重组旋毛虫plancitoxin-1-like活性位点突变体蛋白的核酸酶活性
2017-09-03廖成水王晓利田文静张梦珂张春杰李银聚吴庭才程相朝
廖成水,王晓利,田文静,张梦珂,张春杰,李银聚,吴庭才,程相朝,3
1 河南科技大学 动物科技学院 洛阳市活载体生物材料与动物疫病防控重点实验室,河南 洛阳 471023 2 河南科技大学 医学院,河南 洛阳 471023 3 洛阳职业技术学院,河南 洛阳 471099
重组旋毛虫plancitoxin-1-like活性位点突变体蛋白的核酸酶活性
廖成水1,王晓利2,田文静1,张梦珂1,张春杰1,李银聚1,吴庭才1,程相朝1,3
1 河南科技大学 动物科技学院 洛阳市活载体生物材料与动物疫病防控重点实验室,河南 洛阳 471023 2 河南科技大学 医学院,河南 洛阳 471023 3 洛阳职业技术学院,河南 洛阳 471099
廖成水, 王晓利, 田文静, 等. 重组旋毛虫plancitoxin-1-like活性位点突变体蛋白的核酸酶活性. 生物工程学报, 2017, 33(8):1315-1324.
Liao CS, Wang XL, Tian WJ, et al. Nuclease activity of the recombinant plancitoxin-1-like proteins with mutations in the active site from Trichinella spiralis. Chin J Biotech, 2017, 33(8): 1315-1324.
旋毛虫plancitoxin-1-like (Ts-Pt) 是旋毛虫125种DNaseⅡ家族蛋白中唯一具有典型DNaseⅡ活性区域HKD基序的核酸酶,且普遍认为,组氨酸位点是DNaseⅡ的活性氨基酸位点。为研究Ts-Pt活性位点突变体蛋白的核酸酶活性,利用重叠PCR方法获得Ts-Pt活性位点突变体片段,以pET-28a(+)为载体构建重组表达质粒并在大肠杆菌中诱导表达。重组Ts-Pt突变体蛋白经亲和层析纯化后进行SDS-PAGE分析。利用琼脂糖凝胶电泳法和核酸酶酶谱分析重组Ts-Pt突变体蛋白的核酸酶活性。成功构建含Ts-Pt突变体重组质粒的基因工程菌,SDS-PAGE和亲和层析纯化结果显示,重组Ts-Pt突变体蛋白呈包涵体表达。重组蛋白经复性后并没有表现出核酸酶活性,但核酸酶酶谱分析结果显示,包涵体表达的重组Ts-Pt突变体蛋白表现出降解DNA的能力。同时,N端和C端活性位点H及HCK和DHSK突变并不影响Ts-Pt的核酸酶活性,研究结果为进一步研究庞大的DNaseⅡ家族蛋白在旋毛虫发育和感染方面的作用提供一定的参考。
旋毛虫,plancitoxin-1-like,活性位点,突变体,核酸酶
脱氧核糖核酸酶 (DNase) 是一类能够催化水解DNA骨架上磷酸二酯键的酶类的统称,参与机体细胞凋亡和免疫保护等多种生命活动。根据底物特异性、化学机制和生物学功能的不同,主要将DNase分为DNaseⅠ和DNaseⅡ两大类。根据现有研究资料显示,绝大多数物种存在一种DNaseⅠ或多种DNaseⅡ同源物。2011年公布的旋毛虫基因组序列分析显示,旋毛虫可能存在一个多达 125种 DNaseⅡ蛋白家族。旋毛虫拥有如此庞大的 DNaseⅡ蛋白,这在其他物种中是极为罕见的。DNaseⅡ家族成员氨基酸序列的N端和C端都具有典型的高度保守的活性区域,即HKD基序 (H-x-K-x(4)-D)[1]。但笔者前期利用在线生物信息学软件 Conserved Domain Database (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) 对旋毛虫125种DNase Ⅱ家族蛋白进行保守区域分析,结果显示,只有plancitoxin-1蛋白 (1 095 bp, GenBank AccessionNo. XM_003370715.1) 在 C端具有 1个典型DNaseⅡ活性位点HKD基序。对该基因进行克隆和测序发现,扩增获得的基因序列比GenBank预测的短 210 bp,并且相对应的氨基酸序列中在N端和C端都具有HKD基序,重新命名为plancitoxin-1-like (Ts-Pt)[2]。
虽然有报道称旋毛虫肌幼虫时期排泄分泌物中存在核酸酶活性,但旋毛虫125种DNase Ⅱ蛋白中到底哪个蛋白发挥核酸酶活性仍然不清楚,笔者前期利用核酸酶酶谱证明了经大肠杆菌表达的重组Ts-Pt蛋白表现出一定的核酸酶活性[2]。氨基酸 N端和C端的两个HKD基序是DNaseⅡ的活性位点区域,并且组氨酸可能是DNaseⅡ发挥核酸酶活性的关键位点,但目前仍没有足够的研究证据表明组氨酸就是HKD基序的关键位点。因此,本研究通过重叠PCR方法对 Ts-Pt的活性位点 HKD基序进行突变构建Ts-Pt突变体表达载体,经大肠杆菌表达、纯化后利用核酸酶酶谱分析各种重组突变体蛋白的核酸酶活性,试图探讨HKD基序在DNaseⅡ发挥核酸酶活性的作用。
1 材料与方法
1.1 材料
1.1.1 质粒与菌株
重组质粒 pET28a-Ts-Pt和 Rosetta (DE3)/(pET28a-Ts-Pt) 由笔者构建[2],大肠杆菌Rosetta(DE3) 菌株均由洛阳市活载体生物材料与动物疫病防控重点实验室提供。
1.1.2 主要试剂
胰蛋白胨和酵母浸出物均购自英国 Oxoid公司;Taq DNA聚合酶、DNA marker、T4 DNA连接酶、BamHⅠ和 Hind Ⅲ均购自宝生物工程(大连) 有限公司;质粒小量提取试剂盒和DNA凝胶回收试剂盒均购自美国Axygen公司;BCA蛋白浓度测定试剂盒、氨苄青霉素、卡那霉素、IPTG和咪唑均购自北京鼎国昌盛生物技术有限公司;预染蛋白marker购自美国Genview公司;鲑鱼精DNA和Triton X-100均购自美国Sigma-Aldrich公司;其他化学试剂均为国产分析纯。
1.2 方法
1.2.1 引物设计
参照旋毛虫 plancitoxin-1-like基因 (Ts-Pt)的碱基序列 (GenBank Accession No. KF984291)设计构建DNaseⅡ活性位点HKD基序突变体重组表达载体的引物 (表 1)。引物由生工生物工程 (上海) 股份有限公司合成。所构建的各种突变体分别命名为 H→A、H→L、HCK→LAQ、DHSK→VLLQ和HCK/DHSK→LAQ/VLLQ。
1.2.2 Ts-Pt突变体表达载体的构建
以引物Ts-Pt上游和突变引物上游、突变引物下游和引物 Ts-Pt下游作为引物,利用 Taq DNA聚合酶从质粒 pET28a-Ts-Pt模板上进行PCR扩增Ts-Pt突变体的上游和下游片段,然后以引物Ts-Pt上游和下游通过重叠PCR的方法得到 Ts-Pt突变片段。将 Ts-Pt突变片段亚克隆到pMD18-T载体中并转化入大肠杆菌 DH5α。经PCR、酶切鉴定和测序正确后将Ts-Pt突变片段连接至pET-28a(+)构建Ts-Pt突变体重组表达质粒。
1.2.3 Ts-Pt突变体的原核表达与蛋白纯化
将Ts-Pt突变体重组表达质粒转化入大肠杆菌Rosetta (DE3)株,经PCR、酶切鉴定和测序正确后将重组菌接种于含卡那霉素的 LB液体培养基中,37 ℃、160 r/min振荡培养至OD600约为0.6时,加入终浓度为1 mmol/L的IPTG诱导表达,培养5 h后收集菌体,经超声破碎仪破碎后收集沉淀。用包涵体结合缓冲液溶解包涵体,经 0.22 μm一次性滤器过滤后利用美国GE Healthcare公司的AKTA Purifier 100系统纯化His标签的表达蛋白。用透析袋梯度透析除去咪唑,采用Bradford法检测重组蛋白的浓度。
1.2.4 SDS-PAGE
收集各种Ts-Pt突变体表达纯化的重组蛋白和野生型Ts-Pt重组蛋白用于SDS-PAGE分析。取适量蛋白加入SDS-PAGE蛋白上样缓冲液,煮沸10 min、冰浴5 min,8 000 r/min离心5 min后取上清。将重组蛋白在12%分离胶和5%浓缩胶的聚丙烯酰胺凝胶进行电泳。电泳结束后将凝胶分别于考马斯亮蓝染色液和考马斯亮蓝脱色液中进行染色和脱色。
1.2.5 重组蛋白的复性
根据相关参考文献报道的包涵体复性方法进行重组蛋白的复性[3],即选择 0.2 mmol/L DTT、0.3 mmol/L GSSG和3 mmol/L GSH作为复性液母液,将含有包涵体的复性液 (8 mol/L尿素) 装入截留分子量为8 kDa的透析袋中,在4 ℃条件下,依次用含6、5、4、3.5、3、2.5、2、1.5、1、0.5和0 mol/L尿素的复性液进行透析 (袋内袋外溶液体积比为1∶50),每4 h换液1次,同时充分搅拌。
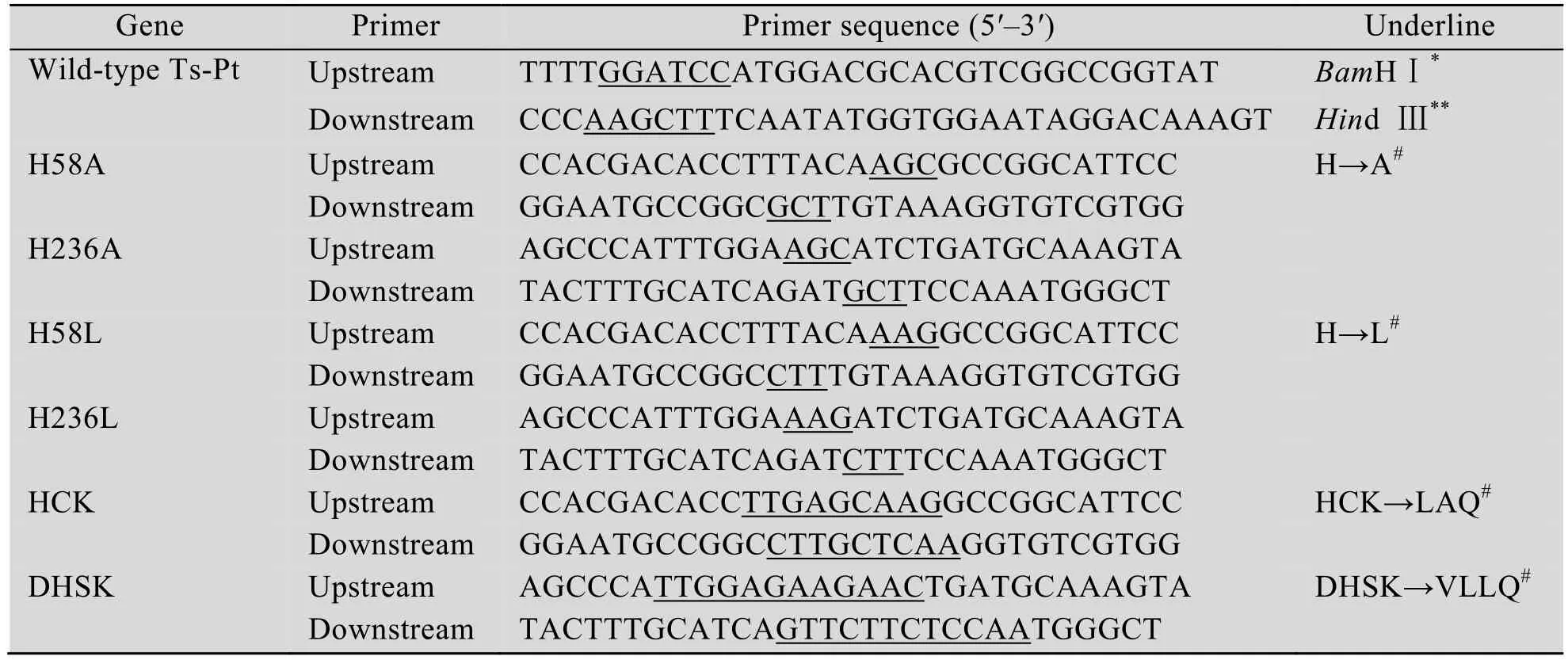
表1 本研究中所用的PCR扩增引物序列Table 1 The primer sequences for PCR amplification in this study
1.2.6 琼脂糖凝胶电泳法测定复性重组 Ts-Pt突变体蛋白的核酸酶活性
配制 50 mmol/L 10×Acetate缓冲液 (pH 5.0),按下列参数配制20 μL反应体系:10×缓冲液 2 μL,鲑鱼精 DNA 0.2 μg,重组 Ts-Pt突变体蛋白或野生型Ts-Pt重组蛋白10 μg,补充ddH2O至总体积20 μL。混匀后置于水浴锅37 ℃反应 1 h后将酶切产物进行 1%琼脂糖凝胶电泳,凝胶成像系统下观察DNA降解程度并拍照。
1.2.7 重组Ts-Pt突变体蛋白的核酸酶酶谱分析
核酸酶的酶谱分析与SDS-PAGE相似,差别在于酶谱在胶内添加底物DNA,且样品不进行加热处理及上样缓冲液中不含 β-巯基乙醇。取 50 μg重组Ts-Pt突变体蛋白或野生型Ts-Pt重组蛋白,加入不含 β-巯基乙醇的上样缓冲液,37 ℃水浴锅孵育15 min后上样,按10 mA/浓缩胶和20 mA/分离胶于4 ℃冰箱中进行横流电泳。电泳结束后取出分离胶置于平皿中,用预冷 ddH2O洗涤后于预冷复性液轻摇孵育30 min,换液4次,预冷ddH2O洗涤后转移至37 ℃预热的反应液中,反应2 d后于含有EB染料中轻摇染色10 min,凝胶成像系统下观察胶内底物DNA降解情况并拍照。
2 结果与分析
2.1 Ts-Pt突变体重组表达载体的鉴定
利用重叠 PCR方法将旋毛虫 Ts-Pt氨基酸序列第58和236位的H突变为A或L,第58/59和60位的HCK突变为LAQ,第235、236、237和238位的DHSK突变为VLLQ,各突变体PCR产物经琼脂糖凝胶电泳检测,在750-1 000 bp之间出现特异性条带,大小均与野生型Ts-Pt相等 (图1A)。目的片段亚克隆至pMD18T构建重组质粒,经酶切鉴定目的条带与预期结果相符(图 1B)。测序正确后回收酶切目的片段与pET-28a(+)相连构建 Ts-Pt突变体重组表达质粒。测序结果显示,各种Ts-Pt突变体重组表达质粒的目的基因片段已成功突变。
2.2 重组Ts-Pt突变体蛋白的鉴定
SDS-PAGE结果显示,各种Ts-Pt突变体重组表达质粒的大肠杆菌Rosetta(DE3) 经IPTG诱导后在分子质量34-43 kDa之间均得到一条重组蛋白,大小与野生型重组Ts-Pt蛋白一致 (图2A)。为了研究重组蛋白的核酸酶活性,利用His-Trap经AKTA Purifier 100纯化系统纯化表达的重组蛋白 (图2B)。但亲和层析纯化结果显示,原核表达的重组Ts-Pt蛋白呈包涵体表达 (图3)。
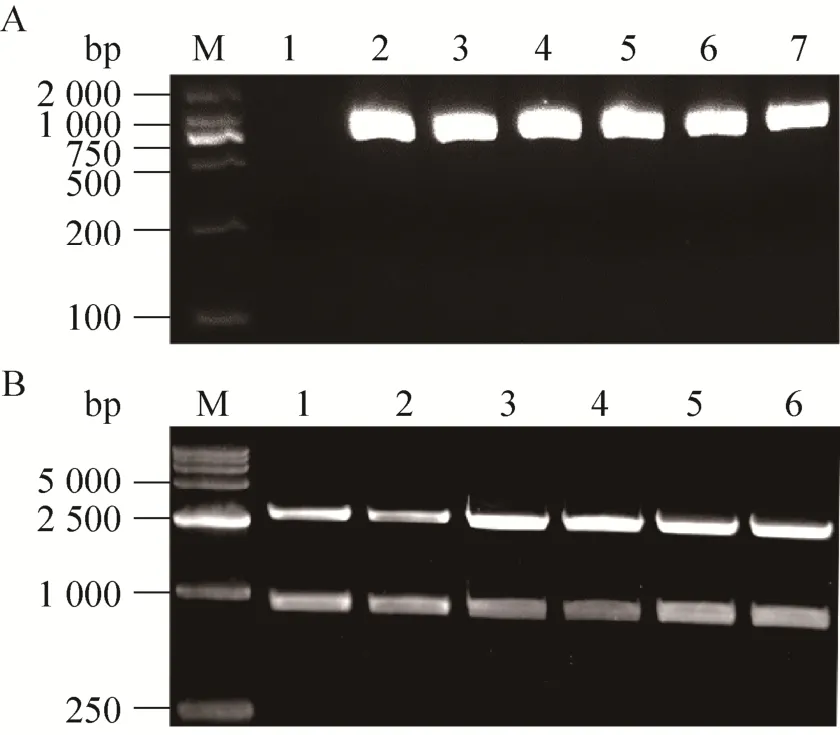
图1 Ts-Pt突变体的PCR扩增和酶切鉴定Fig. 1 PCR amplification and enzyme digestion of Ts-Pt with active site mutant. (A) PCR amplification of Ts-Pt gene with active site mutant. M: DL2 000 DNA ladder; 1: ddH2O; 2: wild-type Ts-Pt; 3: H→A; 4: H→L;5: HCK→LAQ; 6: DHSK→VLLQ; 7: HCK/DHSK→LAQ/VLLQ. (B) Enzyme digestion of pMD18T-Ts-Pt with active site mutant. M: DL15 000 DNA ladder; 1:wild-type Ts-Pt; 2: H→A; 3: H→L; 4: HCK→LAQ; 5:DHSK→VLLQ; 6: HCK/DHSK→ LAQ/VLLQ.
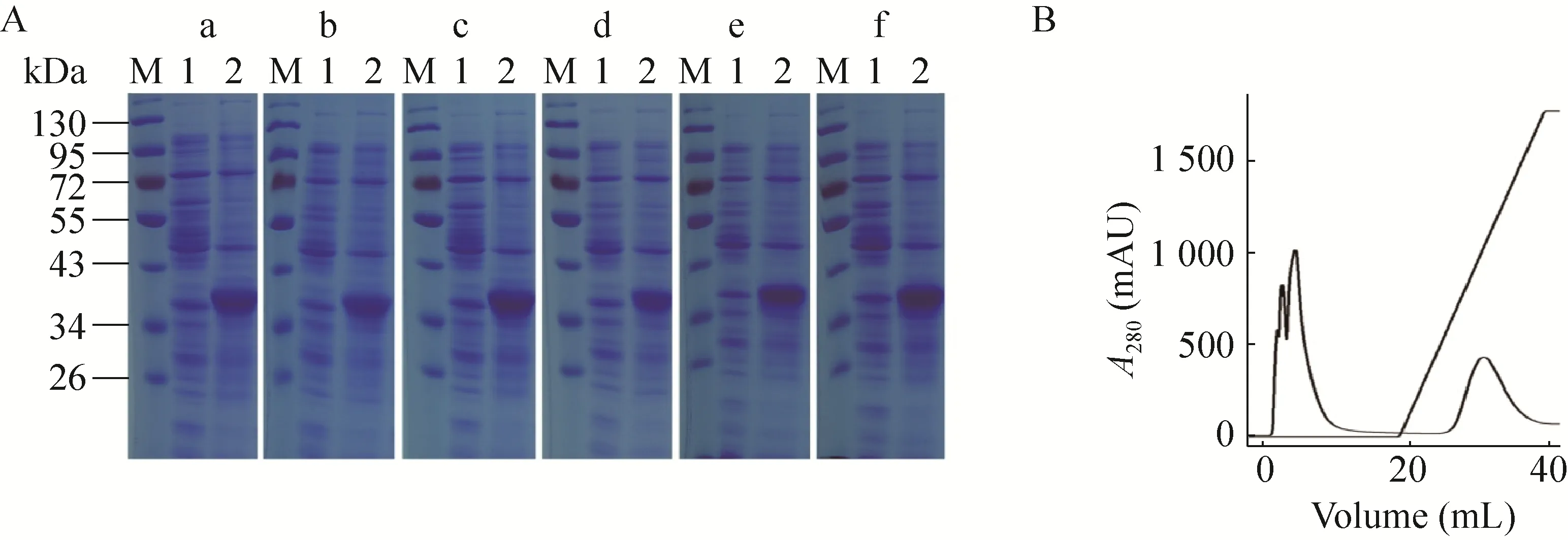
图2 重组Ts-Pt突变体蛋白的纯化及SDS-PAGE分析Fig. 2 Purification and SDS-PAGE analysis of the recombinant proteins. (A) SDS-PAGE analysis of the recombinant proteins.a: wild-type Ts-Pt; b: H→A; c: H→L; 5: HCK→LAQ; d: DHSK→VLLQ; e: HCK/DHSK→LAQ/VLLQ. M: prestained protein marker; 1: E. coli lysate without IPTG; 2: E. coli lysate with IPTG. (B) Purification of the recombinant proteins.
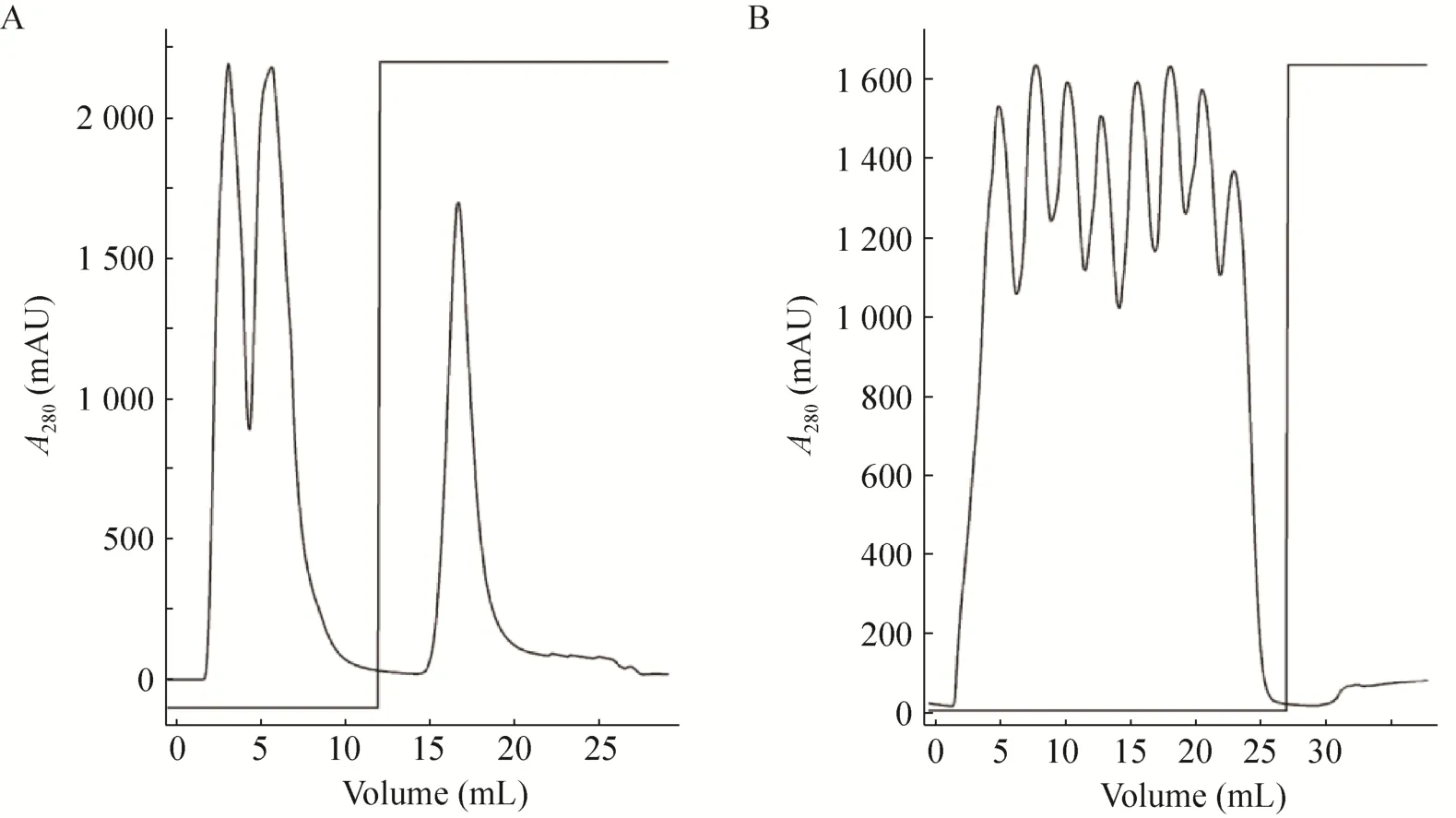
图3 重组Ts-Pt蛋白的亲和层析纯化Fig. 3 Purification of the recombinant proteins by affinity chromatography. (A) Purification of inclusion body. (B)Purification of soluble protein.
2.3 琼脂糖凝胶电泳法分析复性重组 Ts-Pt突变体蛋白的核酸酶活性
收集复性的重组Ts-Pt突变体蛋白,采用琼脂糖凝胶电泳法分析复性重组Ts-Pt突变体蛋白的核酸酶活性,结果如图 3所示,与 DNaseⅡ标准品相比,各种复性的重组Ts-Pt突变体蛋白并未表现出降解鲑鱼精DNA的能力 (图4)。
2.4 核酸酶酶谱分析重组 Ts-Pt突变体蛋白的核酸酶活性
核酸酶酶谱分析结果显示,在紫外灯照射下聚丙烯酰胺凝胶之间有一条黑色条带,大小与SDS-PAGE一致 (图5)。DNaseⅡ标准品活性较强,降解区域呈片状,而BSA蛋白阴性对照则未见降解 DNA的现象。这说明表达的各种Ts-Pt突变体蛋白经SDS-PAGE的胶内复性后能够降解鲑鱼精DNA。
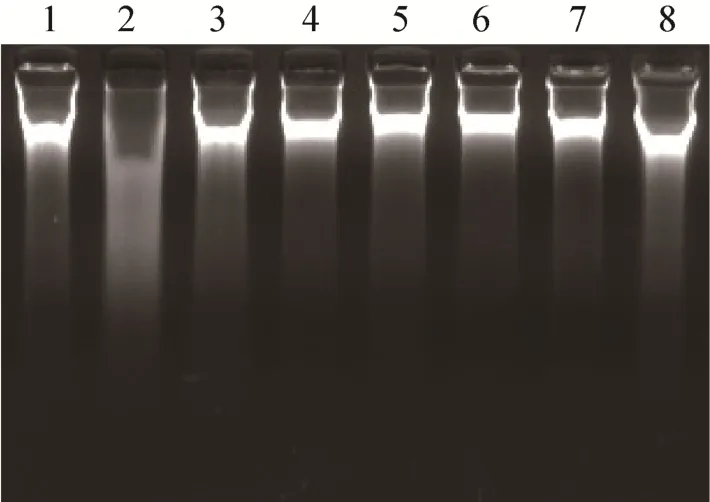
图4 琼脂糖凝胶电泳法分析重组Ts-Pt突变体蛋白的核酸酶活性Fig. 4 Detection of the nuclease activity of the recombinant proteins by agarose gel electrophoresis. 1:ddH2O; 2: DNaseⅡ; 3: wild-type Ts-Pt; 4: H→A; 5:H→L; 6: HCK→LAQ; 7: DHSK→VLLQ; 8:HCK/DHSK→LAQ/VLLQ.
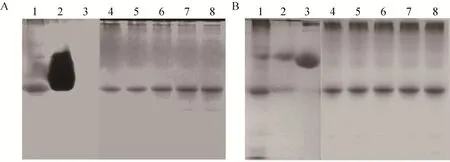
图5 核酸酶酶谱分析重组Ts-Pt突变体蛋白的核酸酶活性Fig. 5 Detection of the nuclease activity of the recombinant proteins by nuclease-zymography. (A)Nuclease-zymography analysis of the recombinant proteins, after staining with ethidium bromide and exposure to UV light. (B) The same gel was stained with Coomassie Brilliant blue after zymography. 1: wild-type Ts-Pt; 2: DNase Ⅱ;3: BSA. 4: H→A; 5: H→L; 6: HCK→LAQ; 7: DHSK→VLLQ; 8: HCK/DHSK→LAQ/VLLQ.
3 讨论
DNaseⅡ于20世纪40年代首先发现于哺乳动物中,随后陆续报道了其生物学特性。但直至1998年首次克隆到人类DNaseⅡ[4],对其研究才深入到分子水平。之后的研究发现,DNaseⅡ广泛存在于低等的原核生物和高等的哺乳动物中[5],包括牛、马、猪、小鼠、大鼠、鸡、斑马鱼、非洲蟾蜍、果蝇、海星、河豚、类鼻疽杆菌、盘基网柄菌、鸡痘病毒、金丝雀痘病毒、冈比亚按蚊、广杆属线虫和秀丽隐杆线虫等20多种生物。从目前已报道的序列上分析,大部分哺乳动物具有DNaseⅡα和DNaseⅡβ两种同源物,而秀丽隐杆线虫是已知存在最多 DNaseⅡ同源物的物种[6],多达10余种,而其他绝大多数物种只有一种或少数几种同源物。
旋毛虫是一种寄生于骨骼肌细胞的重要人兽共患胞内寄生虫,约55个国家1.1亿人感染,死亡率在0.2%左右,主要分布于中国、阿根廷和一些东欧国家等[7]。旋毛虫生活史包括新生幼虫、肌幼虫和成虫。Mitreva等通过构建旋毛虫成虫、新生幼虫和肌幼虫 3个不同发育时期的抑制性差减杂交cDNA文库,筛选到28个期特异性DNaseⅡ家族基因,其中成虫期家族12个、新生幼虫期家族15个、肌幼虫期家族1个。虽然 3个时期 DNaseⅡ间的基因相似性非常低,仅介于32%-36%,但家族内成员的基因相似性则高达 94%-99%。但是根据 2011年公布的旋毛虫基因组数据显示,旋毛虫拥有其他寄生性线虫和非寄生性线虫所无法比拟的 DNaseⅡ基因家族,达到惊人的125个DNaseⅡ蛋白家族,并且一半以上的基因被认为编码排泄/分泌产物[8]。虽然旋毛虫具有如此庞大的DNaseⅡ蛋白家族,但旋毛虫为何表现出完全不一样的DNaseⅡ数量,DNaseⅡ在旋毛虫发育和感染中的作用以及125个DNaseⅡ蛋白家族是否真具有核酸酶活性,目前关于这些相关研究报道较少。
水解酶活性的检测研究的方法包括琼脂糖凝胶电泳、分光光度计和胶内酶谱,琼脂糖凝胶电泳法是最为直接的,但琼脂糖凝胶电泳和分光光度计法一样存在非特异性问题。目前特异性检测酶活性的研究方法主要包括胶内酶谱、原位酶谱和体内酶谱3种酶谱方法[9-10]。胶内酶谱已在各种生物体蛋白质、多肽、低聚糖、多糖、脂质以及核酸酶的检测、分析和鉴定中得到广泛应用[11]。胶内酶谱中的核酸酶酶谱是指在SDS-PAGE过程中在分离胶中加入DNA作为底物,核酸酶降解的区域EB染色在紫外灯下呈黑色条带,可直接观察活性核酸酶的分子量大小[12]。相对于琼脂扩散法的定性和分光光度计的定量,胶内酶谱具有定性和定量的优势[13],胶内酶谱可通过分子量大小直接特异观察核酸酶活性[10]。胶内酶谱已成为研究核酸酶活性的一种常用方法[14],通过胶内核酸酶酶谱的方法已证实了许多动物、植物、细菌、真菌和寄生虫组织及其分泌排泄物的天然核酸酶。
对于真核生物表达或原核生物可溶性表达的核酸酶蛋白可直接分析核酸酶活性。但一些蛋白结构复杂,外源基因在大肠杆菌中呈高水平表达时,由于新生肽链的聚集速率远远大于蛋白正确折叠的速率,就会形成无活性的蛋白聚集体即包涵体[15]。分子和蛋白技术的不断发展使得包涵体经合适的复性过程获得生物活性蛋白成为可能。在本研究中,在重组Ts-Pt以及各种突变体蛋白的表达过程中,通过改变各种表达参数 (培养基、表达载体、宿主菌、IPTG浓度、诱导温度和诱导时间等) 仍然无法获得可溶性蛋白。同时通过梯度稀释、透析和柱上复性等复性方法进行包涵体复性,利用琼脂糖凝胶电泳法对复性后得到的少量蛋白进行核酸酶活性分析,并未观察到降解DNA的能力 (部分数据未提供)。利用核酸酶酶谱分析呈包涵体表达的重组Ts-Pt及突变体蛋白,结果证明了表达蛋白具有核酸酶活性。但是需要指出的是,完全阐明包涵体表达的蛋白经胶内酶谱表现出活性的确切理论对于目前现有的技术仍是个挑战。从包涵体的体外溶解变性和复性原理可能可以解释这种胶内变性复性的机制[16-17],即在不含 β-巯基乙醇的上样缓冲液中,高密度聚集的包涵体于 SDS作用下溶解形成中间体,同时在整个SDS-PAGE过程中也处于一个变性环境足以有效防止聚集,胶内的 SDS可以很容易地利用含有Triton X-100或奶粉的复性液除去[14,18],从而促使中间体重新折叠成蛋白的天然构象,起到蛋白复性的作用[19-20]。
据报道,旋毛虫排泄/分泌产物中存在活性核酸内切酶[21-22]。有学者认为,旋毛虫 p43、SS1和AAK85403三个蛋白的氨基酸与人类DNase Ⅱ表现出一定的相似性,但3个蛋白的氨基酸序列在N端和C端均不存在DNaseⅡ经典的活性区域HKD基序[23],并且未对p43、SS1和AAK85403三个蛋白进行核酸酶活性分析。氨基酸序列N端和 C端的两个 HKD基序 (H-x-K-x(4)-D) 是DNaseⅡ经典的活性位点区域,DNaseⅡ属于单体核酸酶,其催化作用的活性中心由两个 HKD基序形式假二聚体构成[24]。前期研究发现,Ts-Pt在N端和C端保守区域具有2个HKD基序,且原核表达的重组Ts-Pt蛋白在核酸酶酶谱中表现出一定的核酸酶活性[2]。因此,本研究通过重叠PCR方法对Ts-Pt的活性位点N端和C端的H、HCK和DHSK进行突变,利用核酸酶酶谱分析各种重组突变体蛋白的核酸酶活性。令人惊奇的是,各种重组突变体蛋白均表现出降解DNA的能力。因此,可以提出一个科学命题:DNaseⅡ是否是依赖于HKD基序发挥活性?虽然本研究未能解释这种现象的原因,但值得深入探究。虽然吴秀萍等对旋毛虫新生幼虫期脱氧核糖核酸酶T31D4的核酸酶活性及活性位点进行了鉴定[25],但该研究并未证实组氨酸位点是 DNaseⅡ的真正活性氨基酸位点。
REFERENCES:
[1] Schäfer P, Cymerman IA, Bujnicki JM, et al.Human lysosomal DNase IIα contains two requisite PLD-signature (HxK) motifs: evidence for a pseudodimeric structure of the active enzyme species. Protein Sci, 2007, 16(1): 82–91.
[2] Liao CS, Liu MY, Bai X, et al. Characterisation of a plancitoxin-1-like DNase Ⅱ gene in Trichinella spiralis. PLoS Negl Trop Dis, 2014,8(8): e3097.
[3] Zhang SH, Jia WZ, Luo XN, et al. Renaturation,purification and antigenicity identification of recombinant protein of Cysticercus cellulosae expressed in Escherichia coli. Chin J Biotech,2008, 24(8): 1490–1495 (in Chinese).张少华, 贾万忠, 骆学农, 等. 猪囊尾蚴 CE18重组蛋白的复性纯化及抗原性鉴定. 生物工程学报, 2008, 24(8): 1490–1495.
[4] Yasuda T, Takeshita H, Iida R, et al. Molecular cloning of the cDNA encoding human deoxyribonuclease Ⅱ. J Biol Chem, 1998, 273(5):2610–2616.
[5] Evans CJ, Aguilera RJ. DNase II: genes, enzymes and function. Gene, 2003, 322: 1–15.
[6] Lai HJ, Lo SJ, Kage-Nakadai E, et al. The roles and acting mechanism of Caenorhabditis elegans DNase Ⅱ genes in apoptotic DNA degradation and development. PLoS ONE, 2009, 4(10): e7348.
[7] Gottstein B, Pozio E, Nöckler K. Epidemiology,diagnosis, treatment, and control of trichinellosis.Clin Microbiol Rev, 2009, 22(1): 127–145.
[8] Mitreva M, Jasmer DP, Zarlenga DS, et al. The draft genome of the parasitic nematode Trichinella spiralis. Nat Genet, 2011, 43(3): 228–235.
[9] Vandooren J, Geurts N, Martens E, et al.Zymography methods for visualizing hydrolytic enzymes. Nat Methods, 2013, 10(3): 211–220.
[10] Napirei M, Ricken A, Eulitz D, et al. Expression pattern of the deoxyribonuclease 1 gene: lessons from the Dnase1 knockout mouse. Biochem J,2004, 380(3): 929–937.
[11] Fischer H, Scherz J, Szabo S, et al. DNase 2 is the main DNA-degrading enzyme of the stratum corneum. PLoS ONE, 2011, 6(3): e17581.
[12] Rosenthal AL, Lacks SA. Nuclease detection in SDS-polyacrylamide gel electrophoresis. Anal Biochem, 1977, 80(1): 76–90.
[13] Budič M, Kidrič M, Meglič V, et al. A quantitative technique for determining proteases and their substrate specificities and pH optima in crude enzyme extracts. Anal Biochem, 2009, 388(1):56–62.
[14] Wilkesman J, Kurz L. Protease analysis by zymography: a review on techniques and patents.Recent Pat Biotechnol, 2009, 3(3): 175–184.
[15] Fang M, Huang HL. Advances in in vitro refolding of inclusion body proteins. Chin J Biotech, 2001,17(6): 608–612 (in Chinese).方敏, 黄华樑. 包涵体蛋白体外复性的研究进展. 生物工程学报, 2001, 17(6): 608–612.
[16] De Bernardez Clark E. Refolding of recombinant proteins. Curr Opin Biotechnol, 1998, 9(2):157–163.
[17] Eiberle MK, Jungbauer A. Technical refolding of proteins: do we have freedom to operate?Biotechnol J, 2010, 5(6): 547–559.
[18] Ludwig S, Mannherz HG, Schmitt S, et al. Murine serum deoxyribonuclease 1 (Dnase1) activity partly originates from the liver. Int J Biochem Cell Biol, 2009, 41(5): 1079–1093.
[19] Boone DL, Tsang BK. Non-isotopic technique for the identification of endonucleases involved in apoptosis. Biotechniques, 1997, 22(4): 648–649.
[20] Thimon V, Belghazi M, Labas V, et al. One- and two-dimensional SDS-PAGE zymography with quenched fluorogenic substrates providesidentification of biological fluid proteases by direct mass spectrometry. Anal Biochem, 2008,375(2): 382–384.
[21] Mak CH, Ko RC. Characterization of endonuclease activity from excretory/secretory products of a parasitic nematode, Trichinella spiralis. Eur J Biochem, 1999, 260(2): 477–481.
[22] Liu RD, Qi X, Sun GG, et al. Proteomic analysis of Trichinella spiralis adult worm excretorysecretory proteins recognized by early infection sera. Vet Parasitol, 2016, 231: 43–46.
[23] MacLea KS, Krieser RJ, Eastman A. A family history of deoxyribonuclease Ⅱ: surprises from Trichinella spiralis and Burkholderia pseudomallei.Gene, 2003, 305(1): 1–12.
[24] Cheng YC, Hsueh CC, Lu SC, et al. Identification of three crucial histidine residues (His115, His132and His297) in porcine deoxyribonuclease Ⅱ.Biochem J, 2006, 398(2): 177–185.
[25] Wu XP, Yu L, Wang XL, et al. Identification of enzymatic activity and activity sites of DNase Ⅱprotein from Trichinella spiralis. J Jilin Agric Univ, 2010, 32(2): 209–213 (in Chinese).吴秀萍, 于录, 王学林, 等. 旋毛虫脱氧核糖核酸酶-Ⅱ酶活及酶活性位点的鉴定. 吉林农业大学学报, 2010, 32(2): 209–213.
(本文责编 陈宏宇)
Nuclease activity of the recombinant plancitoxin-1-like proteins with mutations in the active site from Trichinella spiralis
Chengshui Liao1, Xiaoli Wang2, Wenjing Tian1, Mengke Zhang1, Chunjie Zhang1,Yinju Li1, Tingcai Wu1, and Xiangchao Cheng1,3
1 Luoyang Key Laboratory of Live Carrier Biomaterial and Animal Disease Prevention and Control, College of Animal Science and Technology, Henan University of Science and Technology, Luoyang 471023, Henan, China 2 Medical College, Henan University of Science and Technology, Luoyang 471023, Henan, China 3 Luoyang Vocational and Technical College, Luoyang 471099, Henan, China
Although there are 125 predicted DNase Ⅱ-like family genes in the Trichinella spiralis genome,plancitoxin-1-like (Ts-Pt) contains the HKD motif, a typical conserved region of DNase Ⅱ, in N- and C-terminal. It is generally believed that histidine is the active site in DNase Ⅱ. To study the nuclease activity of recombinant Ts-Pt with mutations in the active site from T. spiralis, different fragments of the mutated Ts-Pt genes were cloned using overlap PCR technique and inserted into the expressing vector pET-28a(+), and transformed into Escherichia coli Rosseta (DE3). The fusion proteins were purified by Ni-NTA affinity chromatography and SDS-PAGE. Nuclease activity of the recombinant proteins was detected by agarose gel electrophoresis and nuclease-zymography. The recombinant plasmids harboring the mutated Ts-Pt genes were constructed and expressed as inclusive body in a prokaryotic expression system. After renaturation in vitro, the recombinant proteins had no nuclease activity according to agarose gel electrophoresis. However,the expressed proteins as inclusive body displayed the ability to degrade DNA after renaturation in gel. And the nuclease activity was not affected after subjected to mutation of active site in N- and C-termini of Ts-Pt. These results provide the basis to study the relationship between DNase Ⅱ-like protein family and infection of T. spiralis.
Trichinella spiralis, plancitoxin-1-like, active site, mutation, nuclease
January 10, 2017; Accepted: May 8, 2017
s:Chengshui Liao. E-mail: liaochengshui33@163.com Xiangchao Cheng. E-mail: chengxch@126.com
Supported by: National Natural Science Foundation of China (No. 31572489), Henan Science and Technology Key Project (No.152102110078), Key Project of Henan Province for Scientific Research Higher Education of China Colleges and Universities (No.17A230009), PhD Start-up Fund of Henan University of Science and Technology (No. 13480071).
国家自然科学基金(No. 31572489),河南省科技攻关项目 (No. 152102110078),河南省高等学校重点科研项目计划 (No.17A230009),河南科技大学博士启动基金项目 (No. 13480071) 资助。
