喀斯特地区土壤厚度降低和水分减少对两种草本植物混种后光合的影响
2017-08-11赵雅洁宋海燕梁千慧刘锦春
赵雅洁,李 周,宋海燕,张 静,梁千慧,刘锦春
(三峡库区生态环境教育部重点实验室 重庆市三峡库区植物生态与资源重点实验室 西南大学生命科学学院,重庆 400715)
喀斯特地区土壤厚度降低和水分减少对两种草本植物混种后光合的影响
赵雅洁,李 周,宋海燕,张 静,梁千慧,刘锦春
(三峡库区生态环境教育部重点实验室 重庆市三峡库区植物生态与资源重点实验室 西南大学生命科学学院,重庆 400715)
土壤“厚薄不均”和“岩溶干旱”是影响喀斯特地区植物生存、生长和繁殖的关键生态因子,深根植物和浅根植物由于根系生态位的互补可能会缓解资源减少对光合作用的抑制作用。以盆栽深根植物苇状羊茅(Festucaarundinacea)和浅根植物多年生黑麦草(Loliumperenne)为对象,研究土壤厚度降低、水分减少及土壤厚度和水分双重资源减少对两种草本植物混种下其光合作用的影响。结果表明,1)在高资源水平下,混种后苇状羊茅的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和水分利用效率(WUE)与单种间无显著差异(P>0.05);而黑麦草的Pn、Gs和Tr低于单种,WUE显著高于单种(P<0.05)。2)随着资源的减少,不管是单种还是混种,苇状羊茅和黑麦草的Pn、Gs和Tr大体上均呈降低的趋势,黑麦草的WUE呈上升趋势,且混种后降低或升高的幅度明显低于单种;而苇状羊茅的WUE由单种时升高变为混种时降低。3)在低资源水平下,混种后苇状羊茅的Pn、Gs和Tr高于单种,WUE低于单种;黑麦草的Pn、Gs和Tr与单种无显著差异,WUE低于单种。研究表明,在干旱和土壤贫瘠日趋严重的喀斯特地区,通过深浅不同根系的植物混种,可能在一定程度上缓解水分和土壤资源缺乏对植物光合作用的抑制作用。
岩溶干旱;土壤异质性;光合作用;水分利用效率;生态位互补;深根植物;浅根植物
在喀斯特地区,特殊的地质环境背景和长期的岩溶作用等,使得该地区土层浅薄,土壤厚薄变化大且分布不连续[1]。如在喀斯特洼地、盆地和谷地中,土层较厚,且分布较连续;而在山丘坡地上,土壤呈“岩浪土”或缺失,分布极不连续[2]。喀斯特地区的另一个生境特点是由于岩石漏渗性强,土壤保水、蓄水能力差而造成的“岩溶干旱”胁迫[1,3-4]。大多数植物在此环境下生长缓慢,造成该地区植被覆盖率低,植物群落单调,多以灌草丛为主,在小于10 cm的浅土层中一般只能生长草本植物[5-6]。近十几年来,随着全球降水格局的变化,中国西南地区出现高温少雨的趋势,连续极端的天气日益突出,这将加剧喀斯特地区“岩溶干旱”发生的频率和强度[7-9]。
光合作用决定植物能量吸收和有机物积累的数量,与植物生长、发育和繁殖密切相关,是其它生理过程和生命活动的基础[10-11]。植物的光合作用对外界环境变化高度敏感,它对不同的生境环境会表现不同的适应特性和适应机制[12-13]。喀斯特土壤水分不足,直接影响植物的光合作用。如喀斯特的水分胁迫引起火棘(Pyracanthafortuneana)和小果蔷薇(Rosacymosa)等植物的叶片水势下降,造成气孔开度减小,CO2进入叶片受阻,导致植物由于光合底物(CO2)不足引起光合速率下降[14];而荩草(Arthraxonhispidus)和画眉草(Eragrostispilosa)等的叶肉细胞受到严重损伤,叶绿素含量和光合酶活性降低,导致植物光合速率降低[15]。另一方面,植物的光合作用与土壤资源密切相关。土壤资源匮乏造成植物生长所需的养分不足,同时也进一步加重干旱胁迫,因而会抑制气孔开放,也导致植物光合作用降低[16-17]。目前,关于喀斯特生态系统中干旱生境对植物光合作用的影响研究较多,但土壤厚度变化对该地区植物光合作用影响研究很少,而土壤厚度变化和岩溶干旱相互作用对该地区植物的影响研究更为鲜见。
植物地上部分生长所需的水分和矿物质,主要由根系供应。当土壤水分和养分充足时,植物生长旺盛;反之,土壤中水分和养分缺乏,地上部分生长受到抑制[18]。也就是说,在植物地上和地下部分协同实现植物的生长过程中,植物地下根系吸收并传至地上部分的水分和养分的多少影响着植物的光合作用。在喀斯特地区,浅根系植物主要利用来自浅层土壤的水分和养分,而深根系植物常常利用不同深度的水分和养分资源,且能依靠储存在岩石裂缝或裂隙中的水分来抵御干旱胁迫[19-21]。同时,深根植物在干旱的土壤环境中还可以通过根系提水作用,将深层土壤水分吸收后释放到干旱的上层土壤,供邻体植物吸收利用[22-23]。因此,当浅根系植物和深根系植物共存时,由于二者的根系在空间上垂直分层,有效形成生态位的互补关系,避免强烈的种间竞争,提高土壤水分和养分的利用率[24-25]。有研究发现,深根植物和浅根植物混种后能缓解干旱对深根植物的胁迫,提高植物的光合速率和气孔导度等,有利于合成和积累更多的光合产物,增强植物的逆境适应性[26-27]。故当深根植物和浅根植物混种后,由于根系生态位互补能为地上部分提供更多养分和水分,可能会促进二者的光合作用。
苇状羊茅(Festucaarundinacea)为禾本科多年生草本,根系主要分布在0-40 cm土壤层,但仍有16.1%的根系可以分布在大于40 cm的土层深处,为深根植物;而多年生黑麦草(Loliumperenne)根系主要分布在0-20 cm的土层中,为浅根植物[28](表1)。二者均为世界范围内重要的牧草和观赏性草坪草,耐旱耐瘠薄,适应性强,在喀斯特地区作为石漠化草地治理的草种[29-30]。我们选择这两个物种,通过盆栽模拟试验,探究土壤和水分资源减少对两者混种后光合作用的影响,以验证以下假说:土壤和水分资源减少均会抑制植物的光合作用,但深根植物和浅根植物混种后,由于生态位的互补,资源减少对光合作用的抑制作用可能得到缓解(即混种后两物种的光合作用降低幅度均低于单种,以至于在低资源水平下混种后两物种的光合作用均高于单种)。

表1 苇状羊茅和黑麦草的根系分布比例Table 1 Root spatial distribution of F. arundinacea and L. perenne in different soil depth
注:数据来源于成文竞等[23]。
Note: The dates are based on Cheng et al[23].
1 材料与方法
1.1 试验材料
供试材料为苇状羊茅和多年生黑麦草。种子购自重庆中一种业有限公司。供试土壤取自重庆中梁山海石公园的黄色石灰土。土壤的基本理化性状:pH 7.4,有机质0.34%,全氮0.28g·kg-1,全磷0.39 g·kg-1,全钾23.7 g·kg-1,测得田间持水量39.8%。
1.2 试验设计
由于喀斯特地区土层浅薄,土壤厚度普遍小于8 cm,有些地方甚至只有4.28 cm,20 cm厚度的土壤占的比例通常不到30%[31-32],且用喀斯特地区的土壤做预备试验时发现,15 cm土层是黑麦草和苇状羊茅根系生长最旺盛的区域。鉴于此,本研究以15 cm土壤厚度作为高土壤资源,而5 cm土壤厚度作为低土壤资源,这两种土壤资源水平以自制的两种不同深度的长方体栽培容器实现:底面积均为100 cm2,高度分别为15 cm[干土为1 500 g,作为对照土壤厚度(DCK)]和5 cm[干土500 g,作为浅土组(DS)]。2015年1月14日,在西南大学生态园实验基地大棚(透明顶棚,四周开放)下进行播种,园地海拔高度为249 m,年平均温度18 ℃,年平均相对湿度为80%,全年日照1 288.1 h[33-34]。至2015年4月4日,待幼苗长到20 cm左右,选取生长基本一致的两种幼苗移栽到容器中。种植方式分为单种和混种,每盆两株。
待幼苗适应生长一段时间后,于2015年4月14日进行水分处理。水分处理分为两个水平,分别为正常灌溉[每3天灌溉水120 mL,作为对照水分(WCK)]和干旱组[每3天灌溉水36 mL(WD]。水分处理是根据重庆地区1981-2011年这30年间4月-6月降水量计算出每100 cm2面积每日平均降水量为40 mL,高度为4 mm。以日平均降水量40 mL为对照组,干旱组(WD)在对照的水平上减少70%(12 mL)。水分处理历时69 d,每3 d浇水一次。
试验共设置了土壤资源[对照土壤厚度(DCK)和浅(DS)]、水分[对照水分(WCK)和干旱(WD)]和种植方式(单种和混种)3个处理,每个处理两个水平按随机区组设计,每个物种共计8个处理,每个处理5个重复,因单种中有两物种,共计60盆(图1)。在分析数据时,当分析土壤资源减少的影响时,水分处理固定为对照水分(WCK),即分析组为DCKWCK和DSWCK;当分析水分资源减少的影响时,土壤处理固定为对照土壤厚度(DCK),即分析组为DCKWCK和DCKWD;当分析土壤和水分资源同时减少的影响时,分析组是对照组(DCKWCK)和浅土组+干旱组(DSWD)。
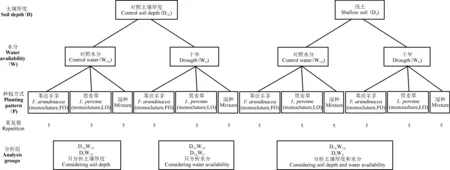
图1 试验设计Fig. 1 Experiment design
1.3 指标测定
在2015年6月15日至18日,于晴朗天气09:30-12:00,每株选取相同部位的成熟叶片,每个处理选取5个植株,使用Li-6400便携式光合分析仪(Li-COR,USA)测定气孔交换参数。测定前,先用饱和光对植物进行30 min的光诱导。研究表明,黑麦草和苇状羊茅的光饱和点大约在800~1 200 μmol·(m2·s)-1,在固定光照为1 000 μmol·(m2·s)-1时,两种植物均表现较强的光合能力[33-34]。因此,测定时控制光合有效辐射(PAR)为1 000 μmol·(m2·s)-1,叶温25 ℃,流速为500 μmol·s-1。测定结束后,将标定放入叶室的植物叶片区域立即放入冰袋带回实验室,使用数字化扫描仪(STD1600Epson USA)获得叶图像,并运用WinRhizo(Version 410B)根系分析软件(Regent Instrument Inc.,Canada)对叶面积进行定量分析,然后通过换算得出各处理的光合参数。仪器自动测定净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr),并计算叶片水分利用效率(WUE)=Pn/Tr。
1.4 统计分析
用Microsoft Office Excel 2013软件整理数据,用SPSS 22.0软件分析数据。利用Two-way ANOVA分析土壤厚度和种植方式、水分和种植方式、土壤厚度+水分和种植方式对植物光合生理参数的影响;利用Independent-samples T-test分析相同土壤厚度和水分条件下同一物种在单种和混种处理下各参数的差异。用Origin 8.6软件作图。
2 结果与分析
2.1 净光合速率
在高资源水平下(对照土壤厚度+水分),混种后苇状羊茅的净光合速率与单种无显著差异(P>0.05)。不管单种还是混种,其净光合速率均随着所有资源(包括土壤厚度、水分、土壤厚度+水分)的减少而降低,但混种后的下降幅度明显低于单种,故在低资源水平下混种后苇状羊茅的净光合速率显著或稍高于单种。在单种情况下,苇状羊茅的净光合速率在浅土、干旱和浅土+干旱3种处理中分别比相应对照降低了57.68%、49.23%和60.16%;混种情况下,其净光合速率在浅土、干旱和浅土+干旱3种处理中分别比相应对照降低了23.74%、21.87%和15.43%。
与单种相比,混种后黑麦草的净光合速率在高资源水平下显著降低(降低幅度为28.08%)(P<0.05)。单种情况下,随着资源的减少黑麦草的净光合速率也随之降低,在浅土、干旱和浅土+干旱3种处理中分别比相应对照降低了44.74%、42.58%和58.76%;在混种情况下,净光合速率随着土壤(浅土)和双重资源(浅土+干旱)的减少而降低,但随着水分(干旱)的减少而升高,升高的幅度为45.22%。
双因素方差分析表明苇状羊茅的净光合速率在土壤厚度和种植方式、水分和种植方式、土壤厚度+水分和种植方式的交互作用下都无显著差异(P>0.05),黑麦草的净光合速率在土壤厚度和种植方式、水分和种植方式、土壤厚度+水分和种植方式的交互作用下有显著差异(表2、3)。
注:D,土壤厚度;W,水分;**,P<0.01;***,P<0.001;*,P<0.05;+,0.05
Note: D, soil depth; W, water; ***,P<0.001; **,P<0.01; *,P<0.05; +, 0.05 2.2 气孔导度 苇状羊茅的气孔导度变化趋势与净光合速率基本一致。在高资源水平下,混种对苇状羊茅的气孔导度无显著影响(P>0.05)(图3)。无论是单种还是混种,气孔导度均随着资源的减少而降低,但混种后降低的幅度明显低于单种。在高资源水平下,混种后黑麦草的气孔导度也呈降低趋势。随着资源的减少,不管是单种还是混种,黑麦草的气孔导度均随之呈降低的趋势,但混种后降低的幅度要低于单种,以至于在低资源水平下二者没有显著差异。 双因素方差分析表明,苇状羊茅和黑麦草的气孔导度在土壤厚度和种植方式、水分和种植方式、土壤厚度+水分和种植方式的交互作用下都无显著差异(P>0.05)(图3和表2、3)。 2.3 蒸腾速率 与净光合速率和气孔导度一样,在高资源水平下,混种后苇状羊茅的蒸腾速率没有显著变化(P>0.05)(图4)。随着资源的减少,单种时苇状羊茅的蒸腾速率显著降低,但在混种时,却随着水分及土壤+水分双重资源的减少而增加(增加幅度分别为45.45%和35.95%);当然,混种后蒸腾速率随着土壤资源的减少而降低,但降低幅度远小于单种。因此,在3种低资源水平下,混种后苇状羊茅的蒸腾速率均显著(P<0.05)或稍高于单种。 在高资源水平下,混种后黑麦草的蒸腾速率显著降低(降低幅度为59.83%),但随着资源的降低,单种时蒸腾速率随之显著下降,而混种时无显著变化(P>0.05)。因此,在3种低资源水平下,混种后黑麦草的蒸腾速率与单种无显著差异(P>0.05)。 双因素方差分析表明苇状羊茅(土壤厚度和种植方式除外)和黑麦草的蒸腾速率在土壤厚度和种植方式、水分和种植方式、土壤厚度+水分和种植方式的交互作用下都有显著或极显著差异(表2、3)。 2.4 水分利用效率 在高资源水平下,混种后苇状羊茅的水分利用效率与单种相比仍然无显著差异(P>0.05)(图5)。随着资源的降低,单种时苇状羊茅的水分利用效率均显著升高(P<0.05),但混种时升高不明显(土壤厚度)或者呈下降趋势(水分、水分+土壤厚度)。总体上,在低资源水平时,混种后苇状羊茅的水分利用效率显著低于单种(P<0.05)。 表3 黑麦草光合生理的双因素方差分析Table 3 Summary of the two-way ANOVA test for the effects of soil depth and planting pattern, water and planting pattern, soil depth + water and planting pattern on the photosynthetic physiology of L. perenne 图3 土壤厚度、水分以及土壤厚度+水分对单、混种条件下苇状羊茅和黑麦草气孔导度的影响Fig. 3 Effect of soil depth, water, and soil depth plus water on stomatal conductance of F. arundinacea and L. perenne when grown in monoculture and mixture 在高资源水平下,混种后黑麦草的水分利用效率得到促进(增加幅度为78.82%)。随着资源的降低,单种和混种后其水分利用效率均呈升高的趋势,但混种后上升幅度总体低于单种。在低资源水平(除了干旱)时,混种后黑麦草的水分利用效率高于单种。 双因素方差分析表明,苇状羊茅的水分利用效率在水分和种植方式、土壤厚度+水分和种植方式的交互作用下有显著差异,黑麦草的水分利用效率在土壤厚度和种植方式、水分和种植方式、土壤厚度+水分和种植方式的交互作用下都无显著差异(P>0.05)(表2、表3)。 光合作用是植物生长和产量的生理基础,对环境中的资源变化有着最直接的反应,能够很好地反映植物对生境中土壤资源的适应状况[35-37]。在高资源水平下,混种后深根植物苇状羊茅的净光合速率与单种无显著差异,这表明苇状羊茅并没有受到混种的促进作用,而混种后浅根植物黑麦草的净光合速率显著低于单种,这表明黑麦草反而受到了混种的抑制作用。本研究发现苇状羊茅的叶面积和株高显著大于黑麦草,具有光竞争优势。研究认为,成功竞争者的叶片一定要处于有利于光截留的位置,而植物叶片在邻体植株之上的排列是获得有效光最显著的手段,相比叶面积小的植物而言,具有较大叶面积的植物常常具有光竞争优势[38]。植物高度与光竞争能力之间存在显著的正相关,矮小型植物受到高生长型植物的严重抑制,高生长型植物比矮小型植株对光的竞争能力强[38-39]。故当黑麦草与具有光竞争优势的苇状羊茅混种后,受到邻株苇状羊茅遮阴的抑制,导致黑麦草光合作用降低,进而使得混种后黑麦草的光合作用低于单种。有研究也证明,具有光竞争优势的植物可使得邻体植物的气孔导度降低,光合速率受到抑制[40-41]。 图4 土壤厚度、水分以及土壤厚度+水分对单、混种条件下苇状羊茅和黑麦草蒸腾速率的影响Fig. 4 Effect of soil depth, water and soil depth plus water on transpiration rate of F. arundinacea and L. perenne when grown in monoculture and mixture 与假设一致,随着资源的减少(土壤厚度、水分及土壤厚度+水分),不管是单种还是混种,苇状羊茅和黑麦草的净光合速率、气孔导度和蒸腾速率均呈降低的趋势,但混种后降低的幅度明显低于单种,说明深根植物和浅根植物混种后,由于生态位的互补,资源减少对光合作用的抑制作用确实得到了缓解。在低资源水平下,光照作为竞争因子之一的重要性降低,地下部分对土壤水分和养分资源竞争激烈[42-43]。由于生态位互补和深根植物根系提水作用逐渐充分发挥其作用,混种后苇状羊茅和黑麦草叶片感受的根源信号相对单种较弱,两者可以通过提高气孔导度,来提高光合作用,因而两者的光合作用受到资源减少的抑制作用得以缓解。另一方面也说明,在低资源水平下苇状羊茅和黑麦草都能一定程度上通过提高光合作用来提高自身能量吸收和物质积累,进而提高竞争能力。 植物以单位水汽蒸腾量来同化最大量碳的原则来控制气孔行为,通常用水分利用效率(WUE)表达这一行为[44]。它反映了植物的水分利用策略,是植物抗旱策略的重要组成部分[45-46]。本研究中,在高资源水平下,混种后苇状羊茅的水分利用效率与单种无显著差异,而混种后黑麦草的水分利用效率显著高于单种。这表明混种对苇状羊茅的水分利用效率无影响,却提高了黑麦草的水分利用效率。这可能是由于混种后黑麦草受到苇状羊茅遮阴的影响,使得净光合速率降低的幅度小于蒸腾速率,造成混种后黑麦草水分利用效率的提高。刘贤赵和康绍忠[47]也发现,夏季遮阴有利于植物水分利用效率的提高。 干旱环境下,保存水分是植物生长、生存和繁殖的本质策略[48]。除了通过发展高密度的根系、提前落叶等来获取或减少水分的散失外,植物还能够通过提高水分利用效率、在有限的水资源下最大化的固定碳来适应干旱环境[21,45-46]。而喀斯特地区因其特殊的地质背景,即使充分降水后,土壤保持的田间持水量一般仅可供植物7~14 d的蒸腾[3,14]。这就要求该地区的植物不仅耐旱,而且应具有较高的水分利用效率,从而更大限度地利用稀缺的喀斯特土壤资源[49]。随着资源的减少,单种情况下苇状羊茅和黑麦草的水分利用效率整体上均呈上升趋势,且苇状羊茅上升的整体幅度显著高于黑麦草,并且在低资源水平下,苇状羊茅的水分利用效率高于黑麦草。总体来看,苇状羊茅要比黑麦草更适合生长在喀斯特地区。水分利用效率作为反映植物耐旱性的有效指标,在相同干旱条件下水分效率高的植物的抗旱能力强[48]。从本研究中也能得出苇状羊茅比黑麦草更抗旱。杨有俊等[50]也发现,苇状羊茅的抗旱性优于黑麦草。而苇状羊茅更耐旱的原因除了与自身遗传因素有关外,还可能是由于苇状羊茅比黑麦草的深层根系更多更长,能吸收更深层土壤水分和养分[51-52]。但在混种情况下,黑麦草的水分利用效率仍然呈上升趋势,且上升幅度总体上低于单种,而苇状羊茅甚至呈降低趋势。研究认为,在较湿润的环境中,较低的水分利用效率有利于植物获得较高的生产力,以提高自身的竞争力[48]。这表明混种后的苇状羊茅和黑麦草均以牺牲水分利用效率为代价而保持较高的净光合速率以提高自身的竞争力。 图5 土壤厚度、水分以及土壤厚度+水分对单、混种条件下苇状羊茅和黑麦草水分利用效率的影响Fig. 5 Effect of soil depth, water and soil depth plus water on water use efficiency of F. arundinacea and L. perenne when grown in monoculture and mixture 由于生态位互补和深根植物根系泵水作用,资源减少对两者光合作用的抑制作用均得到缓解,但以牺牲水分利用效率为代价以提高自身的竞争力。苇状羊茅因其面对干旱胁迫时具有较高的水分利用效率和耐旱性,比黑麦草更适宜生长在喀斯特地区,而其深根性可能是决定其具有较高耐旱性的主要原因之一。 全球气候变化背景下,在干旱和土壤贫瘠日趋严重的喀斯特地区,通过深浅不同根系的植物混种,可能在一定程度上缓解水分和土壤等资源缺乏对植物光合作用的抑制作用。然而本研究只利用了两个草本物种进行了光合试验,具有一定的局限性。期望以后有更多物种和更进一步的试验加以验证。 References: [1] 张军以,戴明宏,王腊春,苏维词,曹立国.西南喀斯特石漠化治理植物选择与生态适应性.地球与环境,2015,43(3):269-278. Zhang J Y,Dai M H,Wang L C,Su W C,Cao L G.Plant selection and their ecological adaptation for rocky desertification control in Karst region in the southwest of China.Earth and Environment,2015,43(3):269-278.(in Chinese) [2] 杨明德.论喀斯特环境的脆弱性.云南地理环境研究,1990,2(1):21-29. Yang M D.On the fragility of karst environment.Yunnan Geographic Environment Research,1990,2(1):21-29.(in Chinese) [3] 郭柯,刘长成,董鸣.我国西南喀斯特植物生态适应性与石漠化治理.植物生态学报,2011,35(10):991-999. Guo K,Liu C C,Dong M.Ecological adaptation of plants and control of rocky-desertification on Karst region of Southwest China.Chinese Journal of Plant Ecology,2011,35(10):991-999.(in Chinese) [4] 许留兴,熊康宁,张锦华,池永宽,谌妍,刘成名.西南喀斯特地区草地生态系统面临的问题及对策.草业科学,2015,32(5):828-836. Xu L X,Xiong K N,Zhang J H,Chi Y K,Chen Y,Liu C M.The problems and resolutions of grassland ecosystem in Karst of southwest China.Pratacultural Science,2015,32(5):828-836.(in Chinese) [5] 蔡秋,陈梅琳.贵州喀斯特山区环境特征与生态系统的恢复和重建.农业系统科学与综合研究,2001,17(1):49-53. Cai Q,Chen M L.The features of environment and ecosystem restoration and rehabilitation in Guizhou karst mountain.System Sciences and Comprehensive Studies in Agriculture,2001,17(1):49-53.(in Chinese) [6] 高帅,袁希平,甘淑,张晓伦.滇东南岩溶区土壤厚度与植被生长的关系研究.价值工程,2013(29):178-179. Gao S,Yuan X P,Gan S,Zhang X L.Study of the relationship between soil thickness and vegetation growth in the karst region in southeast Yunnan.Value Engineering,2013(29):178-179.(in Chinese) [7] IPCC.Climate Change 2014:Synthesis Report.Contribution of Working Groups Ⅰ,Ⅱ and Ⅲ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change.Core Writing Team,Pachauri R K,Meyer L A (eds.).Geneva:IPCC,2014:39-73. [8] Ma Z F,Liu J,Zhang S Q,Chen W X,Yang S Q.Observed climate changes in southwest China during 1961-2010.Advances in Climate Change Research,2013,4(1):30-40. [9] Yang J,Gong D,Wang W,Hu M,Mao R.Extreme drought event of 2009/2010 over southwestern China.Meteorology and Atmospheric Physics,2011,115(3-4):173-184. [10] 李庆康,马克平.植物群落演替过程中植物生理生态学特性及其主要环境因子的变化.植物生态学报,2002,26(Z1):9-19. Li Q K,Ma K P.Advances in plant succession ecophysiology.Acta Phytoecologica Sinica,2002,26(Z1):9-19.(in Chinese) [11] 张华,康雅茸,徐春华.兰州银滩黄河湿地4种植物的光合特性.草业科学,2016,33(4):622-634. Zhang H,Kang Y R,Xu C H.Photosynthetic characteristics of 4 wild plants in Yintan wetland in Lanzhou section of Yellow River.Pratacultural Science.2016,33(4):622-634.(in Chinese) [12] Rubio F C,Camacho F G,Sevilla J M,Chisti Y,Grima E M.A mechanistic model of photosynthesis in microalgae.Biotechnology and Bioengineering,2003,81(4):459-473. [13] Midgley G F,Aranibar J N,Mantlana K B,Macko S.Photosynthetic and gas exchange characteristics of dominant woody plants on a moisture gradient in anAfricansavanna.Global Change Biology,2004,10(3):309-317. [14] 刘长成,刘玉国,郭柯.四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性.植物生态学报,2011,35(10):1070-1082. Liu C C,Liu Y G,Guo K.Ecophysiological adaptations to drought stress of seedlings of four plant species with different growth forms in Karst habitats.Chinese Journal of Plant Ecology,2011,35(10):1070-1082.(in Chinese) [15] 刘成名,熊康宁,苏孝良,许留兴,张锦华,董颖萍.干旱胁迫对石漠化地区3种乡土草种光合作用的影响.四川农业大学学报,2014,32(4):382-387. Liu C M,Xiong K N,Su X L,Xu L X,Zhang J H,Dong Y P.Effects of drought stress on photosynthesis of three native grasses in rocky desertification area.Journal of Sichuan Agricultural University,2014,32(4):382-387.(in Chinese) [16] Ronchi C P,DaMatta F M,Batista K D,Moraes G A B K,Loureiro M E,Ducatti C.Growth and photosynthetic down-regulation inCoffeaarabicain response to restricted root volume.Functional Plant Biology,2006,33(11):1013-1023. [17] 王林,冯锦霞,万贤崇.土层厚度对刺槐旱季水分状况和生长的影响.植物生态学报,2013(3):248-255. Wang L,Feng J X,Wan X C.Effects of soil thickness on dry-season water relations and growth inRobiniapseudoacacia.Chinese Journal of Plant Ecology,2013,37(3):248-255.(in Chinese) [18] 潘瑞炽,董愚得.植物生理学.第二版(下册).北京:高等教育出版社,1984:65-69. [19] Querejeta J I,Estrada-Medina H,Allen M F,Jiménez-Osornio J J.Water source partitioning among trees growing on shallow karst soils in a seasonally dry tropical climate.Oecologia,2007,152:26-36. [20] Querejeta J I,Estrada-Medina H,Allen M F,Jiménez-Osornio J J,Ruenes R.Utilization of bedrock water byBrosimumalicastrumtrees growing on shallow soil atop limestone in a dry tropical climate.Plant and Soil,2006,287(1):187-197. [21] Schwinning S.The ecohydrology of roots in rocks.Ecohydrology,2010,3(2):238-245. [22] Horton J L,Hart S C.Hydraulic lift:A potentially important ecosystem process.Trends in Ecology and Evolution,1998,13(6):232-235. [23] Hirota I,Sakuratani T,Sato T,Higuchi H,Nawata E.A split-root apparatus for examining the effects of hydraulic lift by trees on the water status of neighbouring crops.Agroforestry Systems,2004,60(2):181-187. [24] Jose S,Williams R,Zamora D.Belowground ecological interactions in mixed-species forest plantations.Forest Ecology and Management,2006,233(2-3):231-239. [25] Wanvestraut R H,Jose S,Nair P R,Brecke B J.Competition for water in a pecan (CaryaillinoensisK.Koch)-cotton (GossypiumhirsutumL.) alley cropping system in the southern United States.Agroforestry Systems,2004,60(2):167-179. [26] 吴克顺.荒漠植物霸王根系提水研究.兰州:兰州大学硕士学位论文,2010. Wu K S.Study of hydraulic lift inZygophyllumxanthoxylumof eremophytes.Master Thesis.Lanzhou:Lanzhou University,2010.(in Chinese) [27] 赵纪东.白刺根-土界面水分再分配及其对干旱胁迫的响应.兰州:兰州大学硕士学位论文,2007. Zhao J D.Hydraulic redistribution ofNitrariasphaerocarpabetween root and soil.Master Thesis.Lanzhou:Lanzhou University,2007.(in Chinese) [28] 成文竞,崔建宇,闵凡华,胡林.三种草坪草的根系分布特征及其对土壤养分的影响.草业学报,2009,18(1):179-183. Cheng W J,Cui J Y,Min F H,Hu L.Root distribution characteristics of three turfgrasses and their impact on soil nutrient content.Acta Prataculturae Sinica,2009,18(1):179-183.(in Chinese) [29] Wei Q Z,Gao S R,Jiao L,Wu Y L.Research on model construction and technique support for integrated rehabilitation of rock desertification in Chaoying small watershed of Bijie test area of Guizhou Province.Meteorological and Environmental Research,2014,77(8):41-47. [30] 李馨,熊康宁,龚进宏,陈永毕.人工草地在喀斯特石漠化治理中的作用及其研究现状.草业学报,2011,20(6):279-286. Li X,Xiong K N,Gong J H,Chen Y B.Advances in research on the function of artificial grassland in karst rock desertification control.Acta Prataculturae Sinica,2011,20(6):279-286.(in Chinese) [31] 周运超,王世杰,卢红梅.喀斯特石漠化过程中土壤的空间分布.地球与环境,2010,38(1):1-7. Zhou Y C,Wang S J,Lu H M.Spatial distribution of soils during the process of karst rocky desertification.Earth and Environment,2010,38(1):1-7.(in Chinese) [32] 严冬春,文安邦,鲍玉海,张信宝.黔中高原岩溶丘陵坡地土壤中的137Cs分布.地球与环境,2008,36(4):342-347. Yan D C,Wen A B,Bao Y H,Zhang X B.The distribution of137Cs in hilly upland soil on the Qianzhong Karst Plateau.Earth and Environment,2008,36(4):342-347.(in Chinese) [33] 刘文丹.两种土壤基质中木本植物凋落物分解特性比较.重庆:西南大学硕士学位论文,2014. Liu W D.Comparison of above and belowground organs litter decomposition in two soil substrates.Master Thesis.Chongqing:Southwest University,2014.(in Chinese) [34] 刘锦春.重庆石灰岩地区柏木幼苗对水分胁迫的生理生态适应性研究.重庆:西南大学博士学位论文,2008. Liu J C.Adaptability of physiological ecology ofCupressusfunebrisEndl.seedling under water stress in limestone area in Chongqing.PhD Thesis.Chongqing:Southwest University,2008.(in Chinese) [35] 杨志民.不同光照强度对冷季型草坪草越夏的影响.北京:北京林业大学博士学位论文,2006. Yang Z M.Effects of different light intensity on summer hardiness of cool season turf grasses.PhD Thesis.Beijing:Beijing Forestry University,2006.(in Chinese) [36] 庞瑜.植物生长调节剂对两种冷季型草坪草生长及生理特性的影响研究.南京:南京农业大学硕士学位论文,2009. Pang Y.Research about effects of plant growth regulators on two kinds of cool-season turf-grass growth and physiological characteristics.Master Thesis.Nanjing:Nanjing Agricultural University,2009.(in Chinese) [37] 刘海燕,李吉跃,赵燕,黄看看.干旱胁迫对5个种源沙柳(Salixpsammophila)气体交换及水分利用效率的影响.干旱区研究,2007,24(6):815-820. Liu H Y,Li J Y,Zhao Y,Huang K K.Influence of drought stress on gas exchange and water use efficiency ofSalixpsammophilagrowing in five places.Arid Zone Research,2007,24(6):815-820.(in Chinese) [38] 李博.植物竞争:作物与杂草相互作用的实验研究.北京:高等教育出版社,2001:10-12. Li B.Plant Competition:The Experimental Research of Crop-Weed Interactions.Beijing:Higher Education Press,2001:10-12.(in Chinese) [39] Altieri M A,Liebman M.Weed management in agroecosystems:Ecological approaches.Florida:CRC Press,1988:89-119. [40] Dong T F,Li J Y,Zhang Y B,Korpelainen H,Niinemets Ü,Li C Y.Partial shading of lateral branches affects growth,and foliage nitrogen- and water-use efficiencies in the coniferCunninghamialanceolatagrowing in a warm monsoon climate.Tree Physiology,2015,35(6). [41] 苏本营,宋艳霞,陈圣宾,杨文钰.大豆幼苗对套作玉米遮荫环境的光合生理生态响应.生态学报,2015,35(10):3298-3308. Su B Y,Song Y X,Chen S B,Yang W Y.Photosynthetic responses of soybean (Glycinemax) seedlings to shading caused by maize in an intercropping system.Acta Ecologica Sinica,2015,35(10):3298-3308.(in Chinese) [43] Seabloom E W,Harpole W S,Reichman O J,Tilman D.Invasion,competitive dominance,and resource use by exotic and native California grassland species.Proceedings of the National Academy of Sciences,2003,100(23):13384-13389. [44] 米娜,温学发,蔡福,王阳,张玉书.季节性干旱对千烟洲人工林水分利用效率的影响.林业科学,2014,50(12):24-31. Mi N,Wen X F,Cai F,Wang Y,Zhang Y S.Effects of seasonal drought on the water use efficiency of Qianyanzhou plantation.Scientia Silvae Sinicae,2014,50(12):24-31.(in Chinese) [45] Yu G,Song X,Wang Q,Liu Y,Guan D,Yan J,Sun X,Zhang L,Wen X.Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables.New Phytologist,2008,177(4):927-937. [46] Zhao B,Kondo M,Maeda M,Ozaki Y,Zhang J.Water-use efficiency and carbon isotope discrimination in two cultivars of upland rice during different developmental stages under three water regimes.Plant and Soil,2004,261(1-2):61-75. [47] 刘贤赵,康绍忠.遮荫对番茄单叶水分利用效率的影响研究.中国生态农业学报,2003,11(1):23-26. Liu X Z,Kang S Z.Effects of environment factors under shading conditions on water use efficiency of tomato leaves.Chinese Journal of Eco-Agriculture,2003,11(1):23-26.(in Chinese) [48] 曹生奎,冯起,司建华,常宗强,卓玛错,席海洋,苏永红.植物叶片水分利用效率研究综述.生态学报,2009,29(7):3882-3892. Cao S K,Feng Q,Si J H,Chang Z Q,Zhuomacuo,Xi H Y,Su Y H.Summary on the plant water use efficiency at leaf level.Acta Ecologica Sinica,2009,29(7):3882-3892.(in Chinese) [49] Chi Y K,Xiong K N,Zhao P D,Zhang J H,Wang Y S,Zhou W L.Analysis and study on photosynthetic physiological characteristics of 4 grass species in karst rocky desertification area.Journal of Animal and Veterinary Advances,2013. [50] 杨有俊,郑明珠,秦伟志,陈焘,江世高,刘金荣.干旱及复水处理下坪用黑麦草和高羊茅抗旱特性比较.草业科学,2012,29(3):370-376. Yang Y J,Zheng M Z,Qin W Z,Chen T,Jiang S G,Liu J R.Response of tall fescue and perennial ryegrass turfgrasses to drought stress and re-watering treatment.Pratacultural Science,2012,29(3):370-376.(in Chinese) [51] Pirnajmedin F,Majidi M M,Gheysari M.Root and physiological characteristics associated with drought tolerance in Iranian tall fescue.Euphytica,2015,202(1):141-155. [52] Bonos S A,Rush D,Hignight K,Meyer W A.Selection for deep root production in tall fescue and perennial ryegrass.Crop Science,2004,44(5):1770-1775. (责任编辑 苟燕妮) Effect of decline in soil depth and water resource on the photosynthesis of two grasses under mixed plantation in Karst regions Zhao Ya-jie, Li Zhou, Song Hai-yan, Zhang Jing, Liang Qian-hui, Liu Jin-chun Uneven soil depth and karst drought are key ecological factors affecting plant survival, growth, and reproduction in karst regions. The change in global precipitation patterns will exacerbate the frequency and intensity of karst drought. The amount of water and nutrients absorbed and transported to the aboveground parts by roots systems can affect plant photosynthesis. Shallow-rooted plants mainly use water and nutrients from the top layers of the soil, whereas deep-rooted plants can use water and nutrients from the deeper soil layers. Therefore, when deep-rooted plants and shallow-rooted plants coexist, they might alleviate the inhibitory effect of resource limitation on plant photosynthesis because of their complementary root niche. We grewFestucaarundinacea(deep-rooted) andLoliumperenne(shallow-rooted) under normal versus shallow soil depth and well watered versus drought treatments in a block design, and studied the effect of the decline in soil depth, water availability, and both these resources (soil depth plus water availability) on the photosynthesis of two grasses under mixed plantation by controlled experiment. The results showed that: 1) at high resource levels, no significant difference occurred in net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), and water use efficiency (WUE) ofF.arundinaceabetween mixed plantation and in monoculture (P>0.05); Pn, Gs, and TrofL.perennewas lower in mixed plantations than in monoculture, whereas WUE was significantly higher in mixed plantations than in monoculture (P>0.05). 2) Overall, Pn, Gs, and TrofF.arundinaceaandL.perennedecreased and WUE ofL.perenneincreased with the decline of resources (soil depth, water, and soil depth plus water) either in mixture or in monoculture. Moreover, the range of decrease or increase was lower in mixed plantations than in monoculture; WUE ofF.arundinaceaincreased in monoculture but decreased in mixed plantations. 3) At low resource levels, Pn, Gs, and TrofF.arundinaceawas higher in mixed plantations than in monoculture, whereas WUE was lower in mixed plantations than in monoculture. No significant difference was found in Pn, Gs, and TrofL.perennebetween mixed plantations and monoculture, whereas WUE was higher in mixed plantations than in monoculture. Our results showed that owing to the complementary niche and hydraulic lift of deep-rooted plants, the inhibitory effect of resource limitation on photosynthesis of the deep-rooted and shallow-rooted plants are alleviated when they are planted together, but at the expense of WUE. Therefore, in karst regions, mixed planting with plant of different root depth systems may be an effective method of resolving the increasing drought and poor soil conditions. karst drought; soil heterogeneity; photosynthesis; water use efficiency; niche complementary; deep-rooted plant; shallow-rooted plant Liu Jin-chun E-mail:jinchun@swu.edu.cn 2016-10-17 接受日期:2017-01-03 国家自然科学基金项目(31500399);重庆市自然科学基金项目(cstc2014jcyjA80016);教育部第49批留学回国人员科研启动基金项目 赵雅洁(1994-),女,安徽蚌埠人,在读硕士生,主要从事植物生理生态学研究。E-mail:zhaoyajie1994@126.com 刘锦春(1977-),女,湖南洞口人,副教授,博士,主要从事植物生理生态和植被恢复研究。E-mail:jinchun@swu.edu.cn 10.11829/j.issn.1001-0629.2016-0533 S812.2;Q945.11 A 1001-0629(2017)07-1475-12 赵雅洁,李周,宋海燕,张静,梁千慧,刘锦春.喀斯特地区土壤厚度降低和水分减少对两种草本植物混种后光合的影响.草业科学,2017,34(7):1475-1486. Zhao Y J,Li Z,Song H Y,Zhang J,Liang Q H,Liu J C.Effect of decline in soil depth and water resource on the photosynthesis of two grasses under mixed plantation in Karst regions.Pratacultural Science,2017,34(7):1475-1486.

3 讨论


4 结论
(Key Laboratory of Eco-environment in Three Gorges Reservoir Region Ministry of Education, Chongqing Key Laboratory of Plant Ecology and Resources Research in Three Gorges Reservoir Region, School of life Sciences, Southwest University, Chongqing 400715, China)
