长喙毛茛泽泻不同海拔种植试验
2017-08-09陆丽萍季志利王武军杜有新何小勇
陆丽萍,季志利,王武军,杜有新,何小勇
(1. 丽水莲都林场,浙江 丽水 323000;2. 莲都区林业技术推广站,浙江 丽水 323000;3. 丽水市林业科学研究院,浙江 丽水 323000;)
长喙毛茛泽泻不同海拔种植试验
陆丽萍1,季志利1,王武军2,杜有新3,何小勇3
(1. 丽水莲都林场,浙江 丽水 323000;2. 莲都区林业技术推广站,浙江 丽水 323000;3. 丽水市林业科学研究院,浙江 丽水 323000;)
2016年5月,在浙江省丽水市莲都林场选择不同海拔高度试验地进行濒危植物长喙毛茛泽泻种植试验,同年7-11月观察记录种群变化并计算其繁殖系数。结果表明,长喙毛茛泽泻在海拔80 ~ 1 040 m不同海拔种植均可成活,平均繁殖系数2.75 ~ 39.80,以海拔840 m的繁殖系数最高;不同海拔之间平均株高、地茎、叶片大小、根系长度、纤匍茎节间长均存在极显著差异;2016年11月测量单株生物量,结果显示单株总生物量0.18 ~ 0.25 g,从低海拔到高海拔生境总生物量明显增大,地下部分占比明显增加。
长喙毛茛泽泻;海拔梯度;种植试验;植株形态;生物量
Kew words: Ranalisma rostratum; altitudinal gradient; planting experiment; plant morphology; biomass
长喙毛茛泽泻Ranalisma rostratum Stapf是泽泻科Alismataceae毛茛泽泻属Ranalisma多年生的水生小草本植物,具纤匐枝,叶基生,叶柄细,中空,基部鞘状,叶片宽椭圆形或卵状椭圆形,膜质,顶端锐尖,基部心形或钝,具纤毛。长喙毛茛泽泻在中国产浙江丽水,在越南、印度、马来西亚和非洲热带地区也有分布,在分类学上有特殊的地位,为国家一级保护野生植物[1-2]。长喙毛茛泽泻有两种繁殖方式,由于种子小且没有胚乳,导致种子萌发率和幼苗存活率都很低;无性繁殖是种群维持的主要方式,从植株基部发出纤匍枝卧于水中或地表,纤匍茎上每隔2 ~ 15 cm产生莲座状无性小株,纤匍枝自然断裂后成为独立植株[3]。1930年在浙江省丽水的南明山(119°55′ E ,28°27′ N)曾采到一份长喙毛茛泽泻标本,但之后未再发现这种植物,一度认为在我国已绝灭[4],直到1999年在该地水池中再次发现长喙毛茛泽泻小种群,但由于水质变化,2001年后再次消失。关于长喙毛茛泽泻的研究主要集中在其生物学特性[3]、生殖生物学[4-8]、濒危机制[9-11]等方面,而关于其种群回归种植的研究极少[12]。通过不同海拔条件下的种植试验,为在自然条件下的长喙毛茛泽泻扩繁和回归提供依据。
1 材料与方法
1.1 试验区基本情况
试验地位于浙江省丽水市莲都林场(119°55′ E,28°28′ N),属中亚热带季风气候区,四季分明,气候温和,无霜期长。林场地形起伏较大,从海拔60 m的盆地到海拔1 000余m的中山,最高峰葑垟尖海拔1 326 m。林场总部年平均气温14℃,极端最高气温40.5℃,极端最低气温-12℃,日平均气温≥10℃的227 d,日平均气温≤0℃的41 d,年平均降水量2 069.5 mm,年平均相对湿度87%,年平均日照时数1 098 h。
1.2 试验材料
长喙毛茛泽泻种源来自杭州植物园和浙江人文园林天景水生植物园,是人工迁地培育种群,生长良好。种苗采自丽水市莲都区老竹镇后坑村扩繁种植点。该地海拔119 m,位于公路旁,距离老竹镇1 km,原种植莲花,后经过平整、切坎改造,形成水池,引进附近水库达饮用水标准的天然水源。2015年5月进行种植。
1.3 试验方法
种植地选择。在莲都林场不同的海拔选取5个地点进行种植试验。选取的试验点均为湿地,长年基本有水。各试验点的生境情况如表1。

表1 不同海拔长喙毛茛泽泻试验地基本情况Table 1 Information of experiment plots for planation of R. rostratum at different altitude
种植管理。2016年5月种植。每个试验点按20 cm×20 cm行间距种植10 ~ 18株。种植前除去试验地里的所有杂草,并进行中耕。种植后根据生长情况及时清除杂草,捉去害虫。
观测方法。长喙毛茛泽泻种植后,各试验点分别于2016年7月5日、8月20日、10月7日及11月29日,统计包括母株及其分蘖子株的个体数量,计算繁殖系数。2016年的11月29日,在各试验点采取网格法随机采集20株,洗净泥土,分别测量地上部分的株高、叶片长宽、纤匍茎节间长度和地下根系长度,观察叶色。所有样品在80℃烘箱内烘干10 h至恒重后称重,按地上和地下两部分分开计算生物量,地上部分包括地茎和叶片,地下部分包括根系和纤葡茎,整合计量单株生物量。
1.4 数据分析
繁殖系数按如下公式计算:
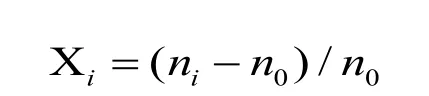
式中,i为调查月份,n0为种植时株数,ni为第i个月调查的株数,Xi为第i个月的繁殖系数。所有数据应用Microsoft Excel 2003进行数理统计,DPS数据系统进行方差分析。
2 结果与分析
2.1 不同海拔对长喙毛茛泽泻生长及繁殖系数的影响
长喙毛茛泽泻在不同海拔试验点种植后均成活,一个月后即开始以纤匍茎的方式进行无性繁殖。各个试验点的不同时间植株数量及其繁殖系数见表2。

表2 不同海拔长喙毛茛泽泻种群株数变化及繁殖系数Table 2 Number of individuals and reproduction coefficient of R. rostratum at different altitude
从表2中可以看出,长喙毛茛泽泻种植后,各试验点繁殖系数从7月5日的0.14 ~ 0.80,8月20日的1.57 ~25.00,到10月7日达到2.50 ~ 34.00,11月29日达到2.75 ~ 39.80。从增殖速率来看,7-8月是增殖高峰期,随着气温逐渐下降,10月后增殖速率放缓。进入秋季后,由于天气干燥,土壤缺水导致葑垟湖试验地上植株出现部分枯死现象;九户垄试验地后期出现了田螺的侵害,导致泽泻植株数略有下降。各个时期均以海拔840 m的圩地种植点繁殖系数最高,其它各海拔高度的种植试验点差异不大,这主要与圩地种植点主要是山泉水,水质优良有关。
2.2 不同海拔对长喙毛茛泽泻植株及其器官形态的影响
不同海拔长喙毛茛泽泻的植株和叶片形态变化特征见表3。长喙毛茛泽泻在各试验点经过一个生长期的生长,平均株高7.1 ~ 13.5 cm,平均地茎粗0.38 ~ 0.52 cm,不同海拔间株高和茎粗存在极显著差异。海拔80 m的试验点植株最高达13.5 cm,其次为840 m试验点,株高13.4 cm,极显著大于其它试验点;但是海拔840 m试验点地茎最小,只有0.38 cm,显著小于其它试验点。
长喙毛茛泽泻在不同生境生长的叶片叶色均为淡绿色或绿色,没有多大差异。叶片平均长2.1 ~ 3.3 cm,平均宽1.7 ~ 2.9 cm,不同生境间叶片长宽存在极显著差异。海拔80 m的源头弄试验点叶片最长3.3 cm,其次为海拔840 m圩地试验点叶长3.0 cm,极显著长于其它试验点;海拔80 m的源头弄试验点叶片最宽2.9 cm,极显著大于其它试验点。
长喙毛茛泽泻根长6.8 ~ 8.8 cm,不同海拔根系长度存在极显著差异,海拔630 m的九户垄试验点最短,显著小于其它试验点。纤匍茎节间长5.7 ~ 8.8 cm,海拔300 m的大贯试验点最短,显著小于其它试验点。

表3 不同海拔长喙毛茛泽泻的形态特征Table 3 Morphological properties of R. rostratum at different altitude
2.3 不同海拔对长喙毛茛泽泻生物量的影响
不同海拔试验地单株地上部分叶与地下部分根的生物量及其占比如表4。

表4 不同生境长喙毛茛泽泻单株生物量分配情况Table 3 Distribution of individual biomass of R. rostratum at different altitude
从表4中可以看出,长喙毛茛泽泻单株总生物量0.18 ~ 0.25 g,其中,地上部分0.11 ~ 0.12 g,占比46.83%~ 69.01%;地下部分0.06 ~ 0.13 g,占比30.99% ~ 53.17%。随着海拔的增高,总生物量有增大趋势,但不存在显著差异。其中,地上部分生物量变化不大,占比减少;地下部分生物量增大,占比增加,不同种植地点存在显著差异,海拔最低的源头弄显著小于其它种植点。
3 小结与讨论
海拔高度对长喙毛茛泽泻的营养生长及物质积累有一定的影响。长喙毛茛泽泻经过7个月的生长,平均株高7.1 ~ 13.5 cm,地茎0.38 ~ 0.52 cm,叶片长2.1 ~ 3.3 cm,叶片宽1.7 ~ 2.9 cm,根长6.8 ~ 8.8 cm,纤匍茎节间长5.7 ~ 8.8 cm,不同试验点间存在极显著差异,说明不同的海拔高度对其生长有较大的影响。单株总生物量0.18 ~ 0.25 g,随着种植点海拔增高,单株总生物量明显增大,地下部分占比明显增加。
长喙毛茛泽泻在丽水市莲都林场不同海拔生境种植均能成活,至冬季生长繁殖停止时繁殖系数达到2.75 ~39.80,其中以海拔840 m的圩地种植点繁殖系数最高,其它海拔高度的试验点差异不大,这也许是与圩地种植点的水源是山泉水有关,关于水质状况对长喙毛茛泽泻生长及种群繁殖的影响有待今后做深入研究[9-11]。
试种地的生态环境是影响长喙毛茛泽泻无性繁殖能力的重要因素之一[9]。长喙毛茛泽泻在南明山原生境地海拔100 m左右,海拔80 m的源头弄生境与其相类似,但由于样地在小谷小淤,常受山洪冲刷,造成泽泻被冲毁,影响了其生长和繁殖。海拔300 m的大贯,由于经常有牛羊等动物的踩踏,影响了泽泻的生长与繁殖。海拔630 m的九户垄,也许由于水质较差及田螺侵害,影响了植株生长及其无性繁殖水平。海拔1 040 m的葑垟湖由于水位过低,经常干燥,影响了长喙毛茛泽泻生长与繁殖[13]。与其他海拔点相比,海拔840 m的圩地种植点的繁殖系数最高,虽然株高最高,但地径最细,导致与其它海拔点的植株生物量之间没有差异。总体上看,长喙毛茛泽泻在其原生地丽水种植是可行的,选择合适的生境更有利于提高种群的扩张水平。
[1] 孙祥钟. 中国植物志(第八卷)[M]. 北京:科学出版社,1992:136
[2] 国务院. 国家重点保护野生植物名录(第一批)[J]. 植物杂志,1999,(5):4-11.
[3] 陈中义,何国庆,陈家宽. 濒危植物长喙毛茛泽泻生物学特性观察[J]. 武汉大学学报(自然科学版),1997,(2):66-69.
[4] 王建波,陈家宽,利容千,等. 长喙毛茛泽泻的胚胎发生[J]. 武汉大学学报(自然科学版),1996,(2):195-201.
[5] 田惠桥,陈家宽,郭友好. 濒危植物—长喙毛茛泽泻的雌雄配子体发育[J]. 植物分类学报,1995,(3):221-224,317-318.
[6] 王建波,陈家宽,利容千. 长喙毛茛泽泻和冠果草种子贮藏及萌发特性(简报)[J]. 植物生理学通讯,1997,(2):119-122.
[7] 胡小辉,陈家宽,王建波,等. 长喙毛茛泽泻遗传多样性及其与繁育系统的关系[J]. 云南植物研究,1999,(2):104-110.
[8] 王建波,陈家宽,利容千,等. 长喙毛茛泽泻的大小孢子发生及雌雄配子体发育[J]. 武汉大学学报(自然科学版),1994,(4):95-101.
[9] 王建波,陈家宽,利容千,等. 长喙毛茛泽泻的生活史特征及濒危机制[J]. 生物多样性,1998,(3):7-11.
[10] 陈中义,陈家宽. 长喙毛茛泽泻的种群分布格局和群落内种间关联[J]. 植物生态学报,1999,(1):57-62.
[11] 陈中义,陈家宽. 长喙毛茛泽泻致濒机制和保护措施[J]. 湖北农学院学报,1997,(1):30-34.
[12] 李今,陈家宽,周进,等. 迁地保护中普通野生稻和长喙毛茛泽泻种群动态及评价[J]. 植物生态学报,1999,(3):84-91.
[13] 陈锦华,姜益泉,戴余军,等. 长喙毛茛泽泻的研究进展[J]. 孝感学院学报,2008,28(6):5-10
Experiment on Plantation of Ranalisma rostratum at Different Altitudes in Lishui
LU Li-ping1,JI Zhi-li1,WANG Wu-jun2,DU You-xin3,HE Xiao-yong3
(1. Liandu Forest Farm of Zhejiang, Lishui 323000, China; 2. Liandu Forestry Extension Station, Lishui 323000, China; 3. Lishui Institute of Forestry, Lishui 323000, China)
Experiments were conducted on planting of endangered Ranalisma rostratum in May of 2016 at different altitude in Liandu Forest Farm of Lishui, Zhejiang province. Observations and determinations were implemented from July to November of the current year on variation of plantings and reproduction coefficient. The results showed that the survival rate of R. rostratum reached 100% at different altitude from 80 m to 1040 m with the mean reproduction coefficients ranged from 2.75 to 39.80, the highest was at 840 m. It h ad significant differences of mean height, ground diameter, leaf size, root and stolon length among different altitudes. Determination on individual biomass November demonstrated it ranged from 0.18 g to 0.25 g, increasing with altitude, and the proportion of underground biomass increased more significantly than aboveground ones.
Q948.8
:A
:1001-3776(2017)03-0068-04
10.3969/j.issn.1001-3776.2017.03.013
2017-01-08 ;
2017-04-25
丽水市莲都区科技项目(“不同海拔对濒危植物长喙毛茛泽泻种群形态特征属性指标的影响”201609)
陆丽萍,工程师,从事森林培育工作;E-mail:2532346605@qq.com。通信作者:何小勇,教授级高工,从事森林培育研究;E-mail:2532346605@qq.com。
