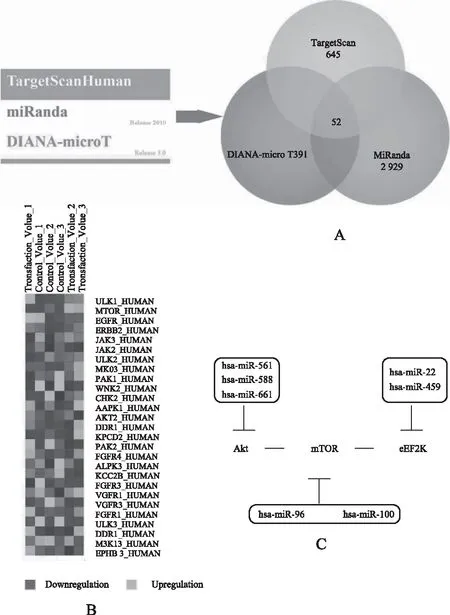
注:A.凋亡和自噬中靶点eEF2K相互作用的miRNA预测;B.乳腺癌中eEF2K-调控途径的基于miRNA的动力学;C.乳腺癌中eEF2K-调控途径的基于miRNA的动力学。图2 乳腺癌eEF2K-调控的自噬途径的识别(187 mm×256mm)
2.4 eEF2K激酶域的保守分析
为了跟踪eEF2K激酶域的保守性,笔者进行了人类和模式生物的eEF2K的主要氨基酸序列的比对。人类eEF2K激酶域与其模式生物的对应体有大量一致序列,表明了物种之间的保守性(见图3A)。基于这个证据,精制虚拟锚定工具的帮助下,eEF2K已经被选作一个潜在的抗癌药物发现的靶点。
2.5 通过分子锚定预测新eEF2K靶点药物
为了进一步锚定研究,对1447个FDA批准的小分子药物通过分子锚定模拟进行了虚拟筛选,首先基于方格网的分值排名前100的药物通过进一步琥珀锚定评估。两个阶段锚定过滤之后,我们分别确定了分值排名前20的药物作为初始匹配记录。预测结合到eEE2K新发现的10个抑制剂、相应的方格网分数和琥珀分数如表1所示,分数越低的结合相互作用越强。据我们所知,这20个药物至今还没有被报道过是eEE2K的靶点。
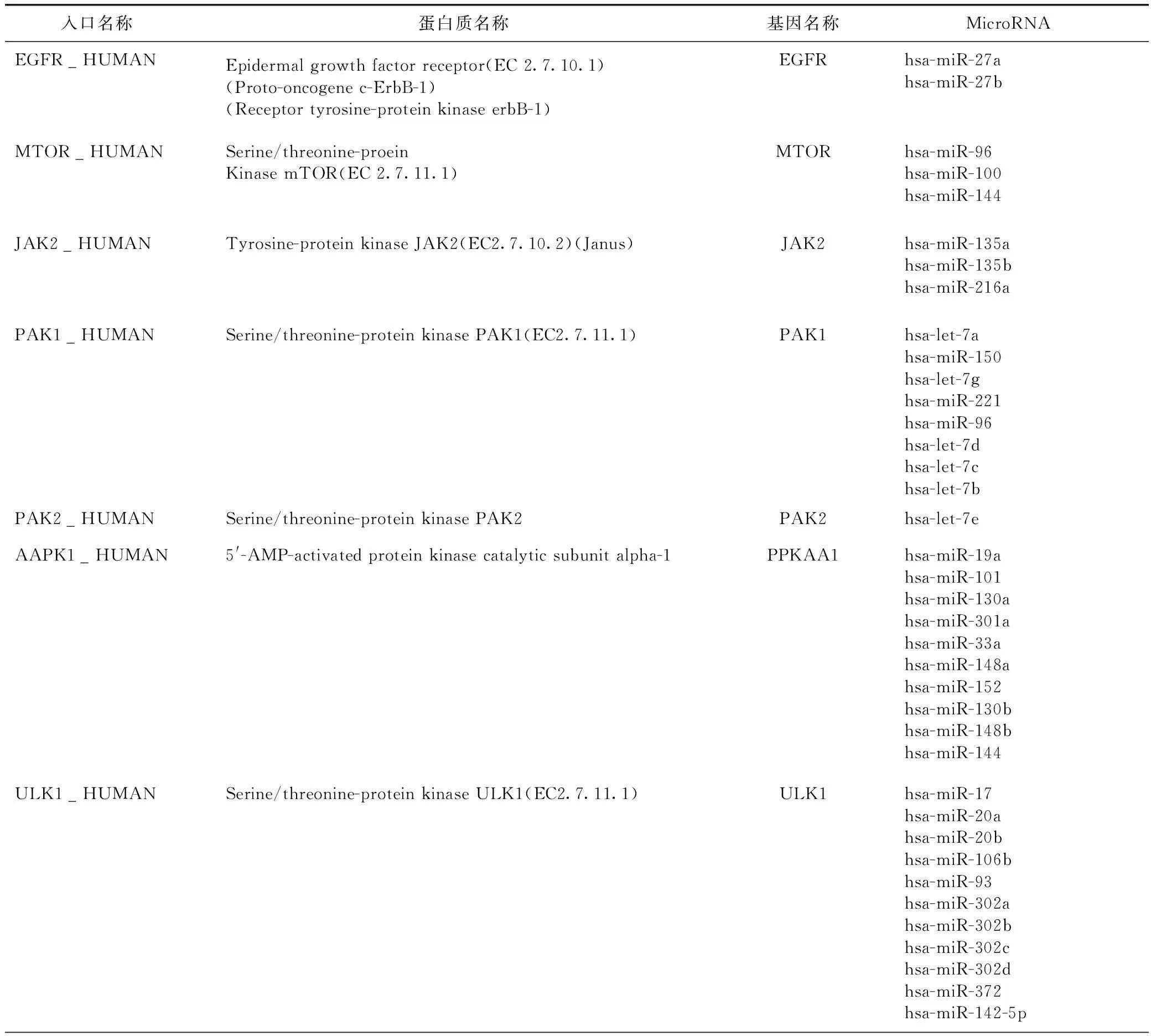
表1 抑制剂及相应的蛋白和基因

表1(续)
然后,对全球eEE2K结合模式和新发现的9个抑制剂进行了分析。如图3B所示,这20种药物中每个都以一个共同的模式结合到eEE2K,也就是药物结合到激酶两个域之间的铰链点上,是在ATP-结合位点。我们选定受体的三维结构中的抑制剂吡唑基吡咯的锚定结构的结合构型,与图3B所示相比,吡唑基吡咯似乎也稳定存在于eEE2K ATP-结合位点。考虑到一些抑制剂已被报道占据了激酶的变构位点,因此对于变构抑制剂功能,我们还研究了已鉴别抑制剂变构抑制eEE2K的可能性。对eEE2K整个结构(信号肽除外)锚定筛选后,没有观察到药物残留结合到除ATP-结合位点外其他部位的构象。由于激酶的ATP-结合位点有至关重要的作用,ATP-位点-指向激酶抑制剂的发展是永恒的,有希望从化合物数据库寻找新型抑制剂和从药品数据库执行创新药物开发,从而有选择性地以eEE2K为治疗干预靶点。随后,我们进一步分析了这两个药物的相关信息,如活性机制、药效、药物相互作用、靶点等。

注:A.eEF2K蛋白激酶域保守性分析;B.eEF2K建模和锚定以及小分子化合物。图3 蛋白激酶域保守性分析、分子建模和eEF2K锚定筛选小分子抑制剂(200 mm×241 mm)
2.6 分子动力学刺激的质量保证
对这2种预测药物(C03189329 C03830401)的MD刺激和相应的eEE2K进一步确认其精确结合机制和相互作用稳定性。均方根偏差(RMSD)是一个MD刺激时评估系统的稳定性的重要参数。如图4所示,分别在eEE2K和C03189329的第一次的1500 PS和1100 PS的快速增长后,有超过5000 PS力度,eEE2K-C03189329复合体达到稳定状态,eEE2K的平均主链RMSD和标准偏差在最后3000 PS为(0.20±0.025)Å;分别在eEE2K和C03830401的第一次的2500 PS和3000 PS的迅速增加之后,在eEE2K-C03830401复合体达到稳定状态,eEE2K的平均主链RMSD和标准偏差在最后2000 PS为(0.19±0.015)Å。上述数据显示,所有这些eEE2K-药物复合体经过一段时间的动态变化后达到了他们的稳定状态。2种eEE2K-药物系统的动能的动力学和潜在能量如图4所示,我们可以看到,这两个系统的动能和潜在的能量在整个MD过程中保持稳定。
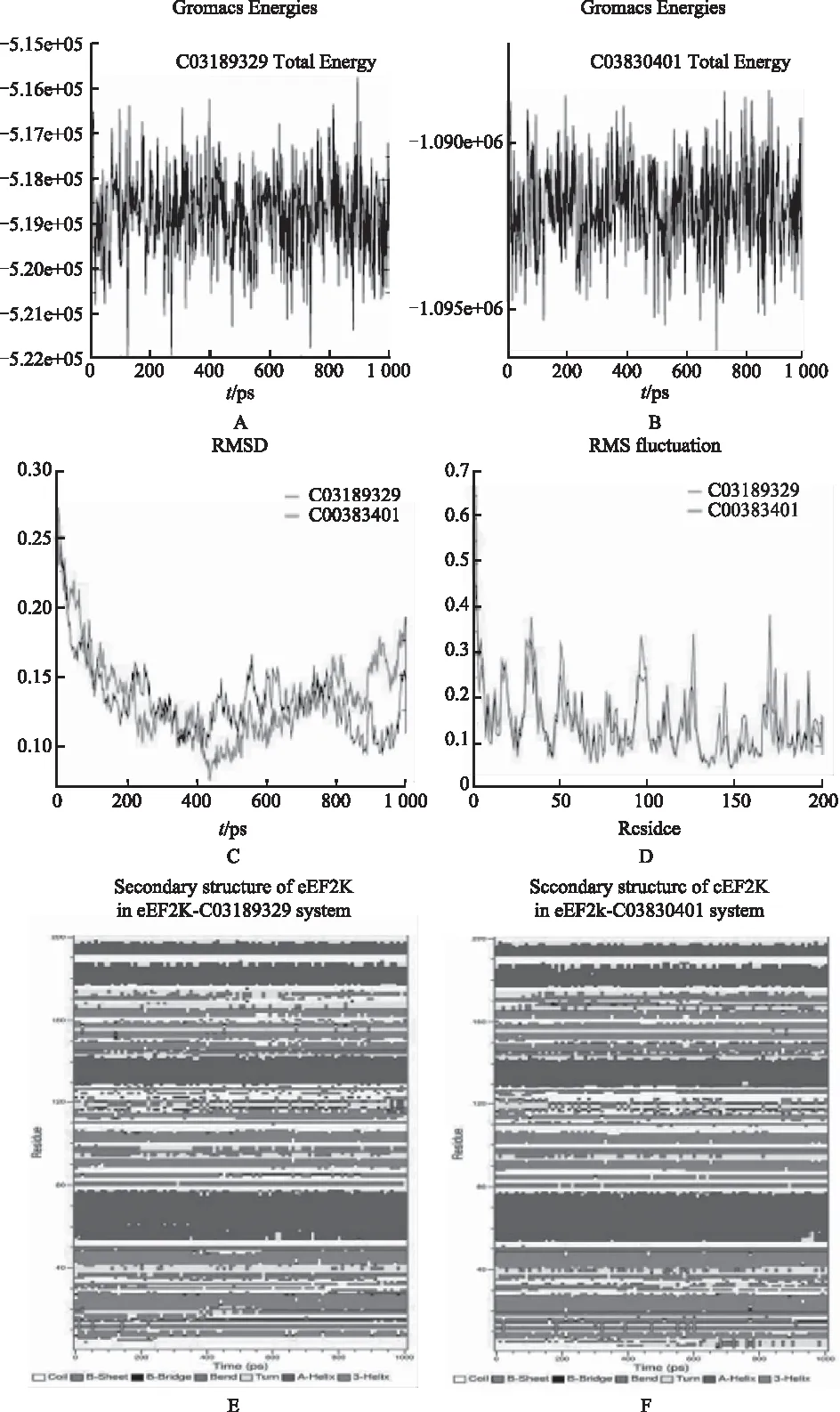
注:A.C03189329总能量;B.C03830401总能量;C.C03189329均方根偏差;D.C03830401均方根;E.eEF2K-C03189329系统中eEF2K的二级结构;F.eEF2K-C03830401系统中eEF2K的二级结构。图4 eEF2K结合的“最佳匹配”小分子的分子动态模拟(180 mm×293 mm)
2.7 eEE2K作为抗癌药物的潜在目标
基于MD刺激的结果,2种小分子eEE2K抑制剂(C03189329 C03830401)可以直接作为ERK2的ATP-结合位点的靶点并与ERK2稳定地进行相互作用。考虑到eEE2K参与不同的细胞过程,如细胞凋亡、分化和自噬,可以推测eEE2K靶向抑制剂通过调节自噬有希望成为一个新的临床抗癌药物。
[1] Guido K,Marja J.Lysosomes and autophagy in cell death control[J].Nat Rev Cancer,2005,5(11):886-897.
[2] Beth L,Junying Y.Autophagy in cell death:an innocent convict?[J].J Clin Invest,2005,115(10):2679-2688.
[3] Shengkan J,Dipaola R S,Robin M,et al.Metabolic catastrophe as a means to cancer cell death.[J].J Cell Sci,2007,120(Pt 3):379-383.
[4] Jin S,White E.Role of autophagy in cancer:management of metabolic stress.[J].Autophagy,2007,3(1):28-31.
[5] Kurt D,Robin M,Brian B,et al.Autophagy promotes tumor cell survival and restricts necrosis,inflammation,and tumorigenesis.[J].Cancer Cell,2006,10(1):51-64.
[6] Ryazanov A G,Shestakova E A,Natapov P G.Phosphorylation of elongation factor 2 by EF-2 kinase affects rate of translation.[J].Nature,1988,334(6178):170-173.
[7] Wu H,Yang J M,Jin S K,et al.Elongation factor-2 kinase regulates autophagy in human glioblastoma cells.[J].Cancer Res,2006,66(6):3015-3023.
[8] Wu H,Zhu H D,Niu T,et al.Silencing of elongation factor-2 kinase potentiates the effect of 2-deoxy-D-glucose against human glioma cells through blunting of autophagy.[J].Cancer Res,2009,69(6):2453-60.
[9] Py B F,Boyce M,Yuan J.A critical role of eEF-2K in mediating autophagy in response to multiple cellular stresses[J].Autophagy,2009,5(3):393-396.
[10] Parmer T G,Ward M D,Yurkow E J,et al.Activity and regulation by growth factors of calmodulin-dependent protein kinase III(elongation factor 2-kinase)in human breast cancer.[J].Brit J Cancer,1999,79(1):59-64.
[11] Bagaglio D M,Cheng E H,Gorelick F S,et al.Phosphorylation of elongation factor 2 in normal and malignant glial cells[J].Cancer Res,1993,53:2260-2264.
[12] Agaglio D M,Hait W N.Role of calmodulin-dependent phosphorylation of elongation factor 2 in the proliferation of rat glial cells.[J].Cell Growth Differ,1995,5(12):1403-1408.
[13] Cheng E H,Gorelick F S,Czernik A J,et al.Calmodulin-dependent protein kinases in rat glioblastoma.[J].Cell Growth Differ,1995,6(5):615-621.
[14] Arora S,Yang J M,Kinzy T G,et al.Identification and characterization of an inhibitor of eukaryotic elongation factor 2 kinase against human cancer cell lines[J].Cancer Res,2003,63(20):6894-6899.
[15] Cheng Y,Li H J,Ren X C,et al.Cytoprotective effect of the elongation factor-2 kinase-mediated autophagy in breast cancer cells subjected to growth factor inhibition[J].Plos One,2010,5(3):606-612.
[16] Prasad T S K,Kandasamy K,Pandey A.Human protein reference database and human proteinpedia as discovery tools for systems biology.[J].Methods in Molecular Biology,2009,577:67-79.
[17] Alfarano C,Andrade C E,Anthony K,et al.The Biomolecular interaction network database and related tools 2005 update[J].Nuceic Acids Res,2005,33(1):418-424.
[18] Kerrien S,Alam-Faruque Y,Aranda B,et al.IntAct-open source resource for molecular interaction data[J].Nucleic Acids Res,2007,35:561-565.
[19] Persico M,Ceol A,Gavrila C,et al.HomoMINT:an inferred human network based on orthology mapping of protein interactions discovered in model organisms[J].Bmc Bioinformatics,2005,6(Suppl 4):S21.
[20] Chris S,Bobby-Joe B,Andrew C A,et al.The BioGRID interaction database:2011 update.[J].Nuceic Acids Res,2011,39(1):637-640.
[21] Ioannis X,Lukasz S,Xiaoqun Joyce D,et al.DIP,the database of interacting proteins:a research tool for studying cellular networks of protein interactions[J].Nucleic Acids Res,2002,30(1):303-305.
[22] Fu L L,Zhou C C,Yao S,et al.Plant lectins:targeting programmed cell death pathways as antitumor agents.[J].Int J Biochem Cell Biol,2011,43(10):1442-1449.
[23] Agarwal S,Deane C M,Porter M A,et al.Revisiting date and party hubs:novel approaches to role assignment in protein interaction networks.[J].PLoS Comput Biol,2010,6(6):889-903.
[24] Ostlund G,Lindskog M,Sonnhammer E L.Network-based identification of novel cancer genes.[J].Mol Cell Proteomics,2010,9(4):648-655.
[25] Qi Y,Ge H.Modularity and dynamics of cellular networks[J].PLoS Comput Biol,2006,2(12):1502-1510.
[26] Laura G M,Jaehong P,Lihua Z,et al.High-throughput ectopic expression screen for tamoxifen resistance identifies an atypical kinase that blocks autophagy.[J].Proc Natl Acad Sci U S A,2011,108(5):2058-2063.
[27] Zhang S.A comprehensive evaluation of SAM,the SAM R-package and a simple modification to improve its performance[J].Bmc Bioinformatics,2007,8(14):1-12.
[28] Page R D M.Visualizing phylogenetic trees using TreeView//[M].Stenson P D,Ball E V,Mort M,et al.Current protocols in bioinformatics,2002:Chapter 6.
[29]Manavalan T T,Yun T,Appana S N,et al.Differential expression of microRNA expression in tamoxifen-sensitive MCF-7 versus tamoxifen-resistant LY2 human breast cancer cells[J].Cancer Lett,2011,313(1):26-43.
[30] Friedman R,Farh K C,Bartel D.Most mammalian mRNAs are conserved targets of microRNAs[J].Genome Res,2011,19(1):92-105.
[31] Krek A,Grün D,Poy M N,et al.Combinatorial microRNA target predictions[J].Nat Genet,2005,37(5):495-500.
[32] Higgins D G,Sharp P M.CLUSTAL:a package for performing multiple sequence alignment on a microcomputer.[J].Gene,1988,73(1):237-244.
[33] P Therese L,Brozell S R,Sudipto M,et al.DOCK 6:combining techniques to model RNA-small molecule complexes[J].RNA,2009,15(6):1219-1230.
[34] Dolinsky T J,Paul C,Hui L,et al.PDBPQR:expanding and upgrading automated preparation of biomolecular structures for molecular simulations[J].Nucleic Acids Res,2008,35(14):522-525.
[35] David V D S,Lindahl E,Hess B,et al.GROMACS:fast,flexible,and free[J].J Comput Chem,2005,26(16):1701-1718.
[36] Schüttelkopf A W,Van Aalten D M F.PRODRG:a tool for high-throughput crystallography of protein-ligand complexes[J].Acta Crystallogr D Biol Crystallogr,2004,60(8):1355-1363.
[37] Kunz A P E,Allison J R,Geerke D P,et al.New functionalities in the GROMOS biomolecular simulation software[J].J Comput Chem,2012,33(3):340-353.
[38] Humphrey W,Dalke A,Schulten K.VMD:visual molecular dynamics.[J].J Mol Graph Model,1996,14(1):33-38.
[39] Fu L L,Xin W,Bao J K,et al.MicroRNA-modulated autophagic signaling networks in cancer[J].Int J Biochem Cell Biol,2012,44(5):733-736.
[40] Fu L L,Zhao X,Xu H L,et al.Identification of microRNA-regulated autophagic pathways in plant lectin-induced cancer cell death[J].Cell Prolif,2012,45(5):477-485.
NetworkAnalysisofNovelAntitumorDrugTargetingeEF2KinaseRegulatingAutophagySignalingPathway
LIUHuan1,ZHANGKe2,WANGHangyu2*,ZHONGYining1*
(1.GuangxiUniversityofTraditionalChineseMedicine,Nanning530299,China;2.SchoolofPharmacy,ShiheziUniversity,Shihezi832002,China)
Objective:EEF-2 kinase is likely to be important component of large mammals autophagy, seek recognition in computer with eEF-2 k regulation of cell autophagy pathway for targets of new antitumor drugs.EEF-2 kinase is likely to be an important component of mammalian autophagy,and a new anti-tumor drug targeting the eEF-2 kinase signaling pathway may solve this problem.Methods:A series of biological information method and several experiment steps such as network construction,hub protein identification,microarray analyses,targeted microRNA prediction and molecular docking were applied for the determination of new antitumor drugs.Results:The data form several online database were modified into the basis of the eEF-2K associated protein interaction network and a global network of human work was built by authors.Based on the different expression of autophagy genes,the key protein of eEF-2K regulating autophagy was determined.Subsequently,it was confirmed that miRNA was able to target the autophagy of eEF-2K.Finally,in the DrugBank and ZINC databases,a series of molecular compounds that can targeted at eEF-2K in breast tumor cells were scanned.Conclusion:the new small molecule product with eEF-2K as the target was confirmed,and the antitumor drugs were predicted.These findings may provide further clues to reveal a complex relationship between eEF-2K and autophagy and discovery the possible cancer drugs.
eEF-2K;autophagy;tumor;AKT
2016-01-12)
广西科技攻关计划项目(桂科重1298001-2-7,桂科攻1346008-4);广西自然基金(2014GXNSFAA118271);2014年博士研究生教育发展项目(050140002)
*
钟益宁,副教授,研究方向:中药药效物质基础研究,E-mail:1186638898@qq.com;王航宇,副教授, 研究方向:中药和天然药物研究,E-mail:18909932852@189.cn
10.13313/j.issn.1673-4890.2017.3.009
