普通小球藻对不同浓度镉胁迫的生理应答
2017-09-12葸玉琴任春燕朱巧巧孔维宝孙对兄
葸玉琴 任春燕 朱巧巧 孔维宝 孙对兄
(西北师范大学, 兰州 730070)
普通小球藻对不同浓度镉胁迫的生理应答
葸玉琴 任春燕 朱巧巧 孔维宝 孙对兄
(西北师范大学, 兰州 730070)
以普通小球藻(Chlorella vulgaris)为研究材料探讨不同镉浓度对小球藻生理生化特性的影响。研究结果表明: 在5—40 μmol/L的Cd2+胁迫下, 普通小球藻的蛋白质、多糖和叶绿素含量均表现为随着Cd2+浓度的增加呈现逐渐增加的趋势, 30 μmol/L略高于对照组后又逐渐降低; 小球藻细胞中SOD活性及过氧化氢(H2O2)含量表现出先增加后减弱的趋势, 35 μmol/L时达到最大; 而小球藻中MDA含量随着Cd2+浓度的升高一直增加。这说明在重金属镉胁迫下小球藻会通过不同的生理应答来维持自身的正常生长代谢。
普通小球藻; 镉胁迫; 渗透调节物; MDA; 氧化酶
重金属镉是一种相对微量的元素, 由于其对水体环境中的生物, 如贝类、海带、海鲜等均有较高的毒性, 因而被认为是水体生态环境中危害最大的重金属之一[1,2]。通过生物富集作用, 这些重金属离子会在水环境生物中累积, 最终通过食物链的作用传递到人体, 严重危害人体健康。随着现代工业化步伐的加快, 废水的违规排放, 重金属污染水体已经达到不容忽视的程度, 我国首个“十二五”规划中就有提出水体重金属污染的防治问题[3], 更加显示出我国水体重金属污染治理的紧迫性和重要性。小球藻(Chlorella vulgaris)是水体中的初级生产者, 其生命周期比较短并且能快速的响应水体中生态环境的变化[4], 因此在水体环境变化中起重要的信号作用[5]。再者, 由于小球藻作为生物吸附的重要材料, 具有无二次污染, 易于培养等多种优点,因此, 小球藻可以作为研究重金属水体污染的较理想材料。国内许多的学者也已经进行了大量的研究[6—11]。
实验将普通小球藻作为研究材料, 研究镉(Cd2+)胁迫对其生长及其部分的生理生化的影响,如蛋白质、多糖、光合色素、丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化氢(H2O2)。初步探讨了小球藻对Cd2+的生理应答机制, 使其能更好地用于重金属水体污染的修复中, 为重金属水体污染的监测和防治提供有效的参考。
1 材料与方法
1.1 实验材料
实验所使用的藻种是购买自中国科学院淡水藻种库的普通小球藻。
1.2 培养方法
将小球藻接种到镉浓度分别为5、10、15、20、25、30、35和40 μmol/L的培养液中每个处理设3个平行, 以不添加Cd2+为对照组, 置于恒温摇床培养, 转速145 r/min, 光照强度3000 lx, 温度31℃, pH 6.0, 光暗比12h:12h, 每天测定各组的藻密度, 连续培养到第6天时测定生理指标。
1.3 方法
小球藻(Chlorella vulgaris)生长曲线的测定用浊度比色法[12]检测藻密度。蛋白质含量的测定用考马斯亮蓝法测定[13]。多糖含量的测定将蒽铜溶解到80%的H2SO4, 蒽铜-浓硫酸法测定[14]。叶绿素含量的测定参照三波长比色法[13]。丙二醛(MDA)含量的测定采用硫代巴比与妥酸法[15]。超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBT)法[16]测定。H2O2含量的测定参考Sergiev等[17]的方法。
2 结果与分析
2.1 不同浓度Cd2+对普通小球藻生物量曲线的影响
如图 1所示, 第1到第2天小球藻的繁殖速度普遍较高, 但在高Cd2+浓度35和40 μmol/L的胁迫下,小球藻的繁殖速度相比其他浓度的繁殖速度低; Cd2+浓度低于40 μmol/L时, 其他各处理组小球藻生长明显, 经过第3到第5天的生长, 小球藻的繁殖速度逐渐减慢, 第5到第6天时, 小球藻的繁殖速度逐渐降低, 其中40 μmol/L时下降最快, 藻出现大量死亡的现象。Cd2+浓度在30 μmol/L时一直呈现较快增长的趋势, 下降也较其他浓度的相对慢, 较对照组的小球藻生物量略高, 说明30 μmol/L的Cd2+浓度利于小球藻的生长, 对小球藻的生长起到了一定的促进作用。李双双[18]研究报道, 普通小球藻在96h与120h时的生长量逐渐趋于稳定。在Cd2+浓度低于30 μmol/L时对小球藻生长有促进作用, 随着浓度的不断增加则对小球藻的生长产生抑制作用,这是符合Hormesis效应的[19]。对小球藻生长的促进是由于低浓度的Cd2+对小球藻有“毒性兴奋效应”[20,21], 而抑制是由于高浓度的Cd2+阻碍了小球藻的正常生理代谢活动而使生长会变的迟缓[22]。
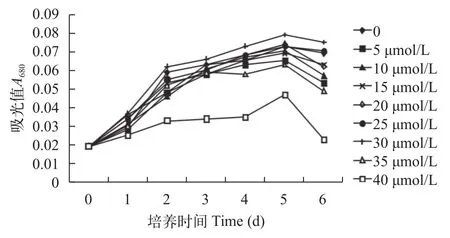
图 1 不同浓度Cd2+影响下普通小球藻的生长曲线
2.2 不同浓度Cd2+对普通小球藻蛋白质含量的影响
由图 2可以看出, 随着Cd2+浓度的增加, 各处理的蛋白质含量呈现高浓度抑制低浓度促进的趋势。从5—30 μmol/L, 各处理的蛋白质含量依次增加, 30—40 μmol/L, 蛋白质的含量依次降低, 在30 μmol/ L时, 蛋白质的含量最高, 且略高于对照组。不同浓度Cd2+对小球藻的蛋白质含量的影响也符合Hormesis效应[19], 说明蛋白质含量的变化与小球藻生物量的变化是相关的。在较低浓度Cd2+胁迫时, 小球藻的生长情况比较好, 其蛋白质合成量也较高, 而高浓度胁迫时, 由于阻碍了小球藻的生长, 其蛋白质的合成量也相应的减少。尹文珂等[23]研究得出, 四尾栅藻在较低Cd2+(0—1 mg/L)胁迫下, 其可溶性蛋白质的含量随Cd2+浓度的升高而增加, 当其含量达到最大量时, 会随着Cd2+浓度的升高其含量逐渐减少,可溶性蛋白质的合成受到抑制。有相关研究表明,蛋白质的合成和细胞内的多种代谢过程有关, 重金属离子的胁迫能诱导相关蛋白基因的表达, 从而减轻重金属对植物本身的毒害作用, 这是植物自身对环境的一种抵抗机制[24]。
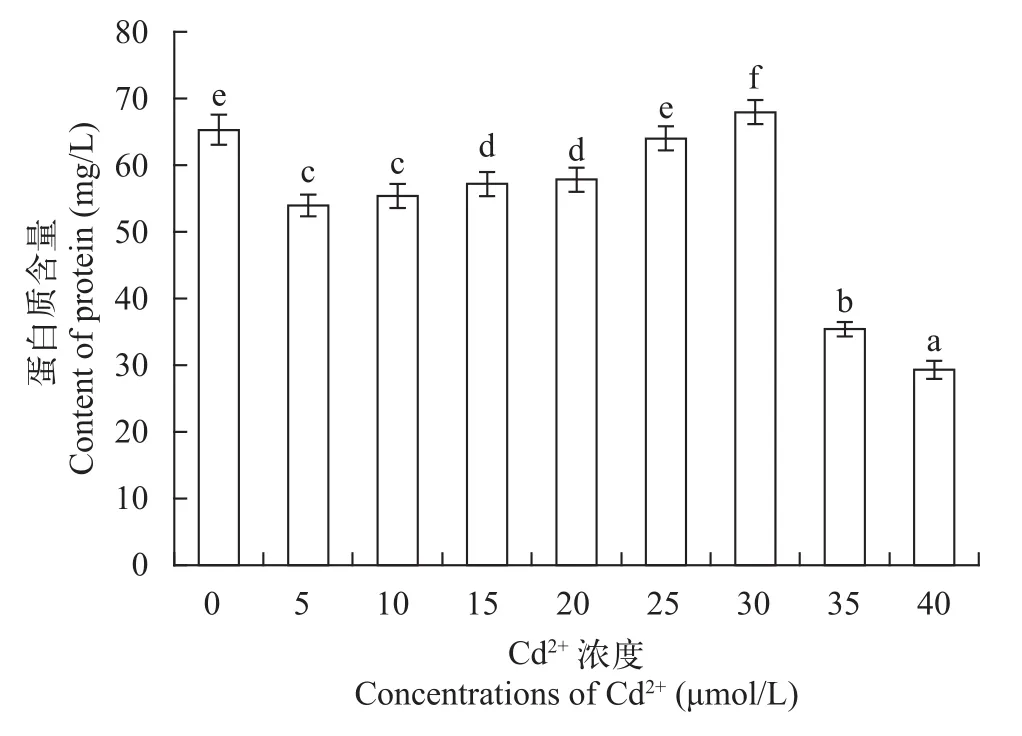
图 2 不同浓度Cd2+对普通小球藻蛋白质含量的影响
2.3 不同浓度Cd2+对普通小球藻多糖含量的影响
从图 3可知, 多糖含量在Cd2+浓度为5—30 μmol/L基本呈现逐渐增多的趋势, 在30—40 μmol/L内, 随着Cd2+胁迫的增强, 其可溶性多糖的含量逐渐减少,在30 μmol/L时, 可溶性多糖含量最高, 且略高于对照组, 比对照组高出23.44%。普通小球藻的多糖含量的变化趋势与和其生物量的变化类似。在低浓度Cd2+胁迫下, 其多糖的含量较高, 但低于对照组,而在高浓度的Cd2+胁迫下, 普通小球藻的生长状况下降, 其多糖的合成也受阻。因此低于30 μmol/L有利于多糖的合成。多糖作为渗透调节物, 是植物碳代谢的主要物质之一。有报道称, 某些植物受到来自环境方面的胁迫时, 可溶性多糖含量增加来调节渗透平衡以维持植物的正常代谢[25]。张学颖等[22]研究表明在重金属胁迫下, 盐藻的多糖含量是逐渐增加的, 当随着重金属浓度的增大且达到一定量时,多糖的含量则会明显的降低。多糖含量的这种变化, 是小球藻适应外界环境胁迫的重要机制之一。
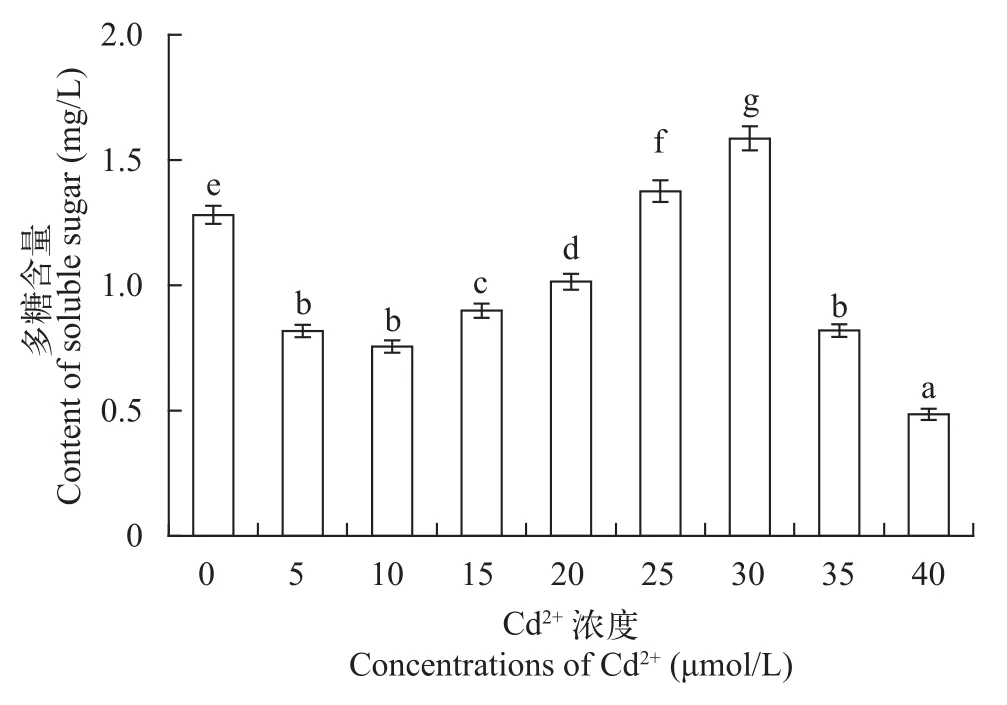
图 3 不同浓度Cd2+对普通小球藻多糖含量的影响
2.4 不同浓度Cd2+对普通小球藻叶绿素的影响
从图 4可以看出, 普通小球藻的叶绿素浓度很明显受到了Cd2+的影响。Cd2+浓度在5—30 μmol/L时叶绿素的浓度和Cd2+的浓度呈现显著的正相关,即Cd2+浓度较低时, 对小球藻的叶绿素含量有一定的促进作用, 但叶绿素含量仍然低于对照组。而在30 μmol/L时, 叶绿素的浓度最高, 且高于对照组7.41%。在40 μmol/L时, 叶绿素浓度比对照组下降了164.87%, 而比30 μmol/L时, 下降了184.48%。可见在高浓度Cd2+胁迫下, 对小球藻叶绿素含量的抑制作用十分显著, 叶绿色素含量下降较快。有研究报道, 在Cd2+胁迫下小球藻的叶绿素含量的降低,与Cd2+抑制了叶绿素酸酯还原酶和影响了氨基-γ-戊酮酸的合成有关系[26], 也有另一种可能, Cd2+直接破坏了叶绿体的结构和功能、干扰了植物对于营养物质的吸收和转移[27]。叶绿素含量的高低是反应植物生长状况的重要指标, 叶绿素较低则表明环境对藻类的生长有一定的抑制作用, 反之则有一定的促进作用[28]。有学者研究表明, Cd2+对轮藻的叶绿素含量也有一定的抑制作用, 会使其叶绿素含量有不同程度的下降[29]。
2.5 不同浓度Cd2+对普通小球藻MDA含量的影响
如图 5所示, MDA的含量随着Cd2+浓度的增加而增加, 其中各处理的MDA含量比对照组分别高出12.17%、23.48%、29.57、34.78%、46.96%、50.43%、55.65%和60.87%, 说明随着Cd2+浓度的增加, 对生物膜的毒害作用逐渐加强, 40 μmol/L的Cd2+浓度时, MDA的含量达到最大值, 说明普通小球藻在这个浓度下, 对小球藻的细胞膜产生严重的毒害作用。杨国远[30]的研究也表明, 在重金属Pb2+胁迫下的斜生栅藻, MDA含量也一直升高, 均高于对照组, 在第4天时, 已比对照组高出130.92%,对膜脂产生的非常严重的毒害。李福燕[31]的研究指出黑藻在Cd2+的毒害作用下, 使得黑藻细胞膜通透性增加, 其细胞膜系统受到明显的毒害作用, 导致其MDA含量进一步升高。应用SPSS 17.0软件对实验的数据进行了差异显著性的检验, 结果P<0.05,说明各处理间出现了显著差异。

图 4 不同浓度Cd2+对普通小球藻叶绿素含量的影响
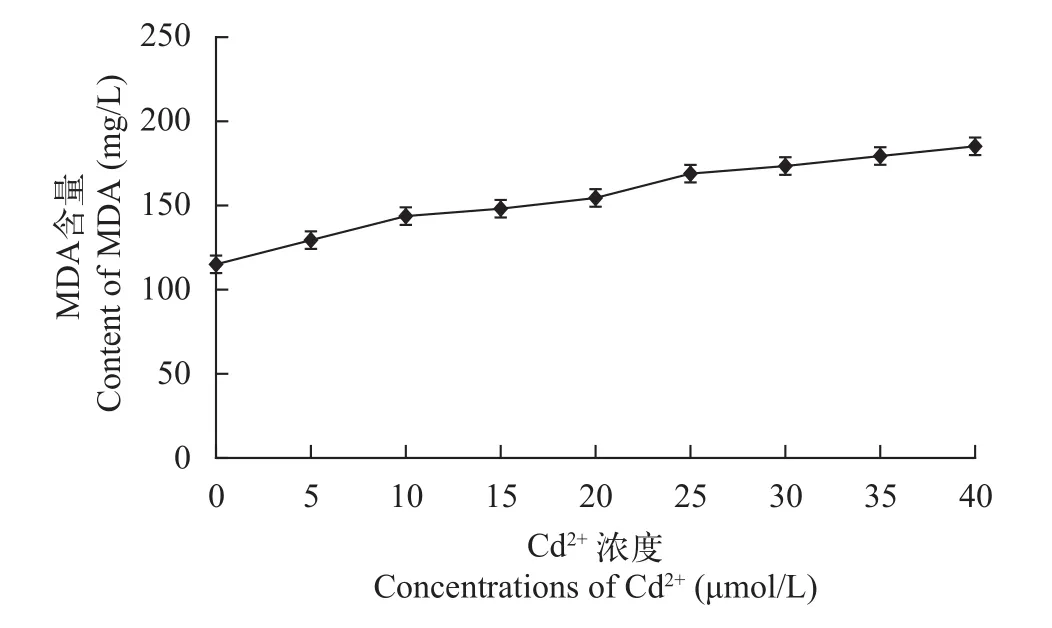
图 5 不同浓度Cd2+对普通小球藻MDA含量的影响
2.6 不同浓度Cd2+对普通小球藻SOD活性的影响
SOD在生物体中分布广泛, 几乎所有生物种类中都有SOD, 它是一种天然的自由基清除剂, 能清除生物体内新陈代谢过程中产生的毒害物质[32]。在图 6中, SOD的活性表现为随着Cd2+浓度的增加而缓慢升高, 15 μmol/L之后又快速升高, 在30 μmol/L以后又缓慢下降。其中Cd2+浓度为30 μmol/L时SOD含量是对照组的1.54倍, 普通小球藻中SOD活性的升高表示氧自由基产生速率的增加[33], 同时可以清除植物体内的自由基。30 μmol/L以后, SOD活性降低可能与小球藻细胞内MDA含量的积累有关,即MDA抑制了SOD的活性[34]。Rijstenbil等[35]研究了Zn2+与Cu2+对布氏双尾藻SOD活性变化的影响,结果显示藻细胞内的SOD活性显著增加, 表明微藻细胞内产生了大量的氧自由基。焦铁男和朱宏[36]研究了黑藻对重金属Cd2+的积累及其生理响应, 结果表明, SOD活性呈现先升后降的趋势, 其在一定浓度的重金属范围内, 对过氧化物有一定的清除作用, 这与黑藻对重金属Cd的抗性有关。应用SPSS 17.0软件对实验的数据进行了差异显著性的检验,结果P<0.05, 说明各处理间出现了显著差异。
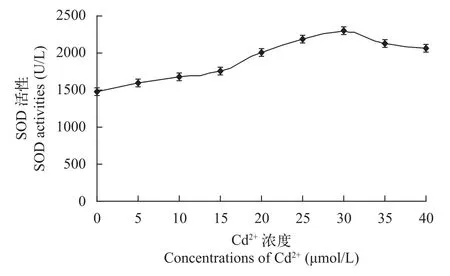
图 6 不同浓度Cd2+对普通小球藻SOD活性的影响
2.7 不同浓度Cd2+对普通小球藻H2O2含量的影响
过氧化氢(H2O2)是植物体内一种较为稳定的活性氧分子[37]。有很多证据表明, 植物在逆境中时, H2O2对植物的抗逆境活动有重要的作用[38]。如图 7所示, 随着Cd2+浓度的增加, H2O2的含量逐渐增加, 其含量在30 μmol/L时略显降低, 可能与SOD的活性有关, SOD能清除部分的H2O2, 防止植物细胞内的大分子的损伤, 如蛋白质和酶等物质[38], 而在30 μmol/L以后, H2O2的含量呈现一个幅度不大的增加后继而呈下降的趋势, 可能与活性氧(ROS)和SOD含量变化有关。应用SPSS 17.0软件对实验的数据进行了差异显著性的检验, 结果P<0.05, 说明各处理间出现了显著差异。

图 7 不同浓度Cd2+对普通小球藻H2O2含量的影响
3 结论
镉是水体污染主要重金属之一, 在通常情况下微量的镉对水体中的生物是有益的, 但是当含量超过一定量时就会对水体中的生物产生一定的危害。因此本次研究用不同浓度的镉来处理小球藻以研究其对小球藻的生长状况及细胞中蛋白质、多糖、叶绿素、MDA和一些氧化酶的影响。结果表明不同浓度的镉对小球藻生理有不同程度的影响, 同时小球藻对镉的胁迫也会产生相应的生理应答来减缓危害。
[1]Surosz W, Palinska K A. Ultrastructural changes induced by selected cadmium and copper concentrations in the cyanobacteria Phormidium: Interaction with salinity [J]. Journal of Plant Physiology, 2000, 157(6): 643—650
[2]Heng L Y, Jusoh K, Ling C H, et al. Toxicity of single and combinations of lead and cadmium to the cyanobacteria Anabaena flos-aquae [J]. Bulletin of Envionmental Contamination and Toxicology, 2004, 72(2): 373—379
[3]Ministry of Environmental Protection of the People’s Republic of China. 12thFive-year Plan for the Pollution Prevention of Major Watershed and Immediate Offshore Area [EB/OL]. Ministry of Environmental Protection of the People’s Republic of China. 2011-5-9
[4]Wang S, Xiao Y, Li Z, et al. Preliminary research on cultivated simulation of Chlorella under axenic culture [J]. Acta Hydrobiologica Sinica, 2016, 40(3): 580—588 [王舒, 肖艳, 李哲, 等. 小球藻纯种室内模拟培养初探. 水生生物学报, 2016, 40(3): 580—588]
[5]Bowler C, Allen A E, Badger J H, et al. The phaeodactylum genome reveals the evolutionary history of diatom genomes [J]. Nature, 2008, 456(7219): 239—244
[6]Yao W T, Cai D S, Lin J C, et al. Diatom assemblages distribution along the middle and lower reaches of Hejiang River in relation to heavy metal pollution [J]. Environmental Science & Technology, 2015, 38(3): 21—25 [姚文婷, 蔡德所, 林金城, 等. 贺江重金属污染对硅藻群落组成的影响. 环境科学与技术, 2015, 38(3): 21—25]
[7]Yang G Y. The physiological, biochemical responses and detoxification of Scenedesmus obliquus to heavy metals lead and chromium [D]. Thesis for Master of Science. Ji’nan University, Guangzhou. 2013 [杨国远. 斜生栅藻对重金属铅、铬胁迫的生理生化响应及其解毒效应.硕士研究生论文, 暨南大学, 广州. 2013]
[8]Fan C C. The biological adsorption effect research of rat tail algae on heavy metal lead, copper, zinc, cadmium in water [D]. Thesis for Master of Science. Zhejiang Ocean University, Zhoushan. 2013 [范彩彩. 鼠尾藻对水体重金属铅、铜、锌、镉的生物吸附效应研究. 硕士研究生论文, 浙江海洋学院, 舟山. 2013]
[9]Lü L Y. Study on bioremediation ability of several seaweeds on heavy metal contamination in seawater [D]. Thesis for Master of Science. Ocean University of China, Qingdao. 2013 [吕利云. 几种大型海藻对海水中重金属污染的生物修复能力研究. 硕士研究生论文, 中国海洋大学, 青岛. 2013]
[10]Ran X F, Liu R, Bai F, et al. The response on the growth and photosystem Ⅱ of Microcystis aeruginosa to cadmium, a heavy metal [J]. Acta Hydrobiologica Sinica, 2015, 39(3): 627—632 [冉小飞, 刘瑞, 白芳, 等. 微囊藻生长及光合系统Ⅱ对重金属镉的响应. 水生生物学报, 2015, 39(3): 627—632]
[11]Meng C X, Gao Z Q. A review and prospective of rehabilitation of heavy metals water pollution by microalgae [J]. Fisheries Science, 2009, 28(12): 795—797 [孟春晓,高政权. 微藻对重金属污染的生物修复研究现状与展望. 水产科学, 2009, 28(12): 795—797]
[12]Becker E W. Microalgae: Biotechnology and Microbiology [M]. Cambridge: Cambridge University Press. 1994, 56—62
[13]Wang X K. Plant Physiological and Biochemical Experimental Techniques [M]. Beijing: Higher Education Press, 2005, 134—136 [王学奎. 植物生理生化实验技术. 北京: 高等教育出版社, 2005, 134—136]
[14]Wang X Q, Qin S Y, Gao T H, et al. Basic Biochemical Experiments [M]. Beijing: Higher Education Press. 1999, 6 [王秀奇, 秦淑媛, 高天慧, 等. 基础生物化学实验. 北京: 高等教育出版社. 1999, 6]
[15]Li H S. Plant Physiological and Biochemical Principles and Techniques [M]. Beijing: Higher Education Press. 2000 [李合生. 植物生理生化原理和技术. 北京: 高等教育出版社. 2000]
[16]Beauchamp C, Fridovich I. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels [J]. Analytical Biochemistry, 1971, 44(1): 276—287
[17]Sergiev I, Alexieva V, Karanov E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants [J]. Comptroller Rend Academy Bulgarian Sciences, 1997, 51(3): 121—124
[18]Li S S. Cultivation of Chlorella and shudy on the conditions of lipid production [D]. Thesis for Master of Science. Zhejiang A&F University, Hangzhou. 2014 [李双双. 小球藻的培养及产油条件研究. 硕士研究生论文,浙江农林大学, 杭州. 2014]
[19]Calabrese E J. Hormesis: principles and applications for pharmacology and toxicology [J]. Amercian Joumal Pharmacology & Toxicology, 2008, 3(1): 59—71
[20]Wang D F, Li H Y, Wang L, et al. Effects of chitosan-RE3+-bentonite on growth of Chlorella vulgaris [J]. Journal of Rare Earth, 2010, 28(10): 149—153
[21]Geng H, Liu J F, Wang N. Effect of mercury and cadmium on the growth of chlorella vulgaris [J]. Journal of South-Central University for Nationalities (Natural Science Edition), 2014, 33(3): 41—43 [耿红, 刘剑锋, 王诺.重金属汞和镉对普通小球藻生长的影响. 中南民族大学学报: 自然学版, 2014, 33(3): 41—43]
[22]Zhang X Y. A study of the optimal cultivation condition of Cr enrichment by Dunaiella salinanal Chiorella pyrenoides [D]. Thesis for Master of Science. Ji’nan University, Guangzhou. 2004 [张学颖. 盐藻和小球藻富集铬(III)的优化培养条件研究.硕士研究生论文, 暨南大学, 广州. 2004]
[23]Yin W K, Cheng J F, Xiao W L, et al. Effect of Cadmium on Scenedesmus quadricauda [J]. Journal of Agro-Environment Science, 2015, 34(4): 633—638 [尹文珂, 程金凤, 肖婉露, 等. 四尾栅藻对重金属镉胁迫的响应. 农业环境科学学报, 2015, 34(4): 633—638]
[24]Xu Q J, Jin X C, Wang X M, et al. Effects of different concentration ammonium-N on Hydrilla verticillata antioxidant enzymes under Cd stress [J]. Chinese Journal of Applied Ecology, 2007, 18(2): 420—424 [许秋瑾, 金相灿, 王兴民, 等. 不同浓度铵态氮对镉胁迫轮叶黑藻生长及抗氧化酶系统的影响. 应用生态学报, 2007, 18(2): 420—424]
[25]Wang Z, Wang S G, Li H S, et al. Plant Physiology [M]. Beijing: China Agricultural Publishing House. 1999 [王忠, 王三根, 李合生, 等. 植物生理学. 北京: 中国农业出版社. 1999]
[26]Stobart A K, Griffiths W T, et al. The effect of Cd2+on the biosynthesis of chlorophyll in leaves of barley [J]. Physiological Plant, 1985, 63: 293—298
[27]Wang X H. Pollution Ecology Foundation [M]. Kunming: Yunnan University Press. 1990 [王校焕. 污染生态学基础. 昆明: 云南大学出版社. 1990]
[28]Rui C, Angela S, Richard C R, et al. The effect of cadmium on antioxidant responses and the susceptibility to oxidative stress in the hydrothermal vent mussel bathymodiolus azoricus [J]. Marine Biology, 2006, 148(4): 817—825
[29]Zhang X L, Shi G X, Xu N, et al. Effects of mercury and cadmium on some of physiological indicators of chara [J]. Journal of Nanjing Normal University (Natural Science Edition), 2002, 25(1): 38—43 [张小兰, 施国新, 徐楠, 等. Hg2+、Cd2+对轮藻部分生理生化指标的影响. 南京师大学报(自然科学版), 2002, 25(1): 38—43]
[30]Yang G Y. The physiological,biochemical responses and detoxification of Scenedesmus obliquus to heavy metals lead and chromium [D]. Thesis for Master of Science.Ji’nan University, Guangzhou. 2004 [杨国远. 斜生栅藻对重金属铅、铬胁迫的生理生化响应及其解毒效应.硕士学位论文, 暨南大学, 广州. 2004]
[31]Li F Y. Physiological and biochemical responses of different rice cultivars to cadmium stress in tropical region of China [J]. Chinese Journal Of Ecology, 2010, 29(4): 821—825 [李福燕. 热带地区不同水稻品种对土壤镉胁迫的生理生化响. 生态学杂志, 2010, 29(4): 821—825]
[32]Zhou X, Zhang T, Ren X M, et al. Research progress on technology of SOD extraction [J]. Cereal & Food Industry, 2015, 22(4): 57—62 [周轩, 张天, 任晓敏, 等. SOD提取工艺研究进展. 粮食与食品工业, 2015, 22(4): 57—62]
[33]Yu J, Tang X X, Zhang P Y, et al. Effects of CO2enrichment on growth, photosynthesis and activities of antioxidant enzymes of two marine micro-green-algae [J]. Acta Ecologica Sinica, 2005, 25(2): 197—202
[34]Qiu C E, Bi Y H, Hu Z Y. The effects of Zn2+stress on the growth, physiological characteristics and cell structure of Chlorococcum sp. [J]. Acta Hydrobiologica Sinica, 2007, 31(4): 503—508 [邱昌恩, 毕永红, 胡征宇. Zn2+胁迫对绿球藻生长、生理特性及细胞结构的影响.水生生物学报, 2007, 31(4): 503—508]
[35]Rijstenbil J W, Derksen J W M, Gerringa L J A, et al. Oxidative stress induced by copper: defense and damage in the marine planktonic diatom Ditylum brightwellii, grown in continuous cultures with high and low zinc levels [J]. Marine Biology, 1994, 119(4): 583—590
[36]Jiao T N, Zhu H. Physiological responses of Hydrilla verticillata to Cadmium and Cadmium Bioaccumulation [J]. Chinese Agricultural Science Bulletin, 2014, 30(5): 249—253 [焦铁男, 朱宏. 黑藻(Hydrilla verticillata)对重金属Cd2+的积累及生理响应. 中国农学通报, 2014, 30(5): 249—253]
[37]Li S W, Xue L G, Feng H Y, et al. Hydrogen peroxide signaling and its biological importance in plants [J]. Chinese Journal of Biochemistry and Molecular Biology, 2007, 23(10): 804—810 [李师翁, 薛林贵, 冯虎元, 等. 植物中的H2O2信号及功能. 中国生物化学与分子生物学报, 2007, 23(10): 804—810]
[38]Song X G, She X P. The generation and the role of hydrogen peroxide in plant [J]. Journal of Lianyungang Normal College, 2010, 27(4): 99—103 [宋喜贵, 佘小平. 植物体内过氧化氢的产生及其生理作用. 连云港师范高等专科学校学报, 2010, 27(4): 99—103]
THE PHYSICAL RESPONSES OF CHLORELLA TO CADMIUM STRESS UNDER DIFFERENT CONCENTRATIONS
XI Yu-Qin, REN Chun-Yan, ZHU Qiao-Qiao, KONG Wei-Bao and SUN Dui-Xiong
(College of Life Science, Northwest Normal University, Lanzhou 730070, China)
Cadmium is highly toxic to plant growth. It causes toxic effects on primary producers in water. The physiological and biochemical characteristics were investigated when Chlorella vulgaris was exposed to different concentrations of Cd2+. The results showed that the contents of protein, soluble sugar and chlorophyll in C. vulgaris cells increased under 5—40 μmol/L of Cadmium, and were slightly higher than those of the control group when the concentration reached 30 μmol/L; however, they all gradually decreased when the Cadmium concentration was higher than 30 μmol/L. The SOD activity and H2O2content increased firstly, and then decreased, which reached their peaks at 35 μmol/L. The content of MDA increased with the increase of Cd2+concentration. These results indicated that C. vulgaris can respond positively to the heavy metal cadmium stress to keep their normal growth and metabolism.
Chlorella vulgaris; Cadmium Stress; Osmotic; MDA; Oxidase
Q142
A
1000-3207(2017)05-1106-06
10.7541/2017.138
2016-11-15;
2017-04-18
甘肃省科技支撑计划社会发展类(1504FKCA025); 国家自然科学基金项目(11564037)资助 [Supported by the Gansu Science and Technology Support Plan Social Development (1504FKCA025); the National Natural Science Foundation of China (11564037)]
葸玉琴(1964—), 女, 甘肃永昌人; 副教授, 硕士; 主要研究方向为低等植物生理生化和环境污染生态学。E-mail: xiyuqin @nwnu.edu.cn
