木荷种子园无性系开花物候及同步性分析
2017-08-07杨汉波周志春
杨汉波,张 蕊,宋 平,周志春
(中国林业科学研究院亚热带林业研究所,浙江省林木育种技术研究重点实验室,浙江 杭州 311400)
木荷种子园无性系开花物候及同步性分析
杨汉波,张 蕊,宋 平,周志春*
(中国林业科学研究院亚热带林业研究所,浙江省林木育种技术研究重点实验室,浙江 杭州 311400)
[目的]研究种子园无性系的花期物候特征和花期同步性及影响因子对种子园科学管理具有指导意义。[方法]2015年和2016年连续2 a对木荷无性系种子园中19个无性系进行花期物候调查,分析其开花物候与花期同步指数。[结果]木荷种子园各无性系开花持续时间为10~28 d,2016年较2015年晚2 d进入始花期。相关分析结果表明,开花数多的无性系比开花数少的无性系花期更长、座果数更多,花期持续时间长的无性系比持续时间短的无性系座果数更多。无性系组合间花期同步指数有较大差异,2015年和2016年变化范围分别为0.552~0.857和0.406~0.808,平均分别为0.758和0.713,其中花期同步指数大于0.80的无性系分别在50%和40%以上。年度内花期同步指数变异系数分别为12.016%~46.476%和15.375%~51.202%,无性系之间平均花期同步指数存在极显著变异(P<0.01)。年度间花期平均同步指数为0.737,平均花期同步指数间不存在显著变异(P>0.05)。年度间花期同步指数相关系数为0.229,表明年度间花期同步指数具有一定的相关性,但不显著。[结论]木荷种子园无性系年度间花期同步指数较为稳定,可根据开花物候和花期同步指数对无性系进行筛选和优化,同时辅以人工授粉等措施弥补花期同步性差异,以达到提高种子园种子产量和质量的目的。
木荷;种子园;开花物候;花期同步性;稳定性
林木种子园是以生产遗传品质较高的林木种子为目的的生产基地,主要分为无性系种子园和实生种子园,其中无性系种子园以能保持优树的优良品质、来源清楚以及开花结实较早等优点而被广泛应用[1-2]。开花物候是植物重要的生活史特征之一[3],同时还是一个重要的适合度因子(植物或植物群体对环境适应性的影响因子),对生殖成功有重要的影响[4-5]。种子园内亲本花期同步性是影响种子园产量和质量的关键因素,亲本花期不同步,会限制亲本间基因的充分交流,降低种子遗传品质,减少种子园产量[6-7]。估算无性系花期同步指数可以评价种子园无性系之间有效异交水平和遗传多样性状况,对于指导种子园控制授粉和辅助授粉工作及去劣疏伐有积极的指导意义和重要的参考价值[7-9]。目前,国内外许多建园树种,如马尾松(PinusmassonianaLamb.)[2]、油松(P.tabulaeformisCarrière)[10]、北美红栎(QuercusrubraLine.)[11]、辐射松(P.radiateD. Don)[12]、柚木(TectonagrandisLinn. f)[13]和杉木(Cunnighamialanceolata(Lamb.) Hook.)[14]等都开展了这方面的研究,并为种子园经营管理提供了科学依据。但这些研究多是针对风媒传粉树种,而对雌雄同花虫媒传粉的阔叶树种子园较少报道。
木荷(SchimasuperbaGardn. et Champ.)属山茶科(Theaceae)木荷属(SchimaReinw.)常绿阔叶大乔木,花白色,常多朵排成总状花序,雌雄同花,虫媒传粉[15]。木荷叶革质,含水量大,材性优良,抗逆性强,是我国南方各省区的珍贵优质阔叶用材和高效生物防火树种。但有关木荷育种的研究起步较晚,涉及到种子园营建及花期同步性等方面的研究较少,目前仅辛娜娜等[16]对6个不同产地木荷优树无性系花期物候进行了分析。本研究在木荷种子园无性系连续2 a开花结实调查的基础上,进一步研究各无性系开花性状的遗传差异,分析各无性系间花期同步指数及年度稳定性,以期为木荷种子园丰产经营提供科学依据。
1 材料与方法
1.1 试验材料
研究材料来源于浙江省金华市兰溪苗圃木荷一代无性系种子园(29°08′16″ N,119°27′47″ E,平均海拔高度42 m,年均温度17.7 ℃,年均降水量1 438.9 mm)。种子园面积20 hm2,共142个亲本无性系,分为17个生产小区,小区内无性系配置以随机与调整相结合定植,株行距5 m×5 m,2013年进入正常开花结实期。研究对象为种子园第4和6小区内进行,其中第4小区调查对象为该小区内正常开花结实且均匀分布的9个无性系(11、12、13、15、17、21、24、26和27号),原产地均为福建省建瓯市(27°02′ N,118°19′ E,海拔高度100~300 m);第6小区调查对象为该小区内正常开花结实且均匀分布的10个无性系(5、29、33、43、60、61、64、70、72和103号),其中5号原产地为浙江省龙泉市(28°05′ N,119°08′ E,海拔高度100~300 m),29、33和43号原产地为福建省建瓯市(27°02′ N,118°19′ E,海拔高度100~300 m),60、61和64号原产地为福建省三明市(26°17′N,118°19′ E,海拔高度134 m),70和72号原产地为福建省南平市(27°43′ N,119°03′ E,海拔高度673 m),103号原产地为江西省鹰潭市(28°21′ N,116°81′ E,海拔高度44 m)。
1.2 试验方法
花期观测和花量调查采用固定标准株法,每个无性系选择生长中庸的5棵标准分株进行一级侧枝数(主干上着生的侧枝数)和花量的调查及花期观测。统计方法如下:在树冠中部东、南、西、北4个方向上各选择一个生长正常的一级侧枝,每2 d分别统计其上的花朵数并计算一级侧枝的平均花朵数量,一级侧枝数与平均花朵数量的乘积即为整个植株的花朵数量[16]。花期各时段的确定标准为[16-17]:每株有15%花朵开放的日期为始花期,每株有45%花朵开放的日期为盛花期,每株有90%的花朵开放的日期为末花期。
2014—2016年兰溪市木荷种子园的温度和降雨量信息由兰溪市气象局提供。
1.3 数据分析
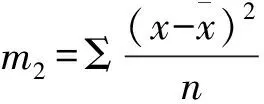
无性系之间花期同步指数(C)采用Gunaga等[6]提出的方法计算:
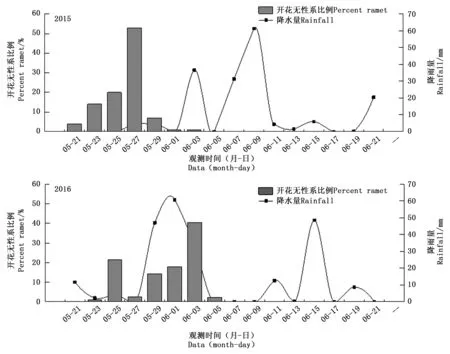
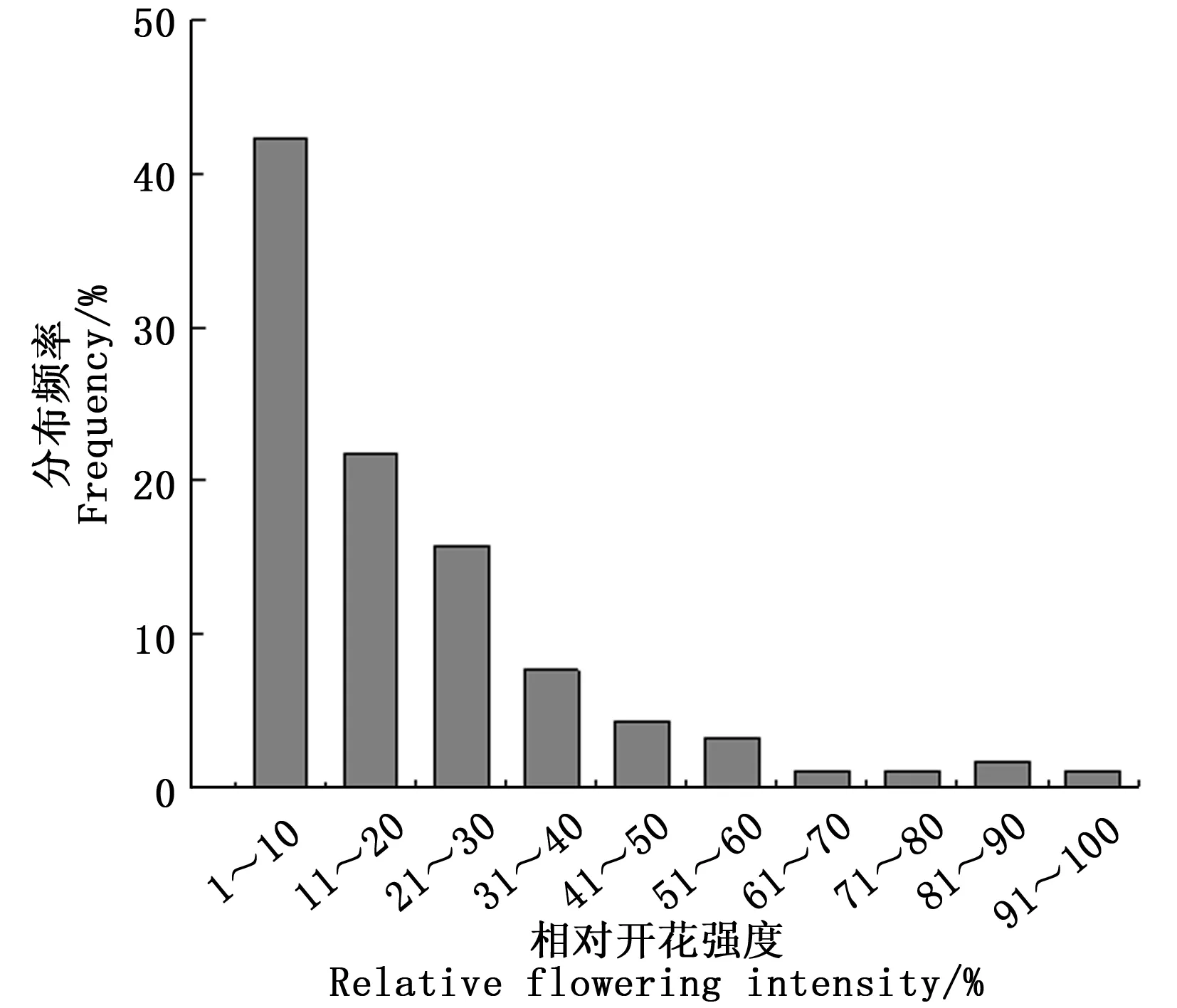
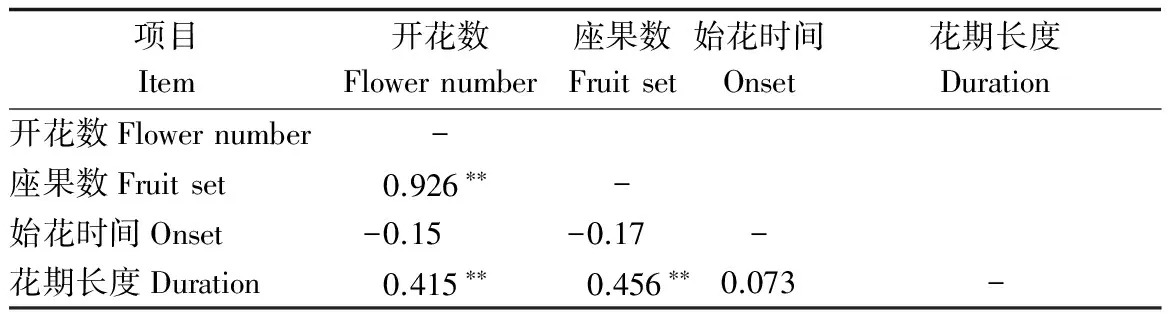
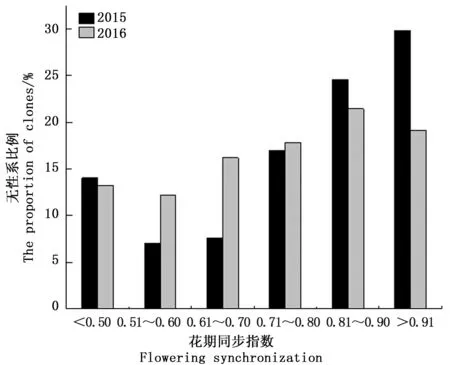
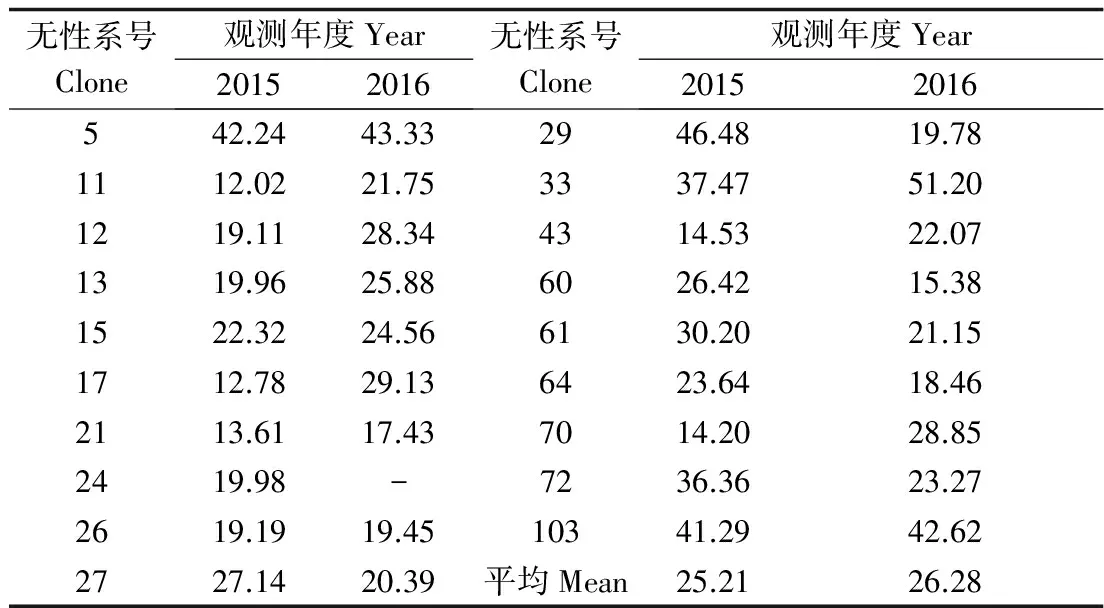
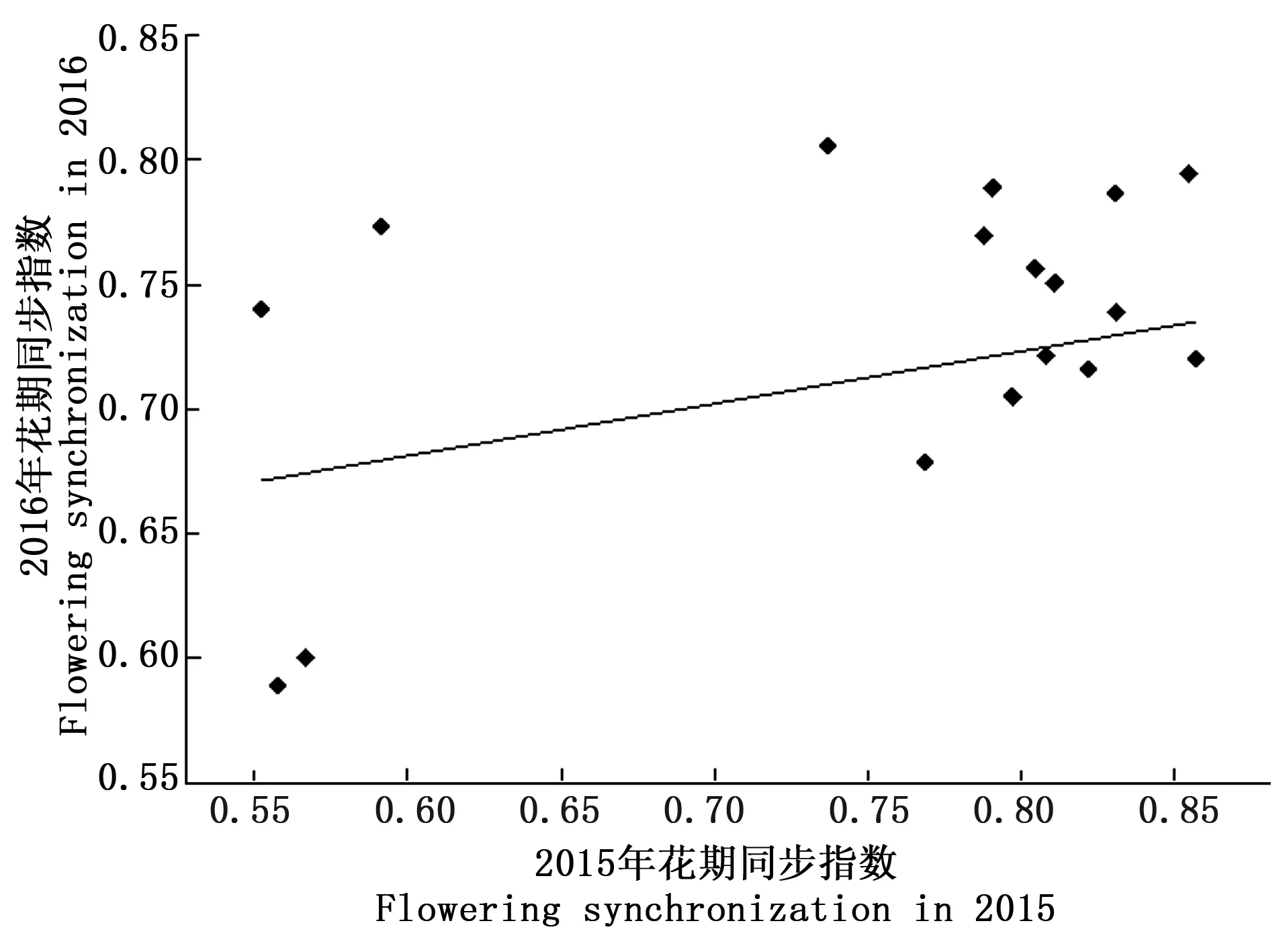
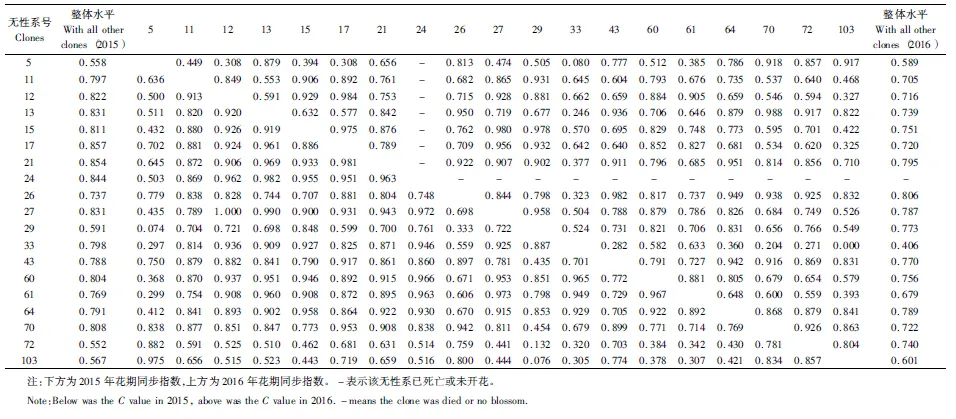
式中:Pij为第j个无性系在第i天进入盛花期植株的比例,Pik为第k个无性系在第i天进入盛花期的植株比例,n为花期观测天数,Pij×Pik为无性系j与k的开花联合概率。当两个无性系的花期完全重叠时,C=1,不重叠时C=0,部分重叠时0 以年度为重复,对各无性系与其它无性系的平均花期同步指数的差异显著性作单因素方差分析。以年度内各无性系的花期同步指数为统计值做年度间相关分析。用Pearson相关系数进行无性系的始花时间、花期持续时间、开花数及座果数之间的相关分析。统计分析借助SPSS20.0软件进行。 2.1 无性系花期观测 木荷单花一般在清晨开放,依其形态和散粉特征可分为3个时期:(1)花瓣松动至花药完全开裂;(2)柱头伸长,顶端乳突状突起明显,不断分泌粘液;(3)花药全部开裂或枯死,柱头变褐或枯黄,花瓣卷缩枯萎。通过对各无性系花期分析(图1)可以看出,无性系开花持续时间为10~28d,2015年内,15、33和60号无性系开花最早,并最早达到开花盛期,33、61和64号最早进入开花末期;2016年内,60号开花最早,较2015年晚2d,并最早达到盛花期,33号最早进入末花期。年内和年间各无性系的始花期、盛花期和末花期的日期及各时段持续天数存在较大的差异。例如,2015年内64与72、103号无性系盛花期仅有2 d重叠。在整体水平上,2016年种子园各无性系的始花期、盛花期和末花期较2015年存在较为明显的延迟现象。进一步分析花期物候进程发现,2014年积温(6 202.2 ℃)略高于2015 年(6 162.6 ℃)。2015年仅存在一个开花高峰时段(5月27日),降雨量仅4.4 mm,而2016年存在两个开花高峰时段(5月25日和6月3日),其中第1个开花高峰时段降雨量与2015年基本相同,为4.1 mm,而第2个开花高峰时间降雨较多,为33.2 mm(图2)。种子园内无性系分株开花强度的主要分布频度在1.0%~30.0%之间(图3),其分布频度的偏斜率为1.88;其中以10.0%左右的开花强度分布频度最高,达到总频度的42.4%。 图1 木荷种子园无性系花期观测结果(月-日)Fig.1 Observation result of flowering period of S. superba clones (month-day) 图2 木荷种子园无性系在各时段开花比例与降雨量Fig.2 The proportion of clones from S. superba seed orchard and rainfall pattern in the flowering phenology 图3 木荷种子园无性系个体相对开花强度的分布Fig.3 Frequency distributions of relative flowering intensity in S. superba seed orchard individuals 2.2 开花指数与生殖成功的相关分析 木荷种子园无性系的始花时间、花期长度、开花数、座果数之间的相关分析结果显示(表1),始花时间与开花数和座果数存在负相关关系,而与花期长度存在正相关关系。花期长度与开花数和座果数均成极显著正相关关系,开花数与座果数间也存在极显著的正相关关系。表明开花数多的无性系比开花数少的无性系花期更长、座果数更多,花期持续时间长的无性系比花期持续时间短的无性系座果数更多。开花数目多、花期长均有利于提高植株的座果率。 表1 种子园无性系花量、始花时间、花期长度及座果率的相关分析 注:**相关性极显著(双尾检验)。 Note: **Correlation is significant at 0.01 level (2-tailed). 2.3 无性系组合间花期同步指数变化 从图4可以看出,木荷种子园内约75%的无性系之间的花期同步指数大于0.6,表明这些无性系之间的花期同步性较高,但仍有约14%(2015年)和13%(2016年)的无性系间的同步指数低于0.5,这不利于木荷种子园内无性系之间充分的随机交配。花期同步指数在无性系组合间有明显的差异(表2),如以2015年的观察结果为例,17、24号与其它无性系花期同步指数均较高,整体水平分别为0.857和0.854,而5、72号与其它无性系花期同步指数均较低,分别为0.558和0.552。同一无性系与其它无性系的花期同步指数也有较大差异,如17号无性系与21号花期同步指数达到0.981,但与29号的同步性则为0.599,而11号无性系与其它无性系的同步指数变化则相对较小。 2.4 年度内和年度间花期同步指数变化 2015年度内木荷种子园各无性系的花期同步指数主要集中在大于0.8的范围内,大于0.9的无性系相对较多;而2016年度内各无性系在大于0.6的各个范围内分布较为均匀,0.8~0.9范围内分布的无性系则相对较多(图4)。2015年和2016年平均花期同步指数变动系数分别为12.02%~46.48%和15.38%~51.20%,平均为25.21%和26.28%(表3)。方差分析表明,2015年(F=5.842,P<0.01)和2016年(F=4.818,P<0.01)年度内无性系的平均花期同步指数均存在极显著变异。 图4 花期同步指数范围内无性系所占比例Fig.4 The proportion of clones into class of flowering synchronization 从整体来看,木荷种子园内无性系年度间花期平均同步指数变化范围为0.406~0.857,平均为0.737。方差分析结果表明,年度间花期同步指数间不存在显著变异(F=1.387,P=0.247>0.05),2015年与2016年的花期同步指数呈正相关(r=0.229,P=0.361>0.05),但未达到显著水平(图5),说明木荷种子园无性系的花期同步指数具有一定的相对稳定性。 表3 木荷种子园无性系的平均花期同步指数变异系数 图5 2015年与2016年年度间花期同步指数相关分析Fig.5 Analysis of correlation of flowering synchronization between 2015 and 2016 表2 木荷种子园各无性系之间的花期同步指数 3.1 无性系开花物候与生殖特征 植物开花物候受许多因素的影响,研究表明,开花物候不仅与植物类群的系统发生及遗传特性有关[20],还与环境条件(如纬度、海拔高度、湿度和温度等)有密切的联系[21]。本研究中,木荷种子园无性系与保存在浙江省龙泉市的优树无性系的开花持续时间基本一致,但种子园无性系的始花时间早于浙江龙泉的同一无性系[16],这可能与两地不同的土壤环境、海拔高度和降雨量等气候条件有关。木荷种子园无性系年度间开花物候表现出较为明显的差异性,2016年始花期、盛花期及末花期均较2015年延迟,这与上一年的年积温有关。2015年仅有一个开花高峰期,表现出一种集中开花模式[18],而2016年具有两个开花高峰期,这应与年度气象影响有关,2016年降雨量相对较多,阴雨天气和较低的温度是木荷开花模式及造成花期延迟的重要环境因素。相对开花强度被认为是植物花资源空间分布的一个指标[22],它会影响到植物花粉的运动模式[23]。多数植物具有较低的相对开花强度[18,22],如准噶尔无叶豆(Eremospartonsongoricum(Litv.) Vass)[24]、血水草(EomeconchionanthaHance)[25]的相对开花强度分布频度均集中在10%~30%。本研究发现,与大多数植物一样,木荷种子园无性系具有较低的相对开花强度,分布频度集中在1%~30%之间。种子园无性系个体开花强度的偏斜率为1.88,高于Herrera[18]对30种灌木群体(1.59)和Buide等[22]对SileneacutifoliaLink ex Rohrb(1.45)的研究结果。木荷无性系开花强度频度分布严重偏斜,表明在种子园内具有相对开花强度极高的无性系(21和29号无性系)。开花物候对植物生殖成功具有明显的影响[21]。相关分析结果表明,木荷种子园无性系的开花数和开花持续时间分别与座果数存在极显著的正相关关系,此结果与马文宝等[24]和孙颖等[26]的研究结果一致。这些特点表明,对木荷种子园无性系而言,开花数量大,有利于吸引更多的传粉者,从而增加了传粉者对每朵花的传粉机会;而花期持续时间长,也更有利于提高传粉者的最后传粉效果,从而提高座果机会。 3.2 无性系花期同步性及利用 无性系花期同步性分析是种子园管理和无性系再选择要考虑的基本因素[10,27]。木荷种子园无性系组合间,年度内各无性系花期与其它无性系花期同步指数间均存在不同程度的差异,这进一步证实了无性系之间花期的同步性差异在林木种子园中是普遍存在的[6,12,28-29]。木荷种子园无性系的花期同步性指数在年度间基本相同,分别为0.758(2015年)和0.713(2016年),而在不同产地间无性系的花期同步指数存在明显差异,如17和21号(福建建瓯)与来自相同产地的(如福建建瓯的12和13号)无性系的花期同步指数均较高,而与来自其它产地的(如来自浙江龙泉的5号和福建南平的72号)无性系的花期同步指数均较低,这表明产地温度、海拔高度和经纬度等环境气候条件和遗传特性是影响木荷种子园无性系花期同步性的重要环境和遗传因素。各无性系花期同步指数存在极显著差异,变异系数25.8%,说明无性系之间进行同步指数的筛选和优化具有较大潜力。木荷种子园各无性系开花同步指数在年度间均有一定程度的变化,但年度间花期同步指数存在正相关关系,说明具有较高的参考价值。种子园无性系之间花期同步稳定性使得在整个花期的每个无性系均能接受到其它无性系的花粉,同时也能为其它无性系提供花粉[12]。2015年和2016年花期同步指数大于0.5的无性系的比例差别不大,分别为86%和87%,表明木荷种子园无性系年度间花期同步性处于相对稳定的状态。有学者指出,花期同步指数与环境条件(如温度、降雨量和海拔高度等)有密切的联系;如辐射松种子园无性系在高温、高降雨量年度的花期同步指数呈现显著下降的趋势[12],而油松种子园无性系在18 a时花期的连续降雨则使得无性系之间花期的同步性显著提高[10]。本研究中,木荷种子园2015年(24.4 ℃)和2016年(24.8 ℃)的温度无差异,2016年花期的连续降雨也未显著降低无性系之间花期同步性指数,因此,推测木荷花期同步性自身遗传特性影响的可能性较大。 连续2 a对木荷无性系种子园中19个无性系进行花期物候调查。相关分析结果表明,开花数多的无性系比开花数少的无性系花期更长、座果数更多,花期较长的无性系比花期较短的无性系座果数更多。无性系平均花期同步指数较高,但无性系间花期同步指数存在显著差异。年度间平均花期同步指数较为稳定,不存在显著变异。可根据花期同步指数对无性系进行筛选和优化,同时辅以人工授粉等措施弥补花期同步性差异,以达到提高种子园种子产量和质量的目的。 [1] 沈熙环. 种子园技术[M]. 北京:北京科学技术出版社, 1992. [2] 谭小梅, 金国庆, 张 一, 等. 截干矮化马尾松二代无性系种子园开花结实的遗传变异[J]. 东北林业大学学报, 2011, 39(4):39-42. [3] Ollerton J, Lack A J. Flowering phenology: An example of relaxation of natural selection?[J]. Trends in Ecology & Evolution, 1992, 7(8):274-276. [4] Abe T. Flowering phenology, display size, and fruit set in an understory dioecious shrub,Aucubajaponica(Cornaceae)[J]. American Journal of Botany, 2001, 88(3):455-461. [5] Primack R B. Relationships among flowers, fruits, and seeds[J]. Annual Review of Ecology & Systematics, 2003, 18(1):409-430. [6] Gunaga R P, Vasudeva R. Overlap index: a measure to assess flowering synchrony among teak (TectonagrandisLinn. f) clones in seed orchards[J]. Current Science, 2009, 97(6):941-946. [7] Torimaru T, Wang X R, Fries A,etal. Evaluation of pollen contamination in an advanced Scots pine seed orchard[J]. Silvae Genetica, 2009, 58(5):262-269. [8] 陈晓阳, 黄智慧. 杉木无性系开花物候对种子园种子遗传组成影响的数量分析[J]. 北京林业大学学报, 1995, (3):1-9. [9] Hansen O K. Mating patterns, genetic composition and diversity levels in two seed orchards with few clones—Impact on planting crop[J]. Forest Ecology & Management, 2008, 256(5):1167-1177. [10] 李 悦, 王晓茹, 李 伟, 等. 油松种子园无性系花期同步指数稳定性分析[J]. 北京林业大学学报,2010, 32(5):88-93. [11] Alexander L W, Woeste K E. Phenology, dichogamy, and floral synchronization in a northern red oak[J]. Canadian Journal of Forest Research, 2016, 46(5):629-636. [12] Codesido V, Merlo E, Fernandezlopez J. Variation in reproductive phenology in aPinusradiataD. Don seed orchard in northern Spain[J]. Silvae Genetica, 2005, 54(54):246-256. [13] Lyngdoh N, Gunaga R P, Joshi G,etal. Influence of geographic distance and genetic dissimilarity among clones on flowering synchrony in a Teak (TectonagrandisLinn. f) clonal seed orchard[J]. Silvae Genetica 2012, 61(1-2):10-18. [14] 方乐金, 施季森. 杉木种子园无性系结实稳定性的遗传变异[J]. 南京林业大学学报: 自然科学版, 2004, 28(1):17-20. [15] 中国科学院中国植物志编辑委员会. 中国植物志: 第49卷 第3分册 山茶科[M]. 北京:科学出版社, 1998:224. [16] 辛娜娜, 张 蕊, 徐肇友, 等. 不同产地木荷优树无性系生长和开花性状的分析[J]. 植物资源与环境学报, 2014, (4):33-39. [17] Dafni A. Pollination ecology: a practical approach[M]. Oxford : Oxford University Press, 1992:165-198 [19] 杜荣骞. 生物统计学[M]. 北京:高等教育出版社, 2009. [20] Ollerton J A. Evidence for stabilising selection acting on flowering time inArummaculatum(Araceae): the influence of phylogeny on adaptation[J]. Oecologia, 1999, 119(119):340-348. [21] Rathcke A B, Lacey E P. Phenological patterns of terrestrial plants[J]. Annual Review of Ecology & Systematics, 2003, 16(4):179-214. [22] Buide M L, Diazperomingo J A, Guitian J. Flowering phenology and female reproductive success inSileneacutifoliaLink ex Rohrb[J]. Plant Ecology, 2002, 163(1):93-103. [23] Dafni A. Practical pollination biology[M]. Cambridge, Ontario, Canada:Enviroquest Ltd,2005:3-26. [24] 马文宝, 施 翔, 张道远, 等. 准噶尔无叶豆的开花物候与生殖特征[J]. 植物生态学报, 2008, 32(4):760-767. [25] 陈 香, 胡雪华, 陆耀东, 等. 中国特有植物血水草开花物候与生殖特性[J]. 生态学杂志, 2011, 30(9):1915-1920. [26] 孙 颖, 王阿香, 刘颖竹, 等. 大花百子莲的开花物候与生殖特性[J]. 西北植物学报, 2013, 33(12):2423-2431. [27] Askew G R, Blush T D. Short note: an index of phenological overlap in flowering for clonal conifer seed orchards[J]. Silvae Genetica, 1990, 168-171. [28] El-Kassaby Y A, Ritland K, Fashler A M K. The role of reproductive phenology upon the mating system of a Douglas-fir seed orchard[J]. Silvae Genetica, 1988, 37(4):76-82. [29] 张 骁, 唐晓杰, 程广有. 黄檗种子园花期同步指数[J]. 东北林业大学学报, 2016, 44(7):46-50. (责任编辑:金立新) Flowering Phenology and Synchronization of Clones among Plant Ages in a Seed Orchard ofSchimasuperba YANGHan-bo,ZHANGRui,SONGPing,ZHOUZhi-chun (Research Institute of Subtropical Forestry, Chinese Academy of Forestry,Key Laboratory of Tree Breeding of Zhejiang Province, Hangzhou 311400, Zhejiang, China) [Objective]Studying the characteristic of flowering phenology and the variation and stability of flowering synchronization of clones. [Method] Based on data of florescence period and flower number of 19 clones in a seed orchard ofSchimasuperbawhich entered the stages of normal flowering and seed setting in 2015 and 2016, the flowering phenology and synchronization of each clone were estimated. [Result] The flowering of clones lasted for 10 to 28 days, and the first flowering date in 2016 was 2 days later than 2015. The correlation analysis showed that the clones with more flowers had a longer flowering duration and more fruit set than the clones with fewer flowers, and the longer the flowering duration, the more the fruit set. There were obvious difference of flowering synchronization among parent clone combinations. The index of flowering synchronization ranged from 0.552 to 0.857 and from 0.406 to 0.808, with an average of 0.758 and 0.713 in 2015 and 2016, respectively. More than 50% clones had a flowering synchronization (C) which greater than 0.8 in 2015, and more than 40% clones had aCvalue which greater than 0.8 in 2016. The coefficient of variation (CV) ofCvalue ranged from 12.016% to 46.476% and 15.375%-51.202%, respectively, during the year. The analysis of variance (ANOVA) showed that there was a significant variance ofCvalue among clones during the year at 0.01 level. The average ofCvalue among years was 0.737, and ANOVA analysis showed that there was no significant variance ofCvalue among years at 0.05 level. The coefficient of correlation between 2015 and 2016 was 0.229, indicating that there was a correlation ofCvalue among years, but it was not significant. [Conclusion] TheCvalue among years is stable in the seed orchard. We could screen and optimize the clones in seed orchard based on the flowering phenology and synchronization, and combined with hand-pollination to decrease the difference of flowering synchronization among clones, that will improve the yield and quality of seed inS.superbaseed orchard. Schimasuperba; seed orchard; flowering phenology; flowering synchronization; stability 10.13275/j.cnki.lykxyj.2017.04.003 2016-10-13 “十三五”浙江省农业(林木)新品种选育重大课题(2012C12908-6);“十二五”国家科技支撑计划课题(2012BA01B04);江西省林业厅林业科技创新专项项目(201503);福建省林木种苗科技攻关五期项目木荷课题 杨汉波(1988—),男,四川广安人,博士研究生,主要从事林木遗传育种研究. * 通讯作者:周志春,男,研究员,博士,主要从事林木遗传育种与培育技术研究.E-mail:zczhou_risf@163.com S722.8 A 1001-1498(2017)04-0551-082 结果与分析

3 讨论
4 结论