基于毛木耳CDS的福寿螺多功能纤维素酶基因密码子优化分析
2017-09-15贾定洪李小林彭卫红
贾定洪,王 波,李小林,谭 伟,彭卫红*
基于毛木耳CDS的福寿螺多功能纤维素酶基因密码子优化分析
贾定洪1,2,王 波1,李小林1,谭 伟1,彭卫红1*
(1.四川省农业科学院土壤肥料研究所,四川成都 610066;2.四川大学生命科学学院生物资源与生态环境教育部重点实验室,四川成都 610064)
【目的】克隆福寿螺多功能纤维素酶基因(multi-functional cellulase gene,M fc),进行密码子优化改造,便于后续毛木耳Mfc基因转导研究。【方法】采用CodonW软件分析福寿螺多功能纤维素酶基因Mfc及毛木耳转录本CDS密码子使用次数和同义密码子相对使用度差异,以毛木耳CDS密码子特征对福寿螺M fc基因进行密码子优化。【结果】改造后的Mfcsg序列GC含量从原始Mfc序列的54.04%增加到58.16%,与毛木耳遗传背景一致。【结论】实验优化后获得的Mfcsg基因序列GC含量与毛木耳遗传背景一致,没有发夹结构、重复序列及反向重复序列,能够作为毛木耳外源基因转化应用。
福寿螺;毛木耳;密码子优化
【研究意义】能源资源是国家的重要财富,也是一种对国家而言必备的生产要素,在社会经济的发展中起到极为重要的作用,具有极为深远的战略地位[1]。目前能源供应中化石能源在世界能源消费结构中所占比例长期保持在85%以上[2],是世界经济的发展重要驱动力量。但由于化石能源存在不可再生性及污染环境的缺点,因而替代能源的发掘就变得十分迫切[3]。相对于化石能源,生物能源是一种可再生及环境友好型替代资源[4]。纤维素降解酶类能够将难以利用的纤维基质分解为可再生的生物燃料,是一种价值巨大的生物资源[3]。【前人研究进展】纤维素酶(cellulase)是将纤维素降解成纤维二糖和葡萄糖等小分子物质的一类酶的总称。纤维素降解生成葡萄糖需要至少3种纤维素酶协同作用才能完成,分别是内切葡聚糖酶(endo-1,4-β-D-glucanase)、外切葡聚糖酶(exo-1,4-β-D-glucanase)和β-葡萄糖苷酶(β-D-glucosidase)[5]。而据报道从福寿螺(Ampullaria crossean)胃组织分离的多功能纤维素酶基因(multi-functional cellulase gene,MFC)能够编码内切葡聚糖酶(endo-1,4-β-D-glucanase)、外切葡聚糖酶(exo-1,4-β-D-glucanase)和β-葡萄糖苷酶(β-D-glucosidase)这3种纤维素酶[5],应用前景看好。目前已在灰盖鬼伞[6]和草菇等食用菌进行了转化研究。但多功能纤维素酶基因(MFC)来源于福寿螺组织,在其它物种细胞中不一定能够稳定高效表达。因为每个氨基酸在同义密码子使用上都存在明显的物种偏好性[7],并且密码子优化与否直接决定着mRNA的稳定性,进而影响核糖体易位和蛋白翻译[8]。因此,异源基因的密码子优化改造对于外源基因的遗传转化表达十分必要。【本研究切入点】为了使福寿螺组织克隆得到的多功能纤维素酶基因(MFC)能够在毛木耳中进行有效稳定表达,课题拟开展福寿螺MFC基因与毛木耳转录本CDS密码子偏好性比较分析,并依据毛木耳密码子偏好性特征优化MFC序列。【拟解决的关键问题】为福寿螺多功能纤维素酶基因(MFC)在毛木耳细胞转化表达提供支撑,并为毛木耳等相关食用菌种类的功能基因发掘及改造等研究提供参考。
1 材料与方法
1.1 供试材料
供试材料:福寿螺,购自成都市青石桥市场。毛木耳菌株“黄耳10号”保存于四川省农业科学院土壤肥料研究所。感受态细胞DH5α购自上海迈其生物科技有限公司。
试剂:本试验RNA提取、1stcDNA合成试剂盒购自南京诺唯赞生物科技有限公司,RT-PCR试剂购自生工生物工程(上海)有限公司合成,高保真Taq酶购自MBIFermentas,pEASY-T1载体购自全式金生物技术有限公司。
1.2 试验方法
1.2.1 MFC基因克隆 将福寿螺胃组织获得后提取RNA,按照1stcDNA合成试剂盒说明书合成cDNA,使用引物对(mfcF1:5′-TCGACGACGCTTCAGTCAAGC-3′,mfcR1:5′-GTTGCCCTCTGAGTGTCGCTC-3′)通过高保真Taq酶PCR扩增MFC基因序列,纯化扩增片段并两端加“A”后将扩增片段连接到pEASY-T1载体,转化质粒到DH5α感受态细胞后挑单克隆送生工生物工程(上海)有限公司测序。
1.2.2 MFC基因序列分析 将测试获得的MFC基因序列与GenBank登录的近源序列EGXA (FJ183727.1,Ampullaria crossean)、MFC(EU5 99577.1,Ampullaria crossean)、EGX1(DQ848667.1,Pomacea canaliculata)、xylanase(AY941794.1,Ampullaria crossean、EGX3(DQ848668.1,Pomacea canaliculata cellulase)、EGX1(DQ848670.1,Pomacea
canaliculata)、EGX3(DQ848669.1,Pomacea canaliculata)进行序列比对,采用MEGA4软件对GenBank中近缘基因序列与实验测定MFC序列进行比对,构建系统发育树。
1.2.3 密码子偏好性分析 根据Paul和Elizabeth (1991)筛选原则[9](一条以ATG为起始密码子,以TAA、TAG或TGA为终止密码子且长度大于300 bp的完整CDS序列),选取德阳什邡毛木耳栽培基地采集的黄耳10号子实体样品测序获得的CDS (comp22688-c0-seq,环三磷酸核苷水解酶蛋白)为毛木耳测试样本,以克隆获得的MFC基因为福寿螺测试样本,运用CodonW软件计算福寿螺及毛木耳密码子使用次数和同义密码子相对使用度。
1.2.4 MFC基因毛木耳适应性密码子优化 利用同义密码子置换的方法,达到破坏mRNA的核糖体结合位点及翻译起始位点的茎环结构,得到具有最佳RNA二级结构及自由能的序列[10],并分析MFC基因改造前后的序列差异。
2 结果与分析
2.1 MFC基因测序及分析
以CTAB法提取的MFC基因组DNA为模板,扩增、克隆测序得到1088 bp的MFC基因序列。该序列GenBank受理号为KF636134.1,基因序列见图1。
2.2 MFC基因与近源基因系统发育分析
将福寿螺MFC基因序列与近缘基因EGXA (FJ183727.1,Ampullaria crossean)、MFC(EU599 577.1,Ampullaria crossean)、EGX1(DQ848667.1,Pomacea canaliculata)、xylanase(AY941794.1,Ampullaria crossean、EGX3(DQ848668.1,Pomacea canaliculata)、EGX1(DQ848670.1,Pomacea canaliculata)、EGX3(DQ848669.1,Pomacea canaliculata)进行序列比对。采用MEGA4软件进行比对,构建系统发育树(图2)。
通过比较发现实验获得MFC基因与其他近源基因存在差异,差异最小的是福寿螺来源的EGXA (FJ183727.1)和MFC(EU599577.1),遗传距离分别为0.006和0.007;差异最大的是同样来源的EGX1(DQ848670.1)及EGX3(DQ848669.1),遗传距离分别为0.799和0.816。
2.3 福寿螺MFC序列与毛木耳转录本序列密码子偏好性分析
运用CodonW软件计算MFC基因序列及毛木耳转录组测序获得CDS序列的密码子使用次数和同义密码子相对使用度。

图1 福寿螺多功能纤维素酶基因(MFC)序列Fig.1 Sequences of MFC gene from Ampullaria crossean

图2 福寿螺MFC基因与其它近源序列的系统发育树Fig.2 Phylogenetic tree based on MFC and closely related genes of other species
运用CodonW软件分析了MFC基因的密码子(共计396个)使用次数和同义密码子相对使用度(表1~2)。其中RSCU>1的密码子共计21个,为福寿螺MFC基因的偏好性密码子,其中CUG、GUG、AGC、AGA这4个密码子RSCU≥2,偏好性较强。而毛木耳CDS(comp22688-c0-seq,环三磷酸核苷水解酶蛋白)共4996密码子,RSCU>1的密码子共计26个,其中RSCU≥2密码子有5个,分别为CUC、AUC、GUC、CGC、GGC,偏好性较强。
总体上,福寿螺与毛木耳偏好密码子不同的氨基酸有10个(不包含起始密码子及终止密码子),相同偏好密码子的氨基酸有11个,不同偏好性密码子约占半数。其中RSCU≥2密码子中,福寿螺MFC基因有4个,毛木耳CDS有5个。其中4个氨基酸的密码子偏好性最强,分别是亮氨酸(Leu)、缬氨酸(Val)、丝氨酸(Ser)、精氨酸(Arg),并且偏好的密码子都各不相同,毛木耳还多一个偏好性较强的异亮氨酸(Ile)。
2.4 MFC基因密码子偏好性改造
利用多次同义密码子置换,排除含有毛木耳CDS中利用率低于10%的密码子以及存在反向重复碱基的候选序列,优化得到具有最佳RNA二级结构及自由能的序列(图3)。
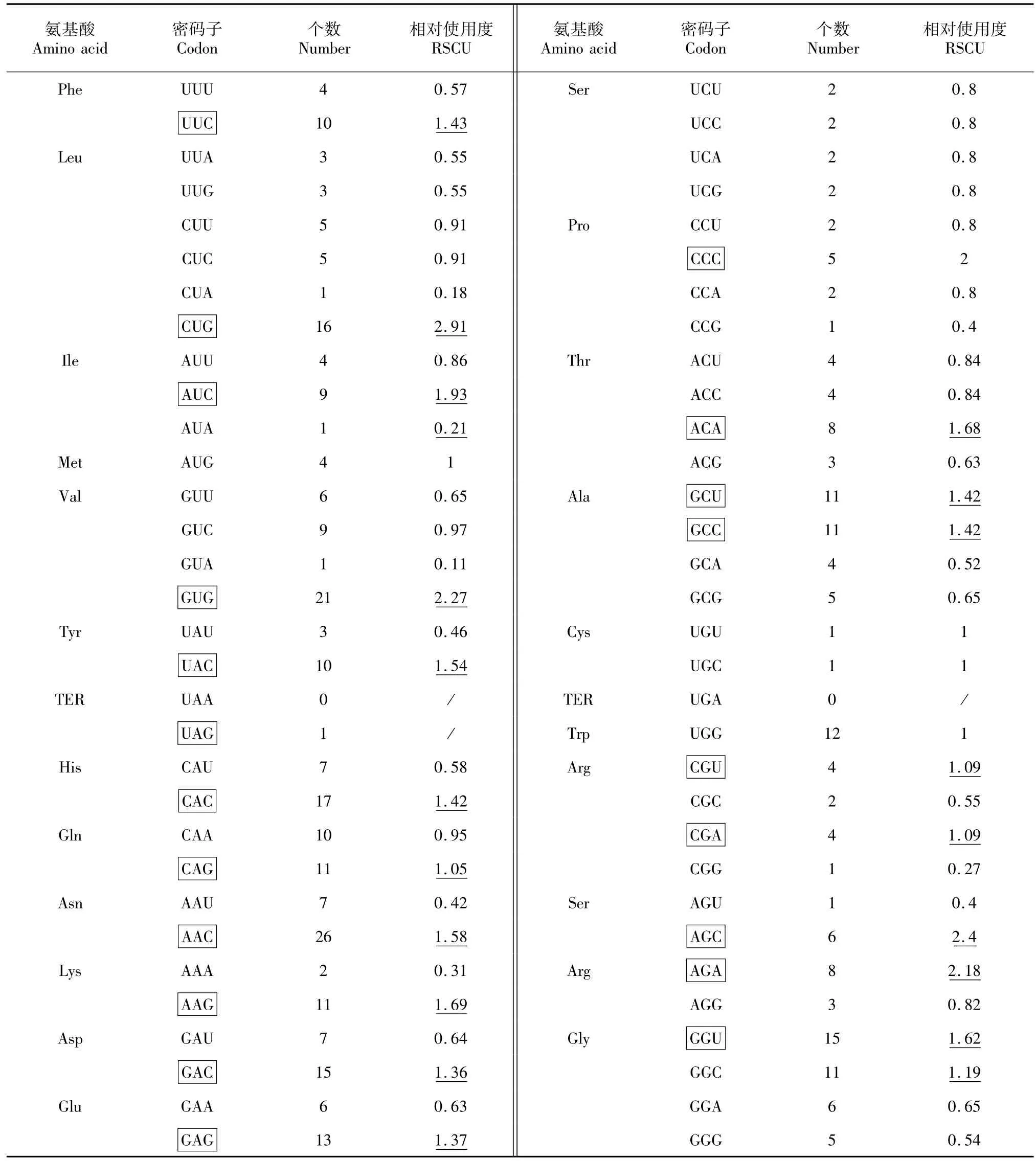
表1 MFC基因密码子偏好性分析Table 1 Codon usage of MFC gene in Ampullaria crossean
优化后的序列GC含量从原始MFC序列的54. 04%增加到58.16%,与毛木耳遗传背景一致。用DNAstar7.1.0软件检测后发现优化后的MFC序列最小自由能为-309.35 Kcal/mol,没有发夹结构、重复序列及反向重复序列(图4),序列优化效果良好。
3 讨 论
本实验分析结果显示福寿螺与毛木耳在氨基酸密码子偏好性上存在明显差异,如果不经密码子优化直接转入毛木耳,可能导致转化后该基因形成mRNA在宿主细胞内不能稳定存在[8]。在翻译上,如果福寿螺的基因密码子在毛木耳细胞内刚好是稀有密码子,这类问题会导致该基因不能得到较好翻译,从而影响该基因高效表达[11]。如本试验中分析发现毛木耳就存在UUA、CUA、AUA、GUA等稀有密码子,如果不排除存在这些含有毛木耳稀有密码子的序列,那么转导进入的毛木耳细胞的外源基因表达就会受到影响,不能实现实验预期目的。排除了稀有密码子、重复序列等不良影响因素设计的序列将是外源基因转导进入宿主细胞表达的最佳材料。本实验优化的MFC基因片段与PPK2连接构建农杆菌转化载体,通过AGL-1工程菌转导进入毛木耳细胞,并得到了良好表达(结果另文发表),这也是对密码子优化型基因高效表达的实验验证。
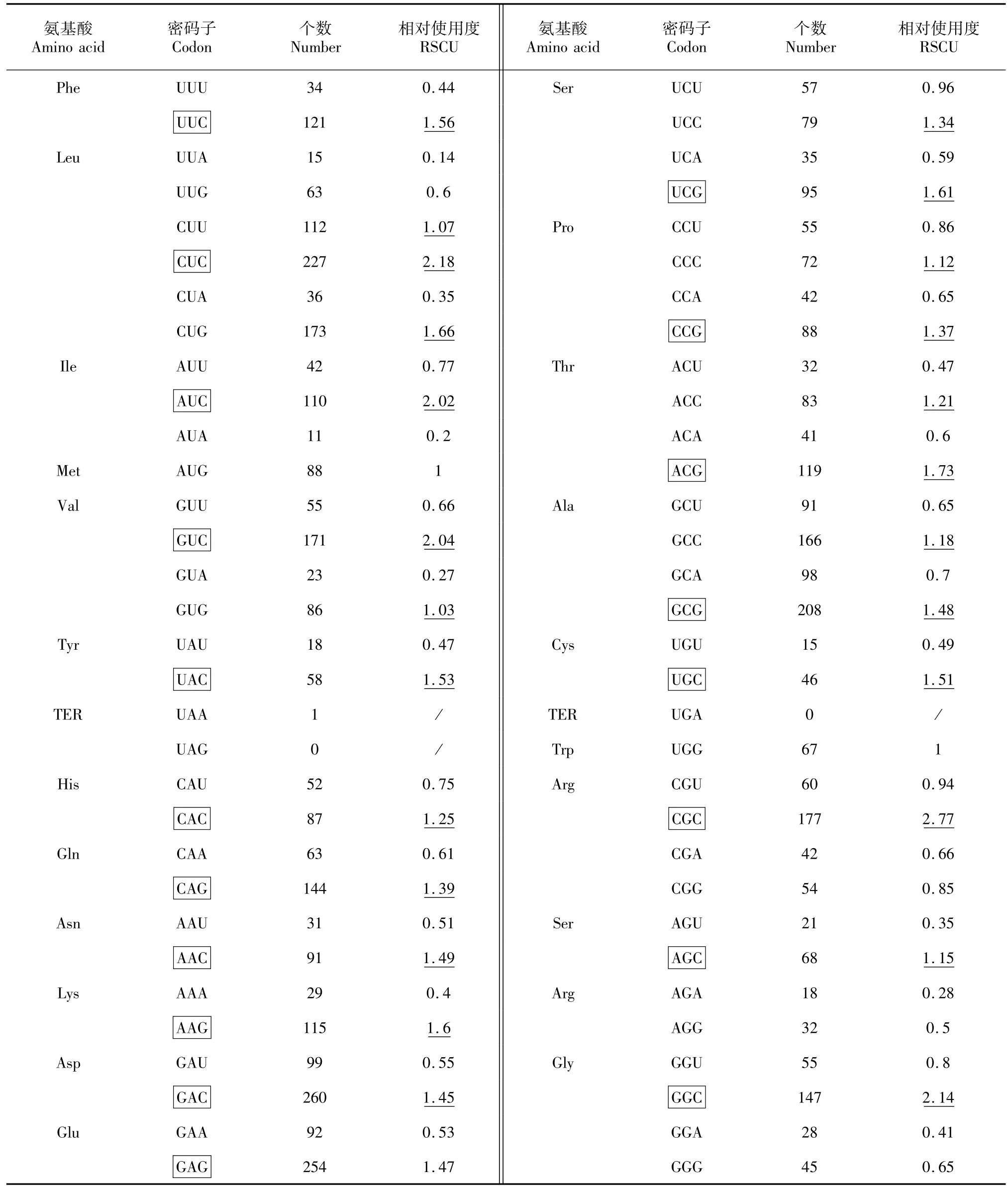
表2 毛木耳CDS密码子偏好性分析Table 2 Codon usage of CDS in Auricularia polytricha
实验将福寿螺基因MFC与毛木耳转录本CDS密码子偏好性进行了比较分析,并依据毛木耳密码子偏好性特征优化获得了改造型MFC序列,并在毛木耳细胞中得到了较好表达。实验结果为毛木耳外源基因改造和表达提供了参考,表明外源基因的密码子优化改造是实现异源基因在宿主细胞高效表达的重要手段。

图3 优化型多功能纤维素酶基因(MFC)序列Fig.3 Codon-optimized sequence of MFC gene
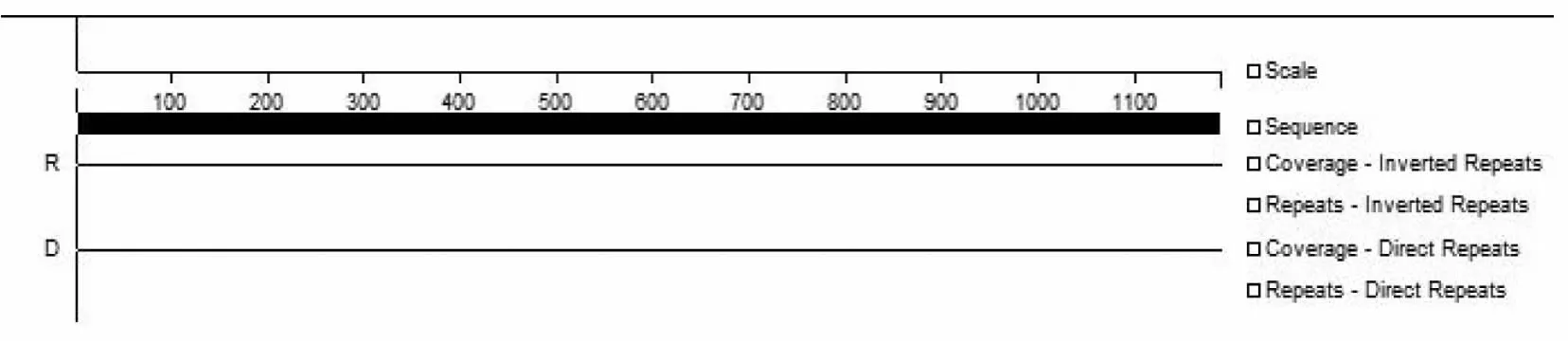
图4 优化型多功能纤维素酶基因(MFC)序列重复序列检测图Fig.4 Determine of repeats in the codon-optimized sequences of MFC gene
[1]权鹏碧.中国能源发展趋势——煤炭清洁利用[J].低碳世界,2015(1):101-102.
[2]张玉卓,蒋文化,俞珠峰,等.世界能源发展趋势及对我国能源革命的启示[J].中国工程科学,2015,17(9):140-145.
[3]Yu G J,Yin Y L,Yu W H,etal.Proteome exploration to provide a resource for the investigation of Ganoderma lucidum[J].PLoSOne,2015,10(3):e119439.
[4]Cao G,Zou D,Zhang X,et al.Bioenergy and Biomass Utilization [J].Biomed Res Int,2015,2015:857568.
[5]杨培周,郭丽琼,王艺红,等.福寿螺(Ampullaria crossean)mfc基因的分子克隆和序列分析[J].中国食品学报,2008,8(4):21-27.
[6]杨培周.多功能纤维素酶基因(mfc)的克隆及其在灰盖鬼伞中的表达研究[D].华南农业大学,2008.
[7]任燕萍,姚正培,倪志勇,等.植物逆境相关基因密码子的偏好性分析[J].湖南农业科学,2014(9):12-16.
[8]Presnyak V,Alhusaini N,Chen Y,et al.Codon Optimality Is a Major Determinant ofmRNA Stability[J].Cell,2015,160(6):1111-1124.
[9]吴正常,王 靖,赵乔辉,等.猪脂多糖结合蛋白基因(LBP)的密码子偏好性分析[J].农业生物技术学报,2013,21(10):1135 -1144.
[10]彭静静.经密码子优化的耐热纤维素酶在大肠杆菌中的表达[J].江苏农业学报,2014,30(3):497-502.
[11]Tokuoka M,Tanaka M,Ono K,et al.Codon Optimization Increases Steady-State mRNA Levels in Aspergillus oryzae Heterologous Gene Expression[J].Applied and Environmental Microbiology,2008,74(21):6538-6546.
(责任编辑 陈 虹)
Codon Optim ization of M ulti-functional Cellulase Gene in Ampullaria crossean Based on Characteristic Codon Usage of CDS in Auricularia polytricha
JIA Ding-hong1,2,WANG Bo1,LIXiao-lin1,TANWei1,PENGWei-hong1*
(1.Soil and Fertilizer Institute,Sichuan Academy of Agricultural Sciences,Sichuan Chengdu 610066,China;2.Key Laboratory of Bio-resource and Bio-environment,College of Life Sciences,Sichuan University,Sichuan Chengdu 610064,China)
【Objective】For subsequent transgenic study onmulti-functional cellulase gene(Mfc)in Auricularia polytricha,its cloning and codon optimization were performed.【Method】The biosoftware of CodonW was used to analyze the differences of codon usage between M fc gene in Ampullaria crossean and CDS in A.polytricha,and codons of Mfc gene in A.crossean were optimized based on characteristic codon usage of CDS in A.polytricha.【Result】The GC contentof optimized Mfcsg sequence increased from 54.04%to58.16%,which was in accordance with A.polytricha genetically.【Conclusion】The GC content of optimized Mfcsg sequence was in accordance with A.polytricha genetically.No hairpins,direct repeats and inverted repeats were detected in the optimized Mfcsg sequence,which indicated that this sequence was able to be used as heterologous gene for the transgenic study of A.polytricha.
Ampullaria crossean;Auricularia polytricha;Codon optimization
S567.3+9
A
1001-4829(2017)8-1726-06
10.16213/j.cnki.scjas.2017.8.006
2016-03-22
四川省财政创新能力提升工程项目(2014QNJJ-012);突破性食用菌新品种选育、育种材料与方法创新项目“毛木耳抗病新品种选育研究”;国家食用菌产业技术体系(CARS-24);四川食用菌创新团队
贾定洪(1977-),男,博士,副研究员,主要从事食用菌分子生物学和遗传育种研究,jdhdragon@qq.com,*为通讯作者。
