富硒对蛹虫草菌丝干重及胞内酶活性的影响
2017-08-09张曦文张国财刘春延李文娟东北林业大学林学院黑龙江哈尔滨150040
张曦文,赵 博,张国财,刘春延,李文娟(东北林业大学林学院,黑龙江哈尔滨 150040)
富硒对蛹虫草菌丝干重及胞内酶活性的影响
张曦文,赵 博,张国财*,刘春延,李文娟
(东北林业大学林学院,黑龙江哈尔滨 150040)
在培养基中添加不同浓度的Na2SeO3液体培养蛹虫草,探究富硒对蛹虫草菌丝干重及胞内酶活性的影响,并进行相关性分析。研究结果表明:当硒添加量为3 mg/kg时,菌丝干重达1.51 g/100 mL,较对照组高19.84%;当硒添加量为7 mg/kg时,菌丝干重较对照组低4.76%,过高的硒添加量会抑制蛹虫草的生长。适宜的硒添加量能在不同程度上提高胞内酶活性,但促进效果存在差异。菌丝干重与蛋白酶活性呈正相关(p<0.01),R2=0.980,表明蛋白酶是富硒蛹虫草生长中的关键酶。当硒添加量为3 mg/kg时,菌丝干重及蛋白酶活性均达到最大值,可以为富硒蛹虫草液体发酵提供理论依据。
富硒,蛹虫草,胞内酶活性,菌丝干重
蛹虫草[Cordycepsmilitaris(Vuill.)Fr.]是麦角菌科(Clavicipitaceae)虫草属(Cordyceps)的模式种[1],富含虫草素、虫草酸、虫草多糖等多种对人体有益的活性物质及微量元素,是冬虫夏草的理想替代品[2]。硒(Selenium,Se)是人体必需的微量元素之一,在延缓衰老、防癌抗癌、增强机体免疫力等方面发挥着重要作用[3]。目前,世界各地普遍存在硒摄入量不足的问题,缺硒会引发多种疾病,但无机硒难以直接吸收,且具有一定的毒性,因此,开发有机富硒保健品势在必行[4]。近年来,富硒酵母[5]、富硒茶叶[6]、富硒大米[7]、富硒食用菌[8]等富硒食品的开发取得了一定的进展。富硒食用菌是一种将无机硒与食用菌有机结合的产物,可以将无机硒转化为有机硒,使硒更容易被人体组织储存和吸收,是人体良好的补硒剂[9-12]。许多科研工作者对富硒蛹虫草进行了较深入的研究,包括富硒蛹虫草栽培技术、活性成分及药理作用等[13-18],但对富硒蛹虫草胞内酶活性机理的研究未见报道。因此,系统地研究硒浓度对蛹虫草胞内酶活性的影响,对富硒蛹虫草保健品的开发具有重要的理论价值和生产实践意义[19-21]。本文拟在培养基中添加Na2SeO3溶液培养蛹虫草,探究富硒对蛹虫草菌丝干重及胞内酶活性的影响。
1 材料和方法
1.1 材料与仪器
蛹虫草母种 由东北林业大学森林保护学重点实验室保藏;无水亚硒酸钠、葡萄糖、蛋白胨、琼脂、磷酸二氢钾、磷酸二氢钠、硫酸镁 均为分析纯,国药集团化学试剂有限公司。
BiofugeStratos高速冷冻离心机 赛默飞世尔科技有限公司;DK-S14型电热恒温水浴锅 上海森信实验仪器有限公司;UV-5100 型紫外分光光度计 上海元析仪器有限公司;F-2500型荧光分光光度计 日立公司;SHB-ⅢA型循环水式真空泵 郑州长城科工贸有限公司。
1.2 实验方法
1.2.1 富硒蛹虫草培养基的制备 配方组成:马铃薯200 g/L,葡萄糖20 g/L,维生素B150 mg/L,蛋白胨5 g/L,酵母粉3 g/L,磷酸二氢钾2 g/L,硫酸镁1 g/L,配制培养液[22]。在培养液中添加Na2SeO3溶液,使培养液Na2SeO3终浓度分别为0、1、3、5、7 mg/kg。用500 mL三角瓶分装培养液,每瓶250 mL,121 ℃灭菌20 min后备用。
1.2.2 菌株活化及培养
1.2.2.1 菌株活化 在超净工作台中,将菌种接种于培养基配方为马铃薯200 g/L,葡萄糖20 g/L,琼脂粉16 g/L,蛋白胨5 g/L,磷酸二氢钾2 g/L,硫酸镁1 g/L的平面培养基中,22 ℃条件下培养10 d,制得活化菌株。
1.2.2.2 菌株培养 在无菌条件下,将各活化后的菌株用打孔器打成大小相同直径为5 mm菌块,并分别接种于不同Na2SeO3终浓度的富硒培养液及不加Na2SeO3的对照培养液中,每菌株设置重复3个,每瓶接入菌块3个,并置于摇床中在20 ℃,160 r/min条件下,恒温振荡培养[22]。
1.2.3 菌丝干重测定 按1.2.2方法连续培养蛹虫草4 d,真空抽滤培养液,并用等量的蒸馏水清洗3次,将所得菌丝置于60 ℃干燥箱中烘干至恒重,分析天平称量即为菌丝干重,每个处理3个重复。
1.2.4 菌丝总硒含量测定 根据食品安全国家标准食品中硒的测定[23]。
1.2.5 粗酶液的制备及胞内酶活测定
1.2.5.1 粗酶液的制备 按时观察1.2.2中培养的菌丝,待菌丝萌发后开始取样,取样时间间隔为12 h。具体方法是将培养的样品分装到50 mL离心管中,于4 ℃条件下10000 r/min离心15 min后弃上清液,合并菌丝,而后再用等量蒸馏水清洗菌丝三次后,将菌丝置于冰浴磷酸缓冲液中研磨破壁。吸取研磨匀浆,离心后上清液即得所需粗酶液[22]。
1.2.5.2 胞内酶活测定 蛋白酶采用福林酚法测定[24],即在试管中加入预热的2%酪蛋白溶液及粗酶液各1 mL,混合均匀后40 ℃恒温水浴10 min,取出后立即加入0.4 mol/L三氯醋酸溶液2 mL以终止反应。离心后取上清液0.5 mL并依次加入2.5 mL 0.4 mol/L碳酸钠溶液及0.5 mL福林酚工作液40 ℃恒温水浴显色20 min,而后于680 nm测定吸光度。以每毫升样品与底物反映10 min改变0.01个吸光度作为一个酶活力单位进行酶活计算。淀粉酶、多酚氧化酶、过氧化物酶、超氧化物歧化酶均采用南京建成生物研究所试剂盒测定,按照说明书进行操作。
1.3 数据处理
实验所得数据利用SPSS 22.0软件进行处理。
2 结果与分析
2.1 硒对蛹虫草菌丝干重及总硒量的影响
2.1.1 硒对菌丝干重的影响 由图1可知,在相同条件下,不同硒添加量培养蛹虫草,菌丝干重随硒添加量的增加整体呈先上升后下降趋势,即当硒添加量为1~3 mg/kg时,菌丝干重随硒添加量的增加而增加,3 mg/kg菌丝干重达1.51 g/100 mL,相比对照组提高了19.84%。当硒添加量为5 mg/kg时,菌丝干重相比硒添加量为3 mg/kg降低了1.99 %,但仍高于对照组。当硒添加量为7 mg/kg时,菌丝干重仅为1.20 g/100 mL,相比对照组降低了4.76%,对蛹虫草菌丝生长显示出了明显的抑制作用。方差分析结果表明,5个处理组菌丝干重间均存在显著差异。因此,仅以菌丝产量单因素分析,在进行蛹虫草富硒培养时,发酵液中最佳硒浓度为3 mg/kg。
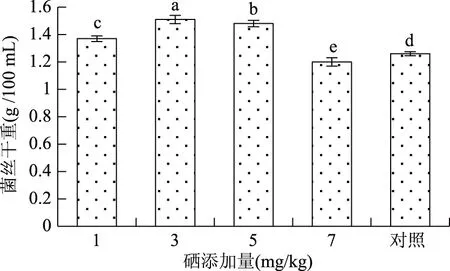
图1 不同硒添加量菌丝干重Fig.1 Mycelial dry weight in different amount of selenium注:小写字母表示差异在0.05水平显著,图2~图7同。
2.1.2 硒对总硒含量的影响 由图2可知,蛹虫草具有较强的富硒能力,通过发酵液中添加硒可以提高菌丝中的硒含量,且硒含量与添加量呈正相关。当硒添加量为7 mg/kg时,菌丝硒含量最高,为165.23 mg/kg。方差分析表明,各处理组间硒含量均存在显著差异。
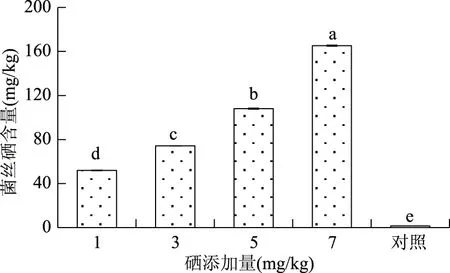
图2 不同硒添加量的总硒含量Fig.2 The total selenium content of different selenium additions
2.2 硒对蛹虫草菌丝胞内酶活性的影响
2.2.1 硒对胞内蛋白酶活性的影响 由图3可知,当硒添加量为1、3、5 mg/kg时,其酶活高峰相比对照组分别提高了16.48%、25.82%、2.60%,富硒液体发酵有利于提高蛹虫草菌丝细胞内蛋白酶的活性。当硒添加量为7 mg/kg时,富硒液体发酵对蛹虫草菌丝蛋白酶活性前期有抑制作用,后期有增强作用。由此说明,适宜低浓度的Na2SeO3可以促进蛹虫草胞内蛋白酶的分泌,提高酶的活性。其中当硒添加量为3 mg/kg时,促进效果最为显著。
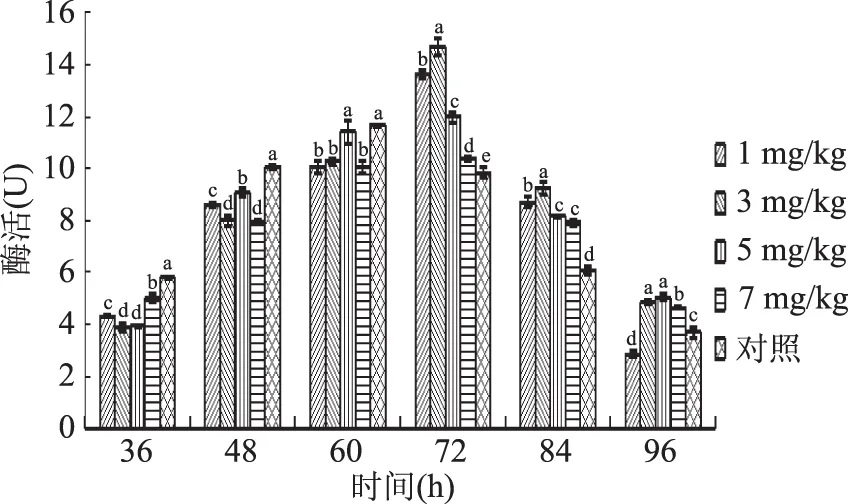
图3 不同硒添加量液体培养蛹虫草胞内蛋白酶活性变化趋势Fig.1 Changes of protease activity in C.militaris under different selenium additions
2.2.2 硒对胞内淀粉酶活性的影响 由图4可知,在整个液体培养过程中,5个处理组的蛹虫草胞内淀粉酶活性整体变化趋势基本一致,即酶活性自培养的开始阶段便快速分泌,直至第60 h大部分出现酶活高峰,随后即有所下降,至第84 h后出现另一酶活高峰。
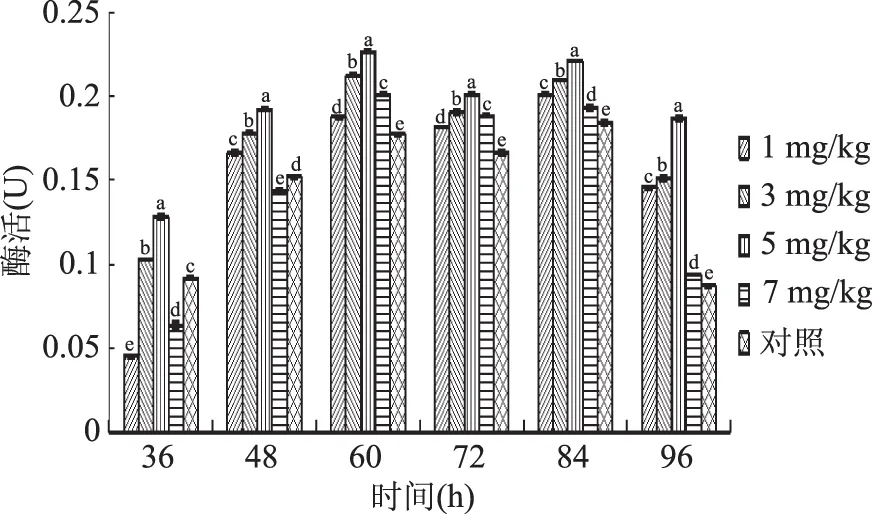
图4 不同硒添加量液体培养蛹虫草胞内淀粉酶活性变化趋势Fig.4 Changes of intracellular amylase activity of C.militaris cultured with different selenium additions
当硒添加量为1~7 mg/kg时,从培养中期开始蛹虫草胞内淀粉酶活性均高于对照组。其中,以硒添加量5 mg/kg处理组促进作用最明显,酶活高峰达0.2264 U,相比对照组和其他处理组分别提高了22.78%、12.52%、6.64%、12.47%。与对照相比,当硒添加量为7 mg/kg时,36~48 h促进作用不显著,但后期促进作用显著,表现出了明显的滞后性,这与蛋白酶活性测定结果相似。由此说明,当硒添加量为1~7 mg/kg时,富硒液体发酵均有利于提高蛹虫草菌丝细胞内淀粉酶活性,促进作用随着硒添加量的增加呈先上升后下降趋势,以5 mg/kg处理组效果最为显著。
2.2.3 硒对胞内多酚氧化酶活性的影响 由图5可知,在整个培养过程中,5个处理组的蛹虫草多酚氧化酶活力变化趋势基本保持一致,自培养开始酶活力逐渐升高,1、3、5 mg/kg及对照组至84 h达到活性顶点随后下降。7 mg/kg处理组酶活至84 h时,酶活快速上升,显示出了明显的滞后性。
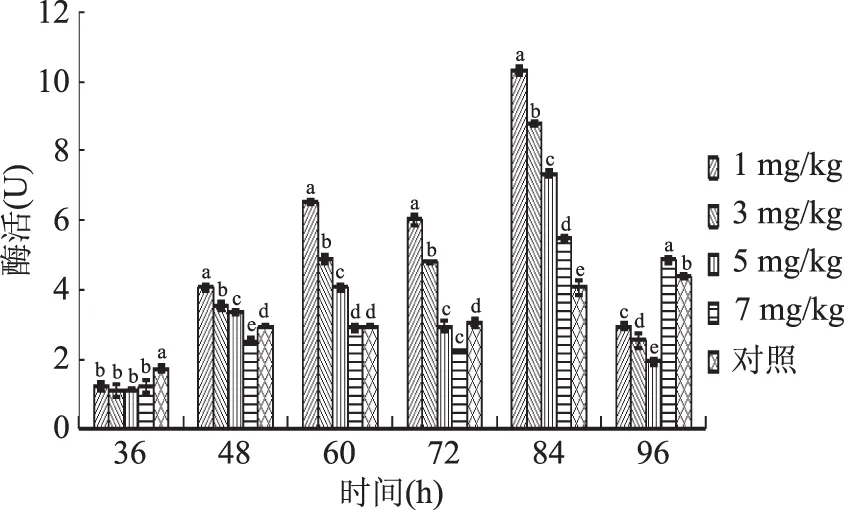
图5 不同硒添加量液体培养蛹虫草胞内多酚氧化酶活性变化趋势Fig.5 Changes of intracellular polyphenol oxidase activity in C.militaris under different selenium additions
当硒添加量为1、3、5 mg/kg时,其多酚氧化酶高峰活性均显著高于对照组,相比对照组分别提高了134.85%、100%和67.43%;而当亚硒酸钠浓度为7 mg/kg时,其多酚氧化酶活性在发酵前72 h均略低于对照组,此后酶活性快速增加,并显著高于对照组酶活性。由此可知,当亚硒酸钠浓度为1~5 mg/kg时,富硒液体发酵可显著提高多酚氧化酶的活性,且硒添加量为1 mg/kg时提高效果最明显;而当亚硒酸钠浓度为7 mg/kg时,富硒液体发酵对蛹虫草菌丝细胞内多酚氧化酶活性在发酵初期具有一定的抑制作用,后显示出促进作用。
2.2.4 硒对胞内过氧化物酶活性的影响 由图6可知,蛹虫草富硒液体发酵过程中,当硒添加量为1~7 mg/kg时,蛹虫草胞内过氧化物酶活性变化整体趋于一致,均于72 h达到酶活高峰,随后缓慢下降。与此同时,1、3、5 mg/kg处理组其胞内过氧化物酶活性均高于对照组,5 mg/kg处理组酶活最高,其酶活峰值达63.20 U,相比1、3 mg/kg及对照组分别提高了6.81%、4.26%、9.08%。当硒添加量为7 mg/kg时,相比对照组酶活性有所降低,显示出了一定的抑制作用。由此说明,当硒添加量为1、3、5 mg/kg时,富硒液体发酵均可促进蛹虫草胞内过氧化物酶的分泌,提高其活性,但当硒添加量为7 mg/kg时,富硒液体发酵对蛹虫草菌丝胞内过氧化物酶活性具有抑制作用。

表1 蛹虫草胞内酶活性与菌丝干重相关性分析Table 1 Correlation analysis within intracellular enzyme activities and mycelial dry weight of C. militaris
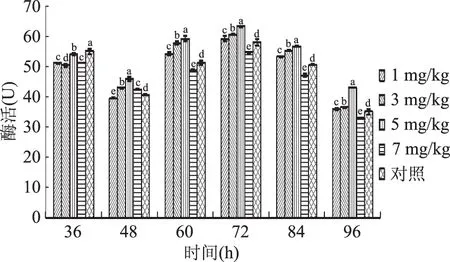
图6 不同硒添加量液体培养蛹虫草胞内过氧化物酶活性变化趋势Fig.6 Changes of peroxidase activity in C. militaris under different selenium additions
注:**表示在0.01水平显著。
2.2.5 硒对胞内超氧化物歧化酶活性的影响 由图7可知,富硒液体发酵过程中,蛹虫草菌丝细胞内超氧化物歧化酶活性随着时间推移整体呈下降趋势。在1~7 mg/kg的范围内,随着硒添加量的增加,超氧化物酶活性峰值随浓度增大而增大,且均高于对照组超氧化物歧化酶活性,在24 h时相比对照组分别增加了1.54%、2.38%、3.50%、22.37%。由此说明,硒添加量为1~7 mg/kg范围内时,富硒液体发酵培养可以提高超氧化物歧化酶的活性,且亚硒酸钠浓度越高,富硒液体发酵对超氧化物歧化酶活性提高效果越显著。
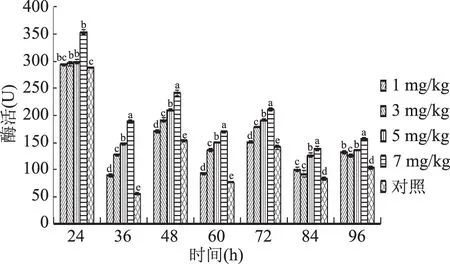
图7 不同硒添加量液体培养蛹虫草胞内超氧化物歧化酶活性Fig.7 Effects of different selenium additions on the activity of superoxide dismutase in C.militaris
2.3 相关性分析
由表1可知,蛋白酶活性、淀粉酶活性、多酚氧化酶、过氧化物酶活性与菌丝干重呈正相关,且蛋白酶活性与菌丝干重在0.01水平极显著。说明这四种酶对菌丝的生长有一定的促进作用,蛋白酶与菌丝干重的关系更为密切,蛋白酶是富硒蛹虫草液体培养的关键酶。结合图1和图7可知,当硒添加量为1~5 mg/kg时,超氧化物歧化酶活性和菌丝干重随着硒添加量的增大而增大,呈正相关;当硒添加量为7 mg/kg时,超氧化物歧化酶活性依然在上升,但菌丝干重下降,可能是因为菌丝受到不利因素的影响导致菌丝干重下降,从而促进保护性的超氧化物歧化酶分泌。
3 讨论与结论
食用菌生长发育与胞内酶的分泌及活性变化存在重要关系,研究食用菌胞内酶活性的变化对筛选优良菌株,提高菌株产量和品质有重要意义[25-26]。在硒添加量为3 mg/kg时,菌丝干重较对照组高19.84%。当硒添加量为7 mg/kg时,菌丝干重仅为1.20 g/100 mL,较对照组低4.76%,较高硒添加量对蛹虫草菌丝生长显示了明显的抑制作用。
富硒能促进蛹虫草胞内蛋白酶、淀粉酶、多酚氧化酶、过氧化物酶、超氧化物歧化酶活性,可促进酶的分泌及增强酶的活性[27-28]。不同硒添加量对酶的作用效果存在差异性,当硒添加量为1 mg/kg时,多酚氧化酶高峰活性较对照组高134.85%;当硒添加量为3 mg/kg时,蛋白酶活性高峰较对照组高25.79%;当硒添加量为5 mg/kg时,淀粉酶和过氧化物酶活性高峰较对照组分别高22.78%和9.08%;当硒添加量为7 mg/kg时,超氧化物歧化酶高活性峰较对照组高22.37%。其原因可能是无机硒通过培养基进入菌丝体后转化成有机硒,促进菌丝生长,从而增强了酶的活性。
通过对蛹虫草菌丝干重与胞内酶活性相关性分析表明:菌丝干重与胞内蛋白酶活性呈显著正相关(p<0.01),表明蛋白酶是富硒蛹虫草液体发酵的关键酶。其原因是蛋白质是菌丝中硒结合的主要营养物质,富硒会促进硒与蛋白质结合,增强了蛋白酶活性[29]。蛹虫草菌丝干重与蛋白酶、淀粉酶、多酚过氧化物酶及过氧化物酶活性均呈正相关,而超氧化物歧化酶活性呈负相关,这可能是因为超氧化物歧化酶是保护性酶,当这种酶活性变强,说明菌丝受到不利因素的影响,过高的硒浓度对菌丝产生一定的抑制作用,导致菌丝干重下降。
综上所述,适当的硒添加量可促进蛹虫草菌丝生长,提高蛹虫草液体发酵产量,提高胞内酶活性。蛋白酶是富硒蛹虫草生长中的关键酶,当硒添加量为3 mg/kg时,菌丝干重及蛋白酶活性达到最大值,研究结果可为富硒蛹虫草液体发酵提供理论依据。
[1]Kirk,Cannon PF,David JC,et al. Ainsworth & Bisby’s Dictionary of the Fungi[M]. CABI Publishing,2001.
[2]Dong J Z,Ding J,Pei Z Y,et al. Composition and distribution of the main active components in selenium-enriched fruit bodies ofCordycepsmilitarislink[J]. Food Chemistry,2013,137(1):164-167.
[3]顾浩峰. 食品中硒元素的形态分析研究进展[J]. 食品与发酵科技,2015(2):1-8.
[4]徐庆国,刘红梅,黄丰. 我国富硒农产品开发与推广的探讨[J]. 作物研究,2013,27(5):461-464.
[5]Demirci A,Pometto A L,Cox D J. Enhanced organically bound selenium yeast production by fed-batch fermentation[J]. Journal of Agricultural and Food Chemistry,1999,47(6):2496-2500.
[6]Wang Y,Li Y,Liu Y,et al. Extraction,characterization and antioxidant activities of Se-enriched tea polysaccharides[J]. International Journal of Biological Macromolecules,2015,77:76-84.
[7]李丽彩,王亚婷,赵建宏,等. 富硒麦芽的生产工艺研究[J]. 食品科技,2016(1):64-67.
[8]林佳,李昌发. 富硒食用菌优势与生产研究[J]. 企业技术开发,2013,1:130-131.
[9]Elbayoumy K,Sinha R. Mechanisms of mammary cancer chemoprevention by organo-selenium compounds[J]. Mutat Res,2004,46(551):181-197
[10]刘春延,赵博,张国财,等. 富硒黑木耳硒多糖超声-微波提取工艺优化及其抗氧化活性[J]. 湖南农业大学学报:自然科学版,2016,42(4):435-440.
[11]李玲飞,蔡松伟,吴根福. 富硒食用菌及其保健功效[J]. 食药用菌,2011,19(6):38-43.
[12]Lintschingeret al. Selenium-enriched sprouts. A raw material for fortified cereal-baded diets[J]. J Agric Food Chem,2000,48:5326-5368.
[13]龙思颖,康德灿,柯江,等. 液体发酵培养富硒蛹虫草菌丝体优化条件的实验[J]. 食用菌,2016(3):11-13.
[14]张国财,赵博,刘春延,等. 响应面法优化超声波-微波协同提取富硒蛹虫草硒多糖工艺[J]. 食品科学,2016,37(12):33-39.
[15]于田田,钱和. 生物富硒对蛹虫草菌丝体化学成分的影响[J]. 食品科技,2006,2006(1):133-135.
[16]朱蕴兰,陈宏伟,张城. 富硒蛹虫草胞内多糖对自由基的清除作用[J]. 农业工程,2011(3):53-57.
[17]钟鸣,高丹,胡昆,等. 富硒蛹虫草对小鼠降血脂和抗氧化作用的影响[J]. 时珍国医国药,2009,20(12):3142-3144.
[18]Das S K,Masuda M,Sakurai A,et al. Medicinal uses of the mushroomCordycepsmilitaris:current state and prospects[J]. Fitoterapia,2010,81(8):961-968.
[19]李海洲,陈正余,张京国,等. 安康富硒蛹虫草黄酒生产工艺探讨[J]. 安康学院学报,2015,27(6):71-73.
[20]张园园,王勇,叶岚,等. 蛹虫草富硒研究及富硒产品开发初探[J]. 食用菌,2014(4):3.
[21]孟泽彬,文庭池,姜金仲. 富硒虫草的研究进展[J]. 微生物学通报,2014,41(11):2339-2348.
[22]赵博,张国财,林连男,等. 生物磁效应对蛹虫草活性物质含量的影响[J]. 植物研究,2013,33(4):508-511.
[23]2010,GB5009 93. 食品安全国家标准食品中硒的测定[S]. Diss. 2010.
[24]程功. 生物磁效应对杏鲍菇液体菌种培养及胞外酶的作用[D]. 哈尔滨:东北林业大学,2007.
[25]杨淑云,刘朝贵,朱凯,等. 富硒姬松茸液体培养条件的研究[J]. 微生物学通报,2003,30(1):9-13.
[26]王孟兰,赵妍,陈明杰,等. 食用菌半纤维素酶系研究进展[J]. 生物学杂志,2014,31(1):64-67.
[27]Wang X,Zuo Z,Zhao C,et al. Protective role of selenium in the activities of antioxidant enzymes in piglet splenic lymphocytes exposed to deoxynivalenol[J]. Environmental Toxicology and Pharmacology,2016,47:53-61.
[28]Adebayo O L,Adenuga G A,Sandhir R. Selenium and zinc protect brain mitochondrial antioxidants and electron transport chain enzymes following postnatal protein malnutrition[J]. Life Sciences,2016,152:145-155.
[29]赵镭. 灵芝生物富硒及富硒灵芝硒蛋白的分离纯化和抗氧化性研究[D].北京:中国农业大学,2004.
Effects of selenium enrichment on mycelia dry weight and intracellular enzyme activity ofCordycepsmilitaris
ZHANG Xi-wen,ZHAO Bo,ZHANG Guo-cai*,LIU Chun-yan,LI Wen-juan
(College of Forestry,Northeast Forestry University,Harbin 150040,China)
The effects of selenium enrichment on the mycelia dry weight and intracellular enzyme activity ofCordycepsmilitariswere investigated and the correlation analysis was carried out by adding different concentrations of Na2SeO3liquid to the culture medium. The results showed that when the selenium content was 3 mg/kg,the dry weight of mycelium was 1.51 g/100 mL,which was 19.84% higher than that of the control.When the selenium content was 7 mg/kg,the dry weight of mycelium was 4.76% lower than that of the control.Excessive selenium addition inhibits the growth ofCordycepsmilitaris. The suitable amount of selenium additions could increase the activity of intracellular enzymes in different extent,but the promoting effect was different. At thep<0.01 level,the dry weight of mycelia was positively correlated with the protease activity,R2=0.980,indicating that protease was the key enzyme in the growth of Se-enrichedCordycepsmilitaris.When the selenium content was 3 mg/kg,the mycelium dry weight and protease activity reached the maximum value,which could provide the theoretical basis for liquid fermentation of selenium-enrichedCordycepsmilitaris.
selenium-enriched;Cordycepsmilitaris;intracellular enzyme activities;mycelial dry weight
2017-01-05
张曦文(1992-),女,硕士研究生,研究方向:食用菌栽培及药理,E-mail:zxw920427@163.com。
*通讯作者:张国财(1964-),男,博士,研究方向:食用菌栽培技术,E-mail:zhang640308@126.com。
中央高校基本科研业务费专项(2572014AA08)。
TS209
A
1002-0306(2017)14-0151-05
10.13386/j.issn1002-0306.2017.14.030
