甲基营养型芽孢杆菌(Bacillusmethylotrophicus)发酵产蛋白酶培养基的双响应面优化
2017-09-15翁佩芳周小敏高志中吴祖芳
曹 红,翁佩芳*,周小敏,高志中,吴祖芳,张 鑫
(1.宁波大学 海洋学院,应用海洋生物技术教育部重点实验室,浙江 宁波 315211;2.浙江兴业集团有限公司,浙江 舟山 316100)
甲基营养型芽孢杆菌(Bacillusmethylotrophicus)发酵产蛋白酶培养基的双响应面优化
曹 红1,翁佩芳1*,周小敏2,高志中2,吴祖芳1,张 鑫1
(1.宁波大学 海洋学院,应用海洋生物技术教育部重点实验室,浙江 宁波 315211;2.浙江兴业集团有限公司,浙江 舟山 316100)
对甲基营养型芽孢杆菌(BacillusmethylotrophicusKC790303.1)产蛋白酶的培养基组成进行优化,以达到菌株生长良好、蛋白酶活力提高的目的。在单因素试验的基础上通过双响应面优化确定发酵培养基各成分的配比。结果表明,发酵培养基的成分及其浓度(m/V)为:葡萄糖2.95%、大豆蛋白胨1.30%和CaCl20.20%,此时菌体生长状态良好,所产蛋白酶活力最终达到312.98 U/mL,较之前提高了21.5%。在摇瓶发酵条件下的培养基优化研究为工业发酵放大提供理论基础。
甲基营养型芽孢杆菌;培养基;蛋白酶;双响应面;优化
蛋白酶是催化蛋白质肽键水解的一类酶,被广泛应用于食品加工行业中[1]。海洋源蛋白酶因其特殊结构和功能,在极端环境下仍保留较高活性和稳定性,因此被广泛应用到鱼酱油发酵、肉类加工等领域[2-3]。因此,在食品加工行业中,经常通过添加海洋蛋白酶改善食品的风味及营养价值。海洋微生物具有资源广泛、生长周期短、耐高压、耐高盐等优点,关于产蛋白酶海洋微生物的研究一直成为国内外的研究热点[4-5]。但目前市场开发的蛋白酶制剂成本较高,成为其广泛应用的限制因素[6]。因此,高效益产蛋白酶海洋源微生物的开发日益备受关注。
产蛋白酶微生物的发酵过程中受多种因素的影响,其中培养基组成是最关键的因素之一。为提高微生物发酵产酶效果,研究者通常优化微生物发酵的培养基成分和发酵条件。目前较多采用的是中心组合设计响应面优化分析(Response Surface Methodology,RSM),较常用的有Box-Behnken设计(Box-Behnken Design,BBD)和中心组合设计(Central Composite Design,CCD)[7-8]。双响应面是指将包含两个响应值的优化方法,实际上响应值也可以多个,国内对于双响应面的研究较少,而国外较多学者将此方法运用到菌株的发酵优化取得了满意的效果[9-10]。孙晓鸣等对产中性蛋白酶芽孢杆菌的培养基进行响应面优化,确定了培养基的最佳组成使其蛋白酶活力提高了42%[11]。Oskouie等对Bacillusclausii的培养基响应面优化使最终产碱性蛋白酶活力提高了近1.5倍[12]。
甲基营养型芽孢杆菌(Bacillusmethylotrophicus)是一种能够利用C—C键低碳化合物的微生物,广泛存在于土壤、水、空气以及动物肠道等处[13-14],具有广泛的商业价值,可以用来生产蛋白酶、多糖等,还可以作为抑菌剂,抵抗外界有害因子[15-17]。本文采用单因素和双响应面优化相结合的方法,对前期筛选出到的一株海洋来源产蛋白酶菌株—甲基营养型芽孢杆菌(KC790303.1)的培养基组成进行优化,旨在使菌株生长状态良好的前提下,能够持续稳定地产生具有较高酶活力的蛋白酶,为工业化应用奠定基础。
1 材料与方法
1.1 材料和试剂
1.1.1 菌株及培养基
菌株:甲基营养型芽孢杆菌(BacillusmethylotrophicusKC790303.1),本实验室保存。
基础发酵培养基(g/L):蛋白胨10 g,牛肉膏3 g,氯化钠5 g,121 ℃下灭菌15 min。
斜面保藏培养(g/L):蛋白胨10 g,牛肉膏3 g,氯化钠5 g,琼脂15 g ,121℃下灭菌15 min。
1.1.2 主要试剂
葡萄糖、蔗糖、麦芽糖、淀粉、糊精、酵母浸膏、NH4Cl、NaNO3、蛋白胨、胰蛋白胨、大豆蛋白胨、牛肉膏、ZnSO4、CaCl2、MnCl2、KH2PO4、CuSO4、MgCl2、酪蛋白、L-酪氨酸,以上试剂均为分析纯,国药集团化学试剂有限公司。
1.2 仪器与设备
QYC-2102C恒温振荡培养箱(宁波江南仪器厂); LV-3300分光光度计(上海美谱达仪器有限公司);5804R高速冷冻离心机(德国Eppendorf公司);5814R小型高速离心机(德国Eppendorf公司)。
1.3 培养方法
1.3.1 菌株活化
挑取斜面保藏的甲基营养型芽孢杆菌,接种到50 mL基础发酵培养基中,放置于恒温摇床中,30 ℃、150 r/min下活化24 h,此时的发酵液即为种子发酵液。
1.3.2 粗酶液的制备
将上述活化的种子发酵液以4%的接种量接种到发酵培养基中,30 ℃、150 r/min发酵48 h。发酵结束后的菌悬液离心(6 000 r/min,4 ℃)15 min,收集上清液即为粗酶液,每组三个平行,测定蛋白酶活力。
1.4 蛋白酶活力的测定
采用福林酚法(GB/T 23527—2009)测定蛋白酶活力。
1.5 菌株生长量测定
随着微生物生长的进行,微生物利用培养基中的营养物质,菌体不断增加,培养基的状态由澄清变为浑浊,培养基的浊度与微生物的生长量呈线性正比关系,因此采用比浊法在600 nm下进行OD值的测定,反映微生物生长量[18-19]。
将发酵液在4℃,6 000 r/min条件下离心15 min,收集下层菌体,加入等体积的蒸馏水震荡均匀,采用相同的方法处理未接种的培养基为空白对照,采用分光光度法测定600 nm时OD值。若菌体浓度太高可以将菌体稀释,并保持菌体的稀释倍数一致,一般OD值控制在0~2.5范围内[10,20]。
1.6 培养基组成的单因素优化
1.6.1 最佳碳源的确定及其浓度对菌株发酵的影响
选择葡萄糖、蔗糖、麦芽糖、淀粉、糊精和酵母浸膏六种不同类型的碳源,以1%(m/V,下同)添加量添加到基础发酵培养基中,不改变培养基其他成分,以基础发酵培养基为空白对照,发酵结束后,测定菌株生长量和粗酶液酶活力,单因素选择最佳碳源。
选择最佳碳源浓度为0.50%、1.00%、1.50%、2.00%、2.50%、3.00%、3.50%、4.00%,同样的条件下发酵培养,每组三个平行,测定菌株生长量和粗酶液酶活力,选出最佳碳源浓度。
1.6.2 最佳氮源的确定及其浓度对菌株发酵的影响
选择NH4Cl、NaNO3、蛋白胨、胰蛋白胨、大豆蛋白胨和牛肉膏六种不同类型的氮源,无机氮源和有机氮源分别以2%、1%(m/V)添加量添加到基础发酵培养基中,不改变培养基其他成分,以基础发酵培养基为空白对照,发酵结束后,测定菌株生长量和粗酶液酶活力,单因素选择最佳碳源。
选择最佳氮源浓度为0.50%、1.00%、1.50%、2.00%、2.50%、3.00%、3.50%、4.00%,同样的条件下发酵培养,每组三个平行,测定菌株生长量和粗酶液酶活力,选出最佳氮源浓度。
1.6.3 最佳无机盐及其浓度对菌株发酵影响
分别以ZnSO4、CaCl2、MnCl2、 MgCl2、CuSO4和KH2PO4为发酵培养基的唯一无机盐,添加量为0.01%,不改变培养基其他组分和发酵条件,以基础发酵培养基为空白对照,发酵结束后,测定菌株生长量和粗酶液酶活力,单因素选择最佳无机盐。
选择最佳无机盐浓度为0.05%、0.10%、0.15%、0.20%、0.25%、0.30%,同样的条件下发酵培养,每组三个平行,测定菌株生长量和粗酶液酶活力,选出最佳无机盐浓度。
1.7 培养基组成的双响应面优化
1.7.1 响应面优化试验设计
在单因素试验基础上,依据Box-Behnken响应曲面法(BBD)进行分析。以最优碳源、氮源、无机盐为自变量,菌株生长量和粗酶液酶活力为响应值。实验结果利用统计软件Design-Expert 8.0.5进行数据分析。所有实验值都是三次实验的平均值,数据以“平均值±标准差”表示[21-22]。
1.7.2 验证试验
对于拟合模型是否显著,需追加试验进行验证,即按照理论中的最佳配方进行试验,并分析实际值与预测值间的相关性,最终确定优化的结果。
1.8 摇瓶发酵曲线的测定
种子液以4%的添加量接种在优化后的液体培养基中,空白培养基对照,重复实验3次,以2 h为一个间隔,取发酵液5 mL迅速测定菌株生长量和产蛋白酶能力[23]。
2 结果与分析
2.1 单因素优化培养基组成
2.1.1 最佳碳源的确定及其浓度对发酵的影响
碳源是微生物正常生长不可缺少的主要营养成分之一,在微生物生长代谢过程中不仅为细胞提供基本的物质基础,参与细胞结构的组成,又是提供微生物生命活动中所需能源的原料[24]。选择6种不同的碳源,探究碳源种类对菌株发酵结果的影响,实验结果如图1所示。由图1可知该菌株的最佳碳源为葡萄糖,菌株生长量和酶活力均达到最高,其次为蔗糖和酵母浸膏,糊精最低,因此选择葡萄糖为最佳碳源。曹龙奎等对枯草芽孢杆菌产碱性蛋白酶培养基的优化中也发现其最佳碳源为葡萄糖[20],与报道结果一致。这是因为葡萄糖作为碳源时可被菌体直接利用,而其它碳源需要被分解为单糖时才能利用。

图1 碳源种类对菌株发酵影响
通过添加不同浓度的葡萄糖,不改变其他条件,测定菌株生长量和蛋白酶酶活力,结果如图2所示。从图2中可以看出菌株生长量和粗酶液酶活力随葡萄糖浓度的变化有显著变化,葡萄糖浓度过低或过高,菌体生长缓慢,其产蛋白酶活力也较低。当葡萄糖添加量为2.50%时菌体生长量达到最高,当葡萄糖添加量为3.00%时产蛋白酶活力最高,且菌体生长趋于稳定。综合考虑均菌株生长量和其粗酶液酶活力这两个指标,最终确定葡萄糖的最佳添加量为3.00%。研究表明,葡萄糖作为最简单的单糖,适宜浓度的葡萄糖很容易被菌体快速吸收利用,但当葡萄糖浓度过高时会抑制蛋白酶产生[25-26]。

图2 葡萄糖浓度对菌株发酵的影响
2.1.2 氮源种类及浓度对菌株发酵影响
氮源也是微生物正常生长过程中十分重要的营养来源,可以用来合成菌体生长代谢过程中的代谢中间产物,例如核苷酸、氨基酸等。固定其他培养基成分和培养条件,加入有机氮源或无机氮源,测定菌体生长量和产蛋白酶活力,结果如图3所示。由图3可知该菌株的最佳氮源为大豆蛋白胨,菌株生长量和酶活力均达到最高,其他的氮源的效果较差,因此选择大豆蛋白胨为最佳氮源。这与曹龙奎等研究枯草芽孢杆菌生长培养基的优化中发现的当大豆蛋白胨为氮源时,其产碱性蛋白酶活力最高结果一致[20]。研究报道指出,有机氮营养丰富,更适合于芽孢杆菌的产酶,因无机氮源被快速代谢利用,抑制蛋白酶的合成[27]。

图3 氮源种类对菌株发酵的影响
不改变其他条件的前提下,向培养基中添加不同浓度的大豆蛋白胨,再次测定菌株生长量和其产蛋白酶能力,结果如图4所示。当大豆蛋白添加量为1.50%时,蛋白酶活力达到最高,菌体生长量也较高。氮源是产蛋白酶菌株的关键限制因素,适宜的氮源浓度对蛋白酶的产生很重要,氮源浓度过低,菌体生长缓慢,不能满足蛋白酶合成的需求;氮源浓度过高,虽然利于菌体生长,但营养过于丰富,使菌体生长旺盛,需要消耗较多的能源维持生存环境,不利于蛋白酶代谢产物的合成,因此1.50%为大豆蛋白胨的最佳添加量。
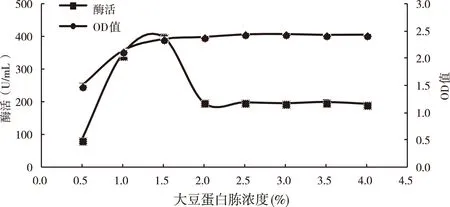
图4 大豆蛋白胨浓度对菌株发酵影响
2.1.3 无机盐种类及浓度对菌株发酵影响
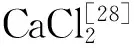
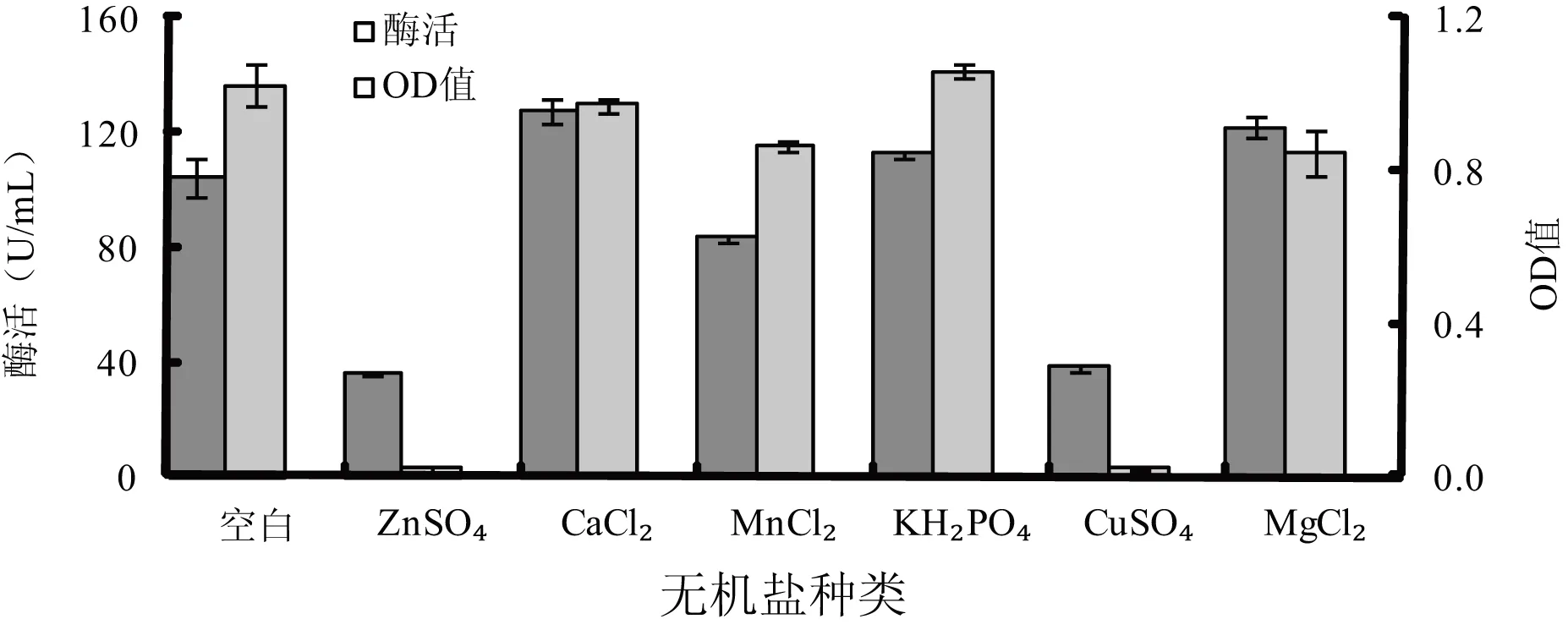
图5 无机盐种类对菌株发酵影响
不改变其他条件的情况下,添加不同浓度的CaCl2,再次测定菌株生长量及其产蛋白酶能力,结果如图6所示。由图6看出CaCl2的添加量为0.20%时,菌体生长良好,产蛋白酶能力最高。因此确定CaCl2的最佳浓度为0.20%。

图6 CaCl2浓度对菌株发酵的影响
上述实验数据表明单因素优化后该菌株的培养基组成为(m/V):葡萄糖3.00%,大豆蛋白胨1.50%和CaCl20.20%。
2.2 培养基组成双响应面模型的建立及其显著性检验
在单因素优化实验的基础上,采用Box-Behnken(BBD)原理,通过软件设计了3因素3水平的实验。葡萄糖(X1)、大豆蛋白胨(X2)、CaCl2(X3)三者的浓度为实验自变量,以菌株生长量(Y1)和酶活力(Y2)为实验响应值,共确定了17组实验。自变量因素编码及水平见表1,双响应面设计方案及实验数据结果分析见表2。通过本实验不仅可以使菌株达到最佳生长状态,而且菌株所产蛋白酶有较高的酶活力。

表1 中心组合设计自变量因素与水平

表2 双响应面试验设计方案与结果
2.2.1 菌株长量的模型建立及其显著性检验
利用软件Design-Expert8.0.5对表2中的实验数据进行拟合多元回归分析,得到菌株生长量的带交互项和平方项的二次多元回归预测方程,如下所示:
Y1=1.94+0.16X1-18.18X2-0.01X3+0.14X1X2-8.33X1X3-0.011X2X3+0.098X12+0.099X22+ 0.072X32
该模型的方差分析如表3所示,由表中可以看出该模型具有非常高的F回归值(281.70)和非常低的P值(P<0.000 1),表明该模型是高度显著的;而较低的F失拟值(5.35),失拟项P=0.0695>0.05,说明失拟不显著;RAdj2=0.927 2,说明该模型能够较好的模拟实验;R2=0.933 7,说明实验值与预测值之间有着良好的相关性。该模型实验误差小,拟合程度良好,所以可用于对菌株生长量进行分析和预测。此外,该模型中X1、X1X2、X12、X22、X32影响极显著。
2.2.2 菌株产蛋白酶活力的模型建立及其显著性检验
对表2中的数据拟合二次多元回归方程,葡萄糖、大豆蛋白胨、CaCl2浓度与菌株产蛋白酶活力的预测方程如下所示:
Y2=295.30+24.95X1-70.55X2-15.28X3+8.61X1X2-9.77X1X3+0.24X2X3-69.26X12-35.35X22- 52.71X32
从表3中可以看出该模型的F回归值(4989.76)和P值(P<0.000 1)表明该模型极显著;较小的F失拟值(3.11),失拟项P=0.150 8>0.05,说明失拟不显著;RAdj2=0.978 2,说明该模型能非常好的模拟实验,能够解释97.82%响应值变化;R2=0.992 8,说明实验值与预测值之间有着良好的相关性。该模型实验误差小,拟合程度良好,所以该模型可用于对菌株产蛋白酶活力进行分析和预测。此外模型中X1、X2、X3、X1X2、X1X3、X12、X22和X32影响极显著。
2.3 菌株培养基组成的双响应曲面分析及其优化
利用软件Design-Expert 8.0.5绘制葡萄糖浓度、大豆蛋白胨浓度、CaCl2浓度这三个因素之间的响应面分析图和相应的等高曲线图。在响应面分析图中,响应曲面坡度越大,其影响也越大,响应值也越大,在等高曲线图中,椭圆程度越高表示两因素间的交互作用越显著,相反,越接近于圆形表示两因素间的交互作用越不显著[31]。
2.3.1 菌株生长量的响应曲面分析及其优化
由表3的数据和图7可以看出,葡萄糖浓度和大豆蛋白胨浓度的P< 0.000 1、等高线图为椭圆,说明两者的交互作用显著。葡萄糖浓度和CaCl2浓度以及大豆蛋白胨浓度和CaCl2浓度的P﹥0.05,等高线图接近圆形,它们交互作用不显著。此外,通过响应曲面的等高曲线预测图可以得到菌株生长良好的培养基成分为:葡萄糖浓度2.94%、大豆蛋白胨浓度1.32%和CaCl2浓度0.20%,考虑到实际操作方便,最终将菌株生长的最佳培养基组成设定为葡萄糖、大豆蛋白胨和CaCl2的浓度分别为2.95%、1.30%和0.20%。在细菌生长过程中,碳源、氮源是培养基成分的关键因素,而无机盐是调节细菌生长代谢的重要组成成分,因此培养基的组成是细菌生长的基础。
2.3.2 菌株产蛋白酶活力的响应曲面分析及其优化
由表3的数据和图8可以看出,葡萄糖浓度和大豆蛋白胨浓度,葡萄糖浓度和CaCl2浓度的P< 0.000 1,它们的等高曲线图为椭圆,说明它们之间的交互作用显著。大豆蛋白胨浓度和CaCl2浓度的P﹥0.05,等高线图接近圆形,两者之间的交互作用不显著。此外,通过响应曲面的等高曲线预测图可以
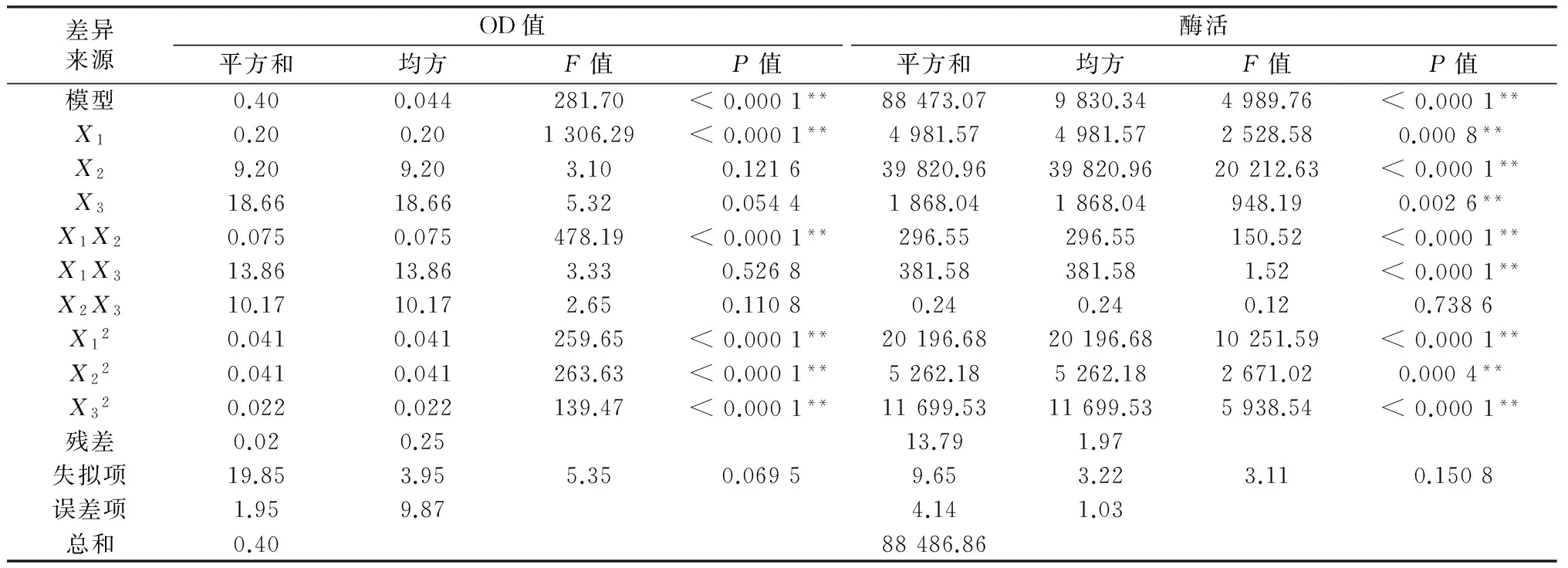
表3 回归模型的方差分析
注:**表示差异极显著(P<0.01)。

图7 各因素交互作用对菌株生长量的响应曲面图和等高曲线图

图8 各因素交互作用对菌株产蛋白酶活力的响应曲面图和等高曲线图
得到菌株产蛋白酶能力的最佳(312.98 U/mL)的培养基组成为:葡萄糖浓度2.94%、大豆蛋白胨浓度1.32%和CaCl2浓度0.20%,考虑到实际操作方便,最终将菌株产蛋白酶活力最佳培养基组成设定为葡萄糖、大豆蛋白胨和CaCl2的浓度分别为2.95%、1.30%和0.20%。
2.3.3 培养基组成双响应面模型验证试验
通过比较可以发现可以利用同样的预测条件下,菌株不仅生长良好,而且所产蛋白酶活力最高。因此最佳培养基发酵组成为:葡萄糖2.95%、大豆蛋白胨1.30%和CaCl20.20%,该模型OD值和酶活的最大响应值分别为1.95和312.98 U/mL。为了验证模型的准确性,在优化的培养基组成下进行10组验证实验(均选择在实验的范围之内,5组实验按照得到的最优条件进行。另外5组随机选择)。在10组验证实验中,实验值是预测值的98.73%,两者的良好拟合性证实了模型的有效性,可见该模型能良好的预测实际发酵情况。在最优发酵条件下蛋白酶活力比未优化的蛋白酶活力(257.67 U/mL)提高了21.5%。
2.4 菌株摇瓶发酵曲线的测定
采用双响应面优化培养基配方,在摇瓶发酵条件下,得到该菌株的发酵过程曲线,如图9。在整个发酵过程中,在0~22 h菌体处于对数生长期,此时的酶活力相对较低,发酵22~36 h菌体达到稳定期,此时酶活力继续不断升高,直到32 h时菌体所产酶活力基本稳定。

图9 菌株摇瓶发酵曲线
摇瓶发酵曲线可为放大发酵提供数据基础,从摇瓶发酵曲线,可以看出该菌株的产蛋白酶周期较短,在工业发酵中,通过进一步优化发酵条件酶活力可进一步提高。
3 结 论
本研究采用甲基营养型芽孢杆菌(Bacillusmethylotrophicus)为发酵菌株,通过单因素和双响应面实验优化其发酵培养基的碳源、氮源和无机盐,确定了菌株发酵的培养基组成。当葡萄糖、大豆蛋白胨和CaCl2的浓度(m/V)分别为2.95%、1.30%和0.20%时,发酵菌株生长状态良好,产蛋白酶活力高达312.98 U/mL,较为优化前提高了21.5%。验证试验与预测值接近,证明了优化结果的可靠性。在此基础上摇瓶发酵曲线的测定为工业发酵生产提供了必要的理论基础。
[1] OSKOUIE S F G, TABANDEH F, YAKHCHALI B, et al. Response surface optimization of medium composition for alkaline protease production byBacillusclausii[J]. Biochemical Engineering Journal, 2008, 39(1): 37-42.
[2] HE H, CHEN X, LI J, et al. Taste improvement of refrigerated meat treated with cold-adapted protease[J]. Food Chemistry, 2004, 84(2):307-311.
[3] AKOLKAR A V, DURAI D, DESAI A J.Halobacteriumsp. SP1 (1) as a starter culture for accelerating fish sauce fermentation[J]. Journal of Applied Microbiology, 2010, 109(1): 44-53.
[4] KIM E H, CHO K H, LEE Y M, et al. Diversity of cold-active protease-producing bacteria from arctic terrestrial and marine environments revealed by enrichment culture[J]. The Journal of Microbiology, 2010, 48(4): 426-432.
[5] FULZELE R, DESA E, YADAV A, et al. Characterization of novel extracellular protease produced by marine bacterial isolate from the Indian Ocean[J]. Brazilian Journal of Microbiology, 2011, 42(4): 1364-1373.
[6] LI Z Y, YOURAVONG W, ARAN H. Protein hydrolysis by protease isolated from tuna spleen by membrane filtration: a comparative study with commercial proteases[J]. LWT-Food Science and Technology, 2010, 43(1): 166-172.
[7] FAKHFAKH-ZOUARI N, HADDAR A, HMIDET N, et al. Application of statistical experimental design for optimization of keratinases production byBacilluspumilusA1 grown on chicken feather and some biochemical properties[J]. Process Biochemistry, 2010, 45(5): 617-626.
[8] PAI C K, ZENG Y F, YUEH P Y, et al. Prediction of optimum reaction conditions for the thermos-tolerant acetylxylan esterase from Neocallimastix patriciarum using the response surface methodology[J]. Journal of Chemical Technology and Biotechnology, 2010, 85(5): 628-633.
[9] SINGH S K, SINGH S K, TRIPATHI V R, et al. Comparative one-factor-at-a-time, response surface (statistical) and bench-scale bioreactor level optimization of thermoalkaline protease production from a psychrotrophicPseudomonasputidaSKG-1 isolate[J]. Microbial Cell Factories, 2011, 10(1): 114.
[10] AVCI A, SAHA B C, DIEN B S, et al. Response surface optimization of corn stover pretreatment using dilute phosphoric acid for enzymatic hydrolysis and ethanol production[J]. Bioresource Technology, 2013, 130: 603-612.
[11] 孙晓鸣, 王萍, 邬向东. 产中性蛋白酶芽孢杆菌配伍发酵响应面法研究[J]. 中国食品学报, 2010 (2): 117-124.
[12] OSKOUIE S F G, TABANDEH F, YAKHCHALI B, et al. Response surface optimization of medium composition for alkaline protease production byBacillusclausii[J]. Biochemical Engineering Journal, 2008, 39(1): 37-42.
[13] ZHANG T, LI R, QIAN H, et al. Biosynthesis of levan by levansucrase fromBacillusmethylotrophicusSK 21.002[J]. Carbohydrate Polymers, 2014, 101: 975-981.
[14] 熊涛, 唐晓星, 黄涛, 等. 产蛋白酶兼性厌氧菌株的筛选, 酶学性质及发酵豆粕应用探究[J]. 食品科学, 2014, 35(9): 162-167.
[15] 吴燕燕, 张岩, 李来好, 等. 甲基营养型芽孢杆菌抗菌肽对罗非鱼片保鲜效果的研究[J]. 食品工业科技, 2013, 34(2): 315-318.
[16] XIE F, QUAN S, LIU D, et al. Purification and characterization of a novel α-amylase from a newly isolatedBacillusmethylotrophicusstrain P11-2[J]. Process Biochemistry, 2014, 49(1): 47-53.
[17] MALA J G S, SUJATHA D, Rose C. Inducible chromate reductase exhibiting extracellular activity inBacillusmethylotrophicusfor chromium bioremediation[J]. Microbiological Research, 2015, 170: 235-241.
[18] 牛天贵. 食品微生物学实验技术[M]. 北京:中国农业大学出版社, 2002.
[19] 蔡毅, 谷新晰, 田晶晶, 等. 乳酸菌摇瓶增殖培养工艺研究[J]. 中国食品学报, 2012, 12(4): 107-113.
[20] 曹龙奎, 吴泽柱, 盛艳. 枯草芽孢杆菌生长培养基的优化[J]. 中国食品学报, 2009, 9(6): 104-109.
[21] RAJ A, KHESS N, PUJARI N, et al. Enhancement of protease production by Pseudomonas aeruginosa isolated from dairy effluent sludge and determination of its fibrinolytic potential[J]. Asian Pacific Journal of Tropical Biomedicine, 2012, 2(3): S1845-
S1851.
[22] BADOEI-DALFARD A, KARAMI Z. Screening and isolation of an organic solvent tolerant-protease fromBacillussp. JER02: Activity optimization by response surface methodology[J]. Journal of Molecular Catalysis B: Enzymatic, 2013, 89: 15-23.
[23] 刘鹏, 王泽南, 李莹, 等. 丛梗孢酵母产赤藓糖醇的诱变选育[J]. 食品科学, 2011, 32(11): 216-221.
[24] 姚刚, 程建军, 孙鹏, 等. 枯草芽孢杆菌发酵产碱性蛋白酶的研究[J]. 食品科学, 2009 (23): 347-351.
[25] NADEEM M, QAZI J I, BAIG S, et al. Effect of medium composition on commercially important alkaline protease production byBacilluslicheniformisN-2[J]. Food Technol Biotechnol, 2008, 46(4): 388-394.
[26] SADEGHEIN B, ALAVI S A. Production of alkaline protease byBacilluslicheniformis[J]. Journal of Chemistry and Chemical Engineering, 2011, 5(2):105-109.
[27] ANNAMALAI N, RAJESWARI M V, SAHU S K, et al. Purification and characterization of solvent stable, alkaline protease fromBacillusfirmusCAS 7 by microbial conversion of marine wastes and molecular mechanism underlying solvent stability[J]. Process Biochemistry, 2014, 49(6): 1012-1019.
[28] CHI Z, MA C, WANG P, et al. Optimization of medium and cultivation conditions for alkaline protease production by the marine yeastAureobasidiumpullulans[J]. Bioresource Technology, 2007, 98(3): 534-538.
[29] BLUNT J W, COPP B R, KEYZERS R A, et al. Marine natural products[J]. Natural Product Reports, 2013, 30(2): 237-323.
[30] 王东, 荣家萍, 唐自钟, 等. 响应面法优化枯草芽孢杆菌产中性蛋白酶的发酵条件[J]. 基因组学与应用生物学, 2016, 35(1): 143-151.
[31] ZHOU X, XIN Z J, LU X H, et al. High efficiency degradation crude oil by a novel mutant irradiated fromDietziastrain by12C6+heavy ion using response surface methodology[J]. Bioresource Technology, 2013, 137: 386-393.
Dual Response Surface Optimization of Protease Producing Mediumby Fermentation ofBacillusmethylotrophicus
Cao Hong1, Weng Peifang1*, Zhou Xiaomin2, Gao Zhizhong2, Wu Zufang1, Zhang Xin1
(1.Key Laboratory of Applied Marine Biotechnology, School of Marine Sciences, Ningbo University,Ningbo 315211, China; 2.Xingye Group Co., Ltd., Zhoushan 316100, China)
In order to achieve a better strain with high protease activity, medium compositions for producing protease through the fermentation ofBacillusmethylotrophicuswere optimized by dual response surface design based on single factor experiments. The results showed that the composition and concentration (m/V)of fermentation medium were glucose 2.95%, soy peptone 1.30% and CaCl20.20%. Under the conditon, the bacteria grew better and the protease activity increased to 312.98 U/mL eventually, increasing by 21.5%. The optimizing culture medium of shaking flask fermentation provides a theoretical basis for larger industrial fermentation.
Bacillusmethylotrophicus; medium; protease; dual response surface; optimization
10.3969/j.issn.1006-9690.2017.04.003
2016-11-18
国家级星火计划(2015GA700089);“水产”浙江省重中之重学科开放基金(XKZSC1535)。
曹红(1989—), 女, 研究生, 主要研究方向: 主要从事水产品加工与高值化利用研究。E-mail:caohongnbu@163.com
*通讯作者: 翁佩芳(1963—), 女, 教授, 主要研究方向: 主要从事水产品加工与高值化利用研究。E-mail:weng-pf@163.com
TQ925+.2
A
1006-9690(2017)04-0009-08
