3种安息香属植物的根尖染色体核型分析
2017-08-09石从广高燕会柳新红李因刚杨少宗蒋冬月朱光权
石从广,高燕会,柳新红,李因刚,杨少宗,蒋冬月,朱光权
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江农林大学,浙江 临安 311300)
3种安息香属植物的根尖染色体核型分析
石从广1,高燕会2,柳新红1,李因刚1,杨少宗1,蒋冬月1,朱光权1
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江农林大学,浙江 临安 311300)
2013年9月和10月,在浙江省杭州市采集芬芳安息香Styrax odoratissimus,越南安息香S. tonkinensis和赛山梅S. confusus种子,砂藏至翌年发芽,待根长至3 ~ 5 cm时,对3种安息香属植物的根尖进行染色体制片和核型分析。结果显示,3树种均为二倍体,体细胞染色体数均为2n=2x=16。芬芳安息香核型公式为2n=16=10 m +6 sm(2 sat),核不对称系数为58.20%,染色体核型为2A型;越南安息香核型公式为2n=16=12 m(1 sat)+4 sm (2 sat),核不对称系数为53.27% ,染色体核型为1A型;赛山梅染色体核型公式为2n=16=12 m(1sat)+4 sm,核不对称系数为56.54%,染色体核型为1A型。表明越南安息香和赛山梅核型较为类似,亲缘关系较近,与芬芳安息香之间差别较大,芬芳安息香比越南安息香和赛山梅进化程度较高。3种安息香属植物染色体核型均为A型,表明其进化地位较低,植物较为原始。
安息香属;染色体;核型分析;二倍体;进化
安息香科 Styracaceae 安息香属 Styrax 植物分布于亚洲东部、美洲北部和西部及地中海地区,在我国主产于长江流域以南各省区[1-2]。浙江有12种和1变种,其中的越南安息香 Styrax tonkinensis又叫白花树,落叶乔木,浙江省泰顺县有分布,其树脂称“安息香”,为贵重药材,种子油可供药用,也是制生物柴油的优良原料。越南安息香是本属中研究较多的树种,研究内容主要包括形态和物候以及大小孢子(配子体)等生物学特征研究[3-5];地理分布、引种栽培、气候群落特征、以及天然群体遗传多样性研究[6-9];种仁和种子油的营养成分分析[10-11]、苗木繁育[12],定向抚育栽培[13]和种子成熟期脂肪酸变化规律[14]、种子油脂毒理学初步评价[15]以及利用种子油制备生物柴油等[16-17];木材加工利用及园林应用等[18-19]。该属的芬芳安息香S. o doratissima和赛山梅S. confusus,都是浙江较为常见的落叶灌木或小乔木,目前只有利用催化剂对提取的赛山梅种子油催化制备生物柴油的报道[20]。
通过对越南安息香、芬芳安息香和赛山梅的根尖进行染色体制片和核型分析,研究这3个种种间的遗传变异和亲缘关系,为杂交育种以及杂种后代鉴定等提供理论依据。
1 材料和方法
1.1 试验材料与处理
试验材料为芬芳安息香、越南安息香和赛山梅的种子。其中芬芳安息香和赛山梅的种子采自浙江省杭州市西湖区午潮山,野生种;越南安息香种子采自杭州市西湖区浙江省林业科学研究院苗圃地,是从江西省吉安市吉水县引种栽培的第二代种子。
芬芳安息香在2013年9月下旬种子成熟后采种,赛山梅和越南安息香在2013年10月上旬采种,采下的果实经风干开裂后脱粒去果壳。2013年12月,将3种种子分别沙藏至第二年春天发芽,待根长至3 ~ 5 cm时,于8:30-9:30,每种各取25 ~ 30株芽苗,用解剖刀截取长约0.5 c m的根尖,放入饱和对二氯苯中,28℃处理2.5 ~ 3.0 h,水洗并经无水乙醇:冰乙酸(3:1)固定24 h,保存于70%酒精中于4℃冰箱中保存备用。将固定好的根尖于1 mol·L-1HCI,65℃水浴解离10 min,在载玻片上染色体制片,在莱卡显微镜观下观察并拍照。
1.2 染色体计数和核型分析
取染色体分散良好、着丝点清晰的中期分裂相进行染色体计数,结合图形分析处理软件Photoshop CS3进行处理计数,测量染色体长臂及短臂长度,再用Excel计算染色体相对长度、相对长度系数、臂比、核型不对称系数,计算思路与公式如下。
1.2.1 染色体计数 (1)每种统计50个以上可准确计数染色体的细胞,以确认其染色体数目;(2)分别从5个以上根尖压片中,选择次缢痕清晰而又分散较好的5 ~ 12个细胞供核型分析;(3)编号并进行测量,随体(sat)长度不计算在总长度内;(4)根据测量所得数据,进行同源染色体配对,并计算其平均值,按染色体的总长度从长至短顺序编号后取5 ~ 12个细胞的平均值,作为该物种的染色体参数;(5)染色体长度参数,按国际惯例,均示相对长度(RL)。

1.2.2 核型分析 按照Levan等[21]和李懋学等[22]的报道,以及Stebbins[23]的核型分类标准进行分析。根据染色体臂比来确定着丝粒的位置,得出核型公式(表1);按核型中最长染色体与最短染色体之比及臂比大于2的染色体所占的比例来确定核型类型;按照Arano[24]的方法计算核型不对称系数,其比值越大表示核型越不对称。
2 结果与分析
2.1 芬芳安息香核型分析

表1 染色体臂比对应的类型和形态特征Table 1 Arm ratio and their karyogram
从表2,表3,图1至图3可知,芬芳安息香的根尖细胞在有丝分裂中前期有8对计16条染色体,根尖体细胞染色体数2n = 2x = 16,表明为二倍体,染色体基数x=8;体细胞染色体核型公式:2n = 16 = 10 m+6 sm(2 sat),其中第1,2,5对染色体为sm,其余为m,第1和第5对染色体的短臂上各带有1对sat。染色体相对长度范围为9.75 ~ 23.41,AAR 1.61,L/S大于2的染色体有2对,比率为25%,Lt/St染色体的比值为2.40,ASI为58.20%,染色体IRL公式为1L+2M2+3M1+2S,核型为2A型。3种安息香属植物染色体参数和核型特征见表2和表3;中期染色体、核型图及核型模式图见图1至图3。
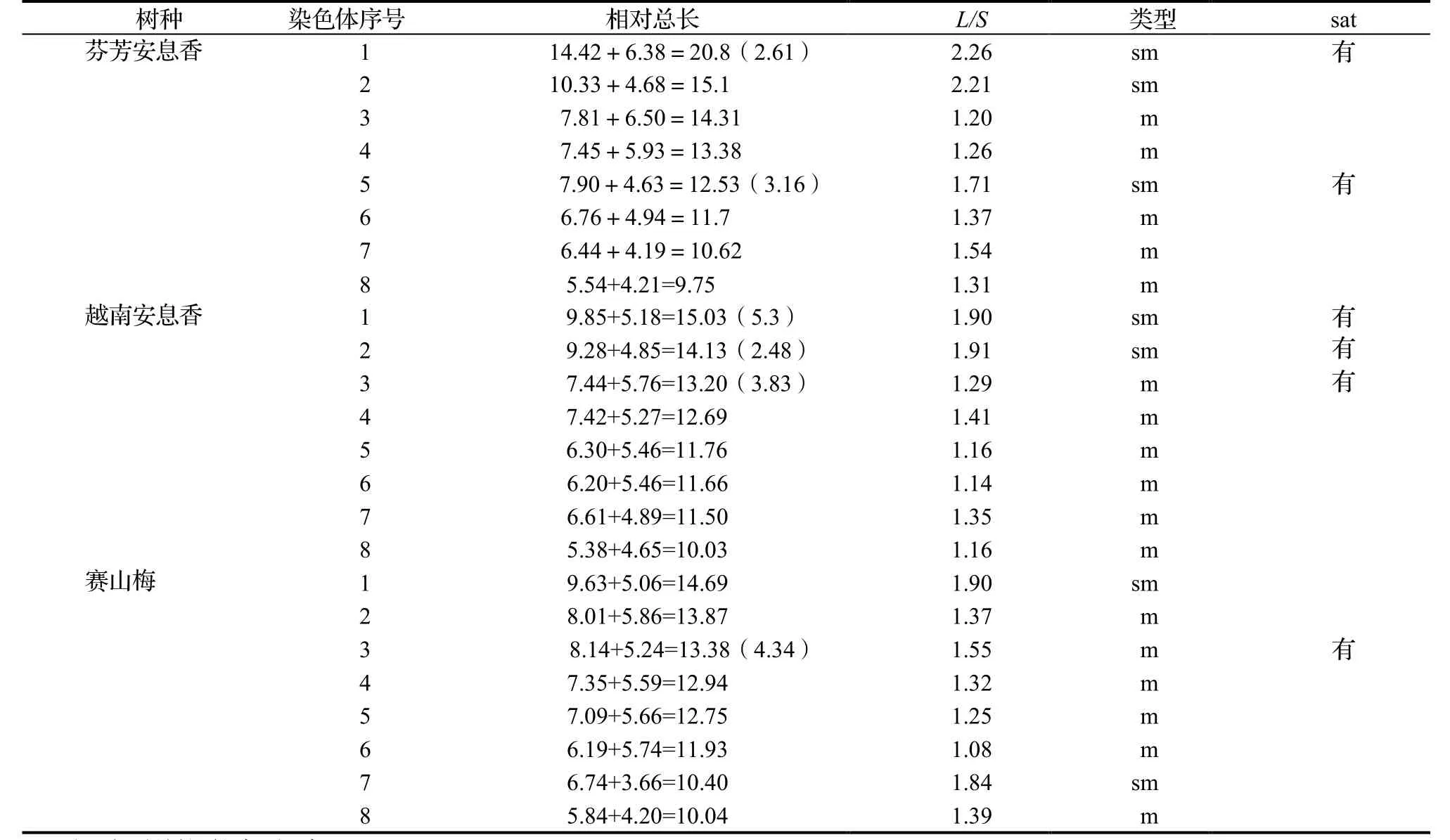
表2 3种安息香属植物的染色体组型分析Table 2 Karyotype analysis of 3 species of Styrax
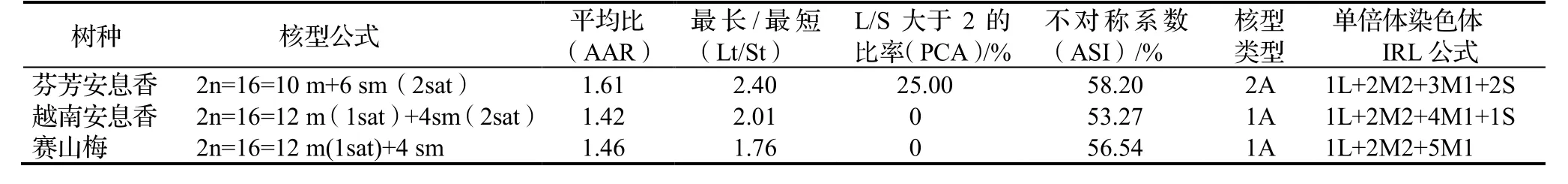
表3 3种安息香属植物核型参数Table 3 Karyotype parameters of 3 species of Styrax

图 1 3种安息香属植物根尖细胞有丝分裂中前期染色体图Figure 1 Metaphase chromosome of root tip cells from 3 species of Styrax

图 2 3种安息香属植物核型模式图Figure 2 Idiogram of 3 species of Styrax
2.2 越南安息香核型分析
从表2,表3,图1至图3可知,越南安息香的根尖细胞在有丝分裂中前期有8对计16条染色体,根尖体细胞染色体数 2n = 2x = 16,表明为二倍体,染色体基数x=8;体细胞染色体核型 公式为2n = 16 = 12 m(1sat)+4 sm (2 sat),其中第1和第2对染色体为sm,其余为m,第1,第2,第3对染色体的短臂上各带有1对sat,L/S最高为1.91。染色体相对长度范围为10.03 ~ 20.33,AAR 1.42,没有臂比大于2的染色体,Ls/St 为2.01,ASI为53.27%,染色体IRL组成为1L+2M2+4M1+1S,核型为1A 型。
2.3 赛山梅核型分析
从表2,表3,图1至图3可知,赛山梅的根尖细胞在有丝分裂中前期有8对计16条染色体,根尖体细胞染色体数2n = 2x = 16,表明为二倍体,染色体基数x = 8;体细胞染色体核型公式为2n = 16 =12 m(1 sat)+4 sm。赛山梅第1和第7对染色体为sm,其余为m,第3对染色体的短臂上各带有1对sat,L/S最高为1.90。染色体相对长度范围为10.04 ~ 14.69,AAR 1.46,没有臂比大于2的染色体,Ls/St为1.76,ASI为56.54%,染色体IRL组成为1L+2M2+5M1,核型为1A型。
3 结论与讨论

图 3 3种安息香属植物染色体核型图Figure 3 Chromosome karyotype of 3 species of Styrax
研究观察到的3种安息香属植物均为二倍体,且体细胞染色体数均为2n = 2x = 16,染色体多数为m,少数为sm,没有st和t。
根据物种进化的趋势,从染色体数目看,在多倍体植物中,一般是从2倍体向多倍体演化,从染色体核型看,是从对称性向非对称性演化。Stebbins按对称性程度的高低将染色体核型分为12种类型(1A,2A,3A,4A;1B,2B,3B,4B;1C,2C,3C,4C),认为核型对称性程度越高的生物,其染色体变异越小,进化程度也越低,对称性程度越低的生物,其染色体变异越大,进化程度越高[23]。根据染色体核型图(图3)和组成公式(表3)可以看出越南安息香与赛山梅核型相近,均为1A型,前者比后者多了两对随体,而与芬芳安息香核型(2A型)差异较大,说明越南安息香与赛山梅亲缘关系较近,而与芬芳安息香亲缘关系较远,芬芳安息香比越南安息香和赛山梅的进化程度要高。但三个种的核型均为A型,表明其进化程度总体上都较低。
安息香属植物染色体数目已知的有n=8和n=20(野茉莉 Styrax japonicus),但本研究中亦发现有极少的n=16的现象(结果未在本论文中列出),出现此现象的原因可能是细胞在有丝分裂时出错造成。从现有资料看,多数安息香属植物体细胞染色体数都为2n = 2x =16,但核型差异明显,这些差异可能造就了安息香属植物的遗传多样性。
[1] 张程. 中国安息香科植物种质资源及研究进展[J]. 江西林业科技,2010年,(6):42-47.
[2] 中国科学院植物研究所.中国高等植物图鉴第三册[M]. 北京:科学出版社,1985,338.
[3] 吴君,李因刚,罗修宝,等. 白花树种子生物学特性[J]. 浙江农林大学学报,2014,31(1):9-13.
[4] 吴君,李因刚,柳新红,等. 越南安息香大小孢子发生和雌雄配子体发育[J]. 浙江林业科技,2014,34(1):47-51.
[5] 吴君,李因刚,柳新红,等. 白花树花粉生活力检测方法与贮藏特性研究[J]. 林业科学研究,2014,27(1):017-023.
[6] 李因刚,柳新红,赵勋,等. 我国越南安息香的地理分布及其气候特征[J]. 西南林业大学学报,2011,31(1):5-10.
[7] 陈涛,陈忠毅. 安息香科植物地理分布研究[J]. 植物研究,1996,35(1):97-99.
[8] 柳新红,李因刚,赵勋,等. 白花树天然群体表型多样性研究[J]. 林业科学研究2011,24(6):694-700.
[9] 谢建秋,柳新红,王军峰,等.东京野茉莉引种试验初报[J]. 浙江林业科技,2006,26(4):33-35.
[10] 顾雁蕾,叶兴乾,柳新红.野茉莉种仁与种子油的营养成分及理化特性研究[J]. 食品工业科技,2010,12(31):328-329.
[11] 肖复明,曾志光,杨桦,等. 东京野茉莉种子油营养成分研究[J]. 天然产物研究与开发,2005,17(3):344-345.
[12] 张亮亮,柳新红,林新春. 白花树组织培养技术研究[J]. 浙江林业科技,2013,33(3):16-19.
[13] 石从广,朱光权,柳新红,等. 施肥对白花树各生长性状的影响[J]. 浙江林业科技,2013,33(3):9-15.
[14] 石从广,李因刚,朱光权,等. 白花树种子成熟期含油率和脂肪酸变化规律探讨[J]. 浙江农林大学学报,2013,30(3):372-378.
[15] 石从广,朱光权,黄海志,等. 白花树种子油精炼及其毒理学初步评价[J]. 中国食品学报,2014,14(5):194-201.
[16] 刘光斌,黄长干,刘苑秋,等. 东京野茉莉油的提取及其制备生物柴油的初步研究[J]. 江西农业大学学报,2007,29(4):685-689.
[17] 陈洁,聂小安,刘振兴,等. 环氧白花果油基脂肪酸甲酯的无羧酸法制备工艺[J]. 中国油脂,2014(7):69-72.
[18] 鲍甫成,江泽慧. 短周期工业用材林木材性质研究[J]. 世界林业研究,1995,(8),50-56.
[19] 骆显春,杨桦,曾志光,等. 东京野茉莉木材性质分析与利用[J]. 江西农业大学学报,2007,29(1):77-80.
[20] LIU Z X,NIE X A,WANG Y G. Biodiesel Preparation from Styrax confusus Hemsl. Oil Catalyzed by Magnetic Catalyst S2O82--ZrO2/Fe3O4[C],International Conference on Advances in Energy & Environment Science,2013,805-806,247-250.
[21] Levan A,Fredga K,Sandberg A A. Nomenclature for centromerie position on chromosome [J]. Hereditas,1964,52:201-220.
[22] 李懋学,陈瑞阳.关于植物核型分析的标准化问题[J]. 武汉植物学研究,1985,3(4):297-302.
[23] Stebbins G L. Chromosome evolution in higher plants [M]. London:Edward Arnold Ltd,1971:87-123.
[24] Arano H. Cytological studies in subfamily Carduoideae (Composite) of Japan IX[J]. Bot Mag Tokyo,1963,76:419-427.
Karyotype Analysis of Root Chromosome in Three Species of Styrax
SHI Cong-guang1,GAO Yan-hui2,LIU Xin-hong1,LI Yin-gang1,YANG Shao-zong1,JIANG Dong-yue1,ZHU Guang-quan1
(1. Zhejiang Academy of Forestry, Hangzhou 310023, China; 2. Zhejiang A & F University, Lin’an 311300, China)
Seeds of Styrax odoratissimus, S. tonkinensis and S. confusus were collected on September and October of 2013 in Hangzhou, Zhejiang province. They were treated by sand storage until the next spring. When seeds germinated and root had 3-5 cm, the apices of three species were used for chromosome sectioning and karyotype analysis. The results showed that the tested species were diploid, and their genome chromosome number was 2n=2x=16. Karyogram of S. o doratissimus was 2n=16=10 m+6 sm (sat), n uclear as ymmetry i ndex was 58.20%, and karyotype was 2A. Karyogram of S. tonkinensis was 2n=16=12m(1sat)+4sm(2sat), nuclear asymmetry index was 53.09%, andt karyotype was 1A. Karyogram of S. confusus was 2n=16=12 m(1sat)1+4 sm, nuclear asymmetry index was 56.54%, and the karyotype was 1A. The test concluded that karyotypes of S. tonkinensis and S. confusus were similar, but different with that of S. odoratissimus, indicating that the chromosome evolution of three species was low, and they were primitive.
Styrax; chromosome; karyotype analysis; diploid; evolution
S722.3
:A
:1001-3776(2017)03-0023-05
10.3969/j.issn.1001-3776.2017.03.004
2016-11-16;
2017-02-21
本稿件受浙江省农业公益性研究项目“安息香属优质能源树种选择与开发利用”资助(2012C22097)
石从广,博士,副研究员,从事林木遗传改良研究;E-mai:363928874@qq.com。通信作者:朱光权,教授级高工,从事植物分类学研究;E-mail:2554396001@qq.com。
