千岛湖被子植物枝叶性状分化及其与种多度的关系
2017-08-02余晶晶金毅郑诗璐胡广刘金亮袁金凤刘佳佳于明坚
余晶晶, 金毅, 郑诗璐, 胡广, 刘金亮, 袁金凤, 刘佳佳, 于明坚*
(1.浙江大学 生命科学学院, 浙江 杭州, 310058; 2.浙江理工大学 建筑工程学院, 浙江 杭州 310018)
千岛湖被子植物枝叶性状分化及其与种多度的关系
余晶晶1, 金毅1, 郑诗璐1, 胡广2, 刘金亮1, 袁金凤1, 刘佳佳1, 于明坚1*
(1.浙江大学 生命科学学院, 浙江 杭州, 310058; 2.浙江理工大学 建筑工程学院, 浙江 杭州 310018)
种间生态位分化是物种共存的重要原因,了解种间生态位分化特征对于推测物种多样性维持机制具有重要意义.植物的种间功能性状分化在一定程度上能够反映其生态位分化,进而导致植物在群落内的适合度差异.据此,本研究针对千岛湖地区75种常见木本被子植物,分析了8个与水分输导、资源获取和利用策略相关的功能性状分化,及其与物种多度的关系.主成分分析表明,种间功能性状的分化主要发生在比叶面积、叶绿素含量以及叶气孔密度上.8个性状中有5个,即叶绿素含量、叶片厚度、叶面积、比叶面积和最大树高具有一定程度的系统发育保守性(即Blomberg’s K的p<0.05). 功能性状间的相关性广泛存在,且在使用系统发育独立对比方法(phylogenetic independent contrast, PIC)控制系统发育的影响后,相关性大多仍然显著. 单个性状以及所有性状的主成分1和主成分2与物种多度间均没有显著相关性. 结果表明:千岛湖地区片段化次生马尾松林内木本被子植物的功能性状分化受系统发育和进化历史共同影响;且性状分化发生在主要的环境资源梯度 (即光照和水分)上;但性状对处于演替过程中的片段化森林群落内的物种多度不一定会产生重要影响.
功能性状;系统发育;片段化景观;马尾松林
植物的生态位需求在环境资源梯度上发生分化,即生态位分化 (niche differentiation),是物种共存的主要原因之一[1-2]. 与环境中主要资源 (如光照、水分等)梯度相关的生态位需求特性的分化,在很大程度上决定了物种共存的可能性范围[3-4]. 同时,植物的生理过程、对环境的适应策略等与其生态位相关的特性,可表征为功能性状[5]. VIOLLE等[6]指出,植物功能性状与其获取、利用和保存资源能力相关,并影响着植物个体适合度的形态、生理、物候等特征,即这些特征能间接影响物种在群落中的适合度. 因此,植物的生态位分化特点可体现为与其资源获取和利用策略相关的功能性状分化 (假设1)[7-8]. 同时,与资源策略 (resource strategy)相关的功能性状之间应存在协调性,以优化植物的资源获取和利用效率 (假设2)[9-10]. 虽然此类功能性状间的相关性可能是系统发育保守 (物种形成过程中其功能性状进化的保守性,导致亲缘关系近的物种间功能性状也倾向于较相似)所致,但也可能是性状间进化历史的相关性 (自然选择导致的不同功能性状间进化的协调性,以同步形成相应功能)的体现 (假设3)[11]. 与资源获取和利用相关的功能性状可影响植物的繁殖和更新,从而影响其在植物群落中的适合度,进而影响其物种多度 (假设4)[12-14].
本研究以我国亚热带东部的千岛湖片段化景观为平台,检验以上4个假设对片段化次生森林群落是否成立[15]. 具体地,主要以与植物资源获取和利用策略相关的叶和枝性状,以及物种的最大高度 (maximum height, MaxH)为研究对象,分析种间性状分化特点及其相关性,以检验假设1和2. 通过分析各性状的系统发育保守性,了解系统发育关系的相互影响;利用系统发育独立对比 (phylogenetic independent contrast, PIC)方法,去掉系统发育关系的影响后,分析性状间的相关性,了解其进化历史,以检验假设3. 最后,通过PIC方法校正后,分析性状与物种多度的相关性,以检验假设4. 值得注意的是,生境片段化和次生演替会影响生境条件和扩散等过程[16],从而影响植物更新[17],进而影响植物在群落中的多度以及物种共存情况[18]. 本研究有助于提高对植物资源的性状分化及协调性,以及功能性状与片段化森林群落结构 (例如物种多度)关系的认识.
1 材料和方法
1.1 研究地概况
研究地为浙江省淳安县境内的千岛湖(29°22′~29°50′ N, 118°34′~119°15′ E; 见图1), 建成于1959 年,拥有1 078 个陆桥岛屿 (最高水位108 m时). 库区水域面积约573 km2,平均水深为34 m,水库容积为1.784×1010m3. 该地区为典型亚热带季风气候,温暖湿润、四季分明. 年均气温17 ℃,年均降水量1 430 mm,上半年雨、热同季,下半年光照充足,冬夏长、春秋短,无霜期长. 森林土壤主要有红壤、黄壤和岩性土三大类,厚度中等、肥力较好. 库区的地带性植被为常绿阔叶林,但建坝时原有植被遭到严重破坏,岛上的树木基本被砍伐[19]. 目前,湖中岛屿和周边大陆的森林覆盖率高于85 %,以天然次生马尾松 (Pinusmassoniana)林为主,仅有10多个岛屿,如密山岛、猴岛等被开发为旅游景点,其他岛屿受到干扰较少[20].
图1 本研究所在的千岛湖库区Fig.1 The study region in Thousand Island Lake注 涂黄处为岛屿,涂灰处为大陆;彩图详见www.zjujournals.com/sci.
1.2.1 样品采集及测量方法
以人为干扰较少和植被主要为马尾松林为标准,于2009~2010 年选取千岛湖库区29 个岛屿作为固定的样岛进行植物群落调查. 研究样品的采样地点为整个千岛湖库区,采集对象为29 个样岛植物群落调查所记录的75 种木本被子植物. 每个物种随机选取5~10 株长势良好的成体,在取样个体的冠层向阳位置,随机枝剪20 张当年生且完整、健康、成熟的叶片和10 根完整、健康的枝条.
1.2.1.1 叶面积 (leaf area, LA)
植物的叶面积是评价植物光合能力、群落净初级生产力的重要指标.将采集的10片叶用干布清理干净,去除所有叶柄,将其平铺在坐标纸上,用透明塑料板压平,确保叶片完全展开、伸直,然后将所有叶片置于扫描仪计算叶片总面积. 如果叶片面积过大,可分多次扫描,相加后得到总面积.
1.2.1.2 比叶面积 (specific leaf area, SLA)
比叶面积为新鲜叶片的单面面积与叶片干重的比值. 它通常与光饱和速率、叶氮含量正相关,与叶寿命和碳相关的次生化合物如木质素则负相关[21].叶片干重:用干净的布清洁采集到的10张叶片 (去除叶柄),用电子天平称量10张叶片总鲜重;将称过的叶片装入纸袋,于烘箱中80 ℃烘48 h,取出称总干重.
萍萍抬起脸来看看我,她的脸上已经有泪水了,可是更多的还是惊魂未定的神色,似乎她也没有完全明白发生了什么,这时卫生间的门砰地打开了,林孟从里面出来时像是换了一个人,他撤完尿以后就平静下来了,他对我说:“你坐下。”
1.2.1.3 叶厚度 (leaf thickness, LT)
叶厚度在决定植物叶片物理结构强度中起关键作用[21]. 从采集的10 张叶片中,随机选取5张,用螺旋测微仪在每张叶片上随机选取3个不同位置 (避开叶脉,尽量选择叶片中心部位)重复测量,取平均值作为该个体叶片的厚度.
1.2.1.4 叶绿素含量 (chloroplast concentration, Chl)
叶绿素的相对含量能反映植物叶片的光合能力. 从采集的10 张叶片中,随机选取5张,每张叶片上随机选取3个不同位置,使用叶绿素仪重复测量其叶绿素相对含量,取平均值作为该个体叶片的叶绿素含量.
1.2.1.5 气孔密度 (stomata density, StoD)
气孔是植物与外界环境进行水分和气体交换的主要通道,其变化反映了植物水分供给与需求之间的平衡[22]. 从采集到的叶片中随机选取5张,在其下表皮随机选取3个点 (约1 cm2,注意避开主脉和大的次级脉)制作水装片,在光学显微镜下随机选取3 个视野,分别统计气孔数量,每张叶片的气孔密度为3 个视野观测到的气孔个数平均值与视野面积之比值.
1.2.1.6 叶干物质含量(leaf dry matter concen- tration, LDMC)
叶干物质含量为叶片干重与叶片鲜重的比值,与叶片组织的平均密度有关[21].
1.2.1.7 木质密度 (wood density, WD)
木质密度是反映植物在自身构建投资策略中碳的分配比重的重要参数. 枝条鲜重:去除枝条上的叶、花、果实和其他附属物,将枝条从近端 (靠近二年生枝条处)开始截取约10 cm,用电子天平称量总鲜重. 枝条体积:将称量过鲜重的10 根枝条用排水法测量其总体积. 枝条干重:将测量过体积的枝条装入纸袋,烘箱里80 ℃烘72 h,取出称量总干重. 木材密度=枝条总干重/枝条总体积.
1.2.1.8 最大高度 (maximum height, MaxH)
植株高度是决定植物个体对光资源竞争能力的主要因素之一,反映物种在光照资源梯度上的分化特点[23]. 植株最大高度参考《中国在线植物志》 (www.eflora.cn).
1.2.1.9 物种多度(abundance)
物种多度为2014~2015 年对千岛湖库区29 个岛屿样地群落进行复查所记录到的胸径 (diameter at breast height, DBH)≥1 cm的每个物种的个体总数.
1.2.2 系统发育树构建
本研究所使用的系统发育树 (phylogeny)来源于QIAN等[24],由ZANNE等[25]的系统发育树优化得到. 对于本研究所涉及的但在该系统发育树中不存在的物种,采用BLADJ (blanch adjuster)方法 (即QIAN等[24]的“scenario 3”)调整物种枝长后,组装到系统发育树上,然后去掉本研究不涉及的种,得到所使用的系统发育树.
1.2.3 统计分析
主成分分析 (principle component analysis, PCA)采用prcomp方程实现. 相关性分析使用Pearson检验,采用cor.test方程实现. 系统发育保守性分析使用Blomberg’s K指数[26],采用phylosignal方程实现. 系统发育对比方法 (phylogenetic independent contrasts, PIC)采用picante软件包中的pic方程实现[27]. 本研究所有分析均在R3.2.4软件中进行[28].
2 结 果
研究75种千岛湖常见木本被子植物,分属于31个科(见附表).
PCA结果表明,轴PC1的解释度为39 %,该轴主要与叶绿素含量和SLA等性状相关;轴PC2的解释度为20 %,该轴主要与气孔密度等性状相关 (见图2、 表1).Blomberg’s K分析发现,8个性状中,除气孔密度、叶干物质含量和木质密度外,其余均显现一定的系统发育保守性 (见表2).

表1 各性状与主成分1和2间的相关性
注 黑体代表数值的p<0.05.
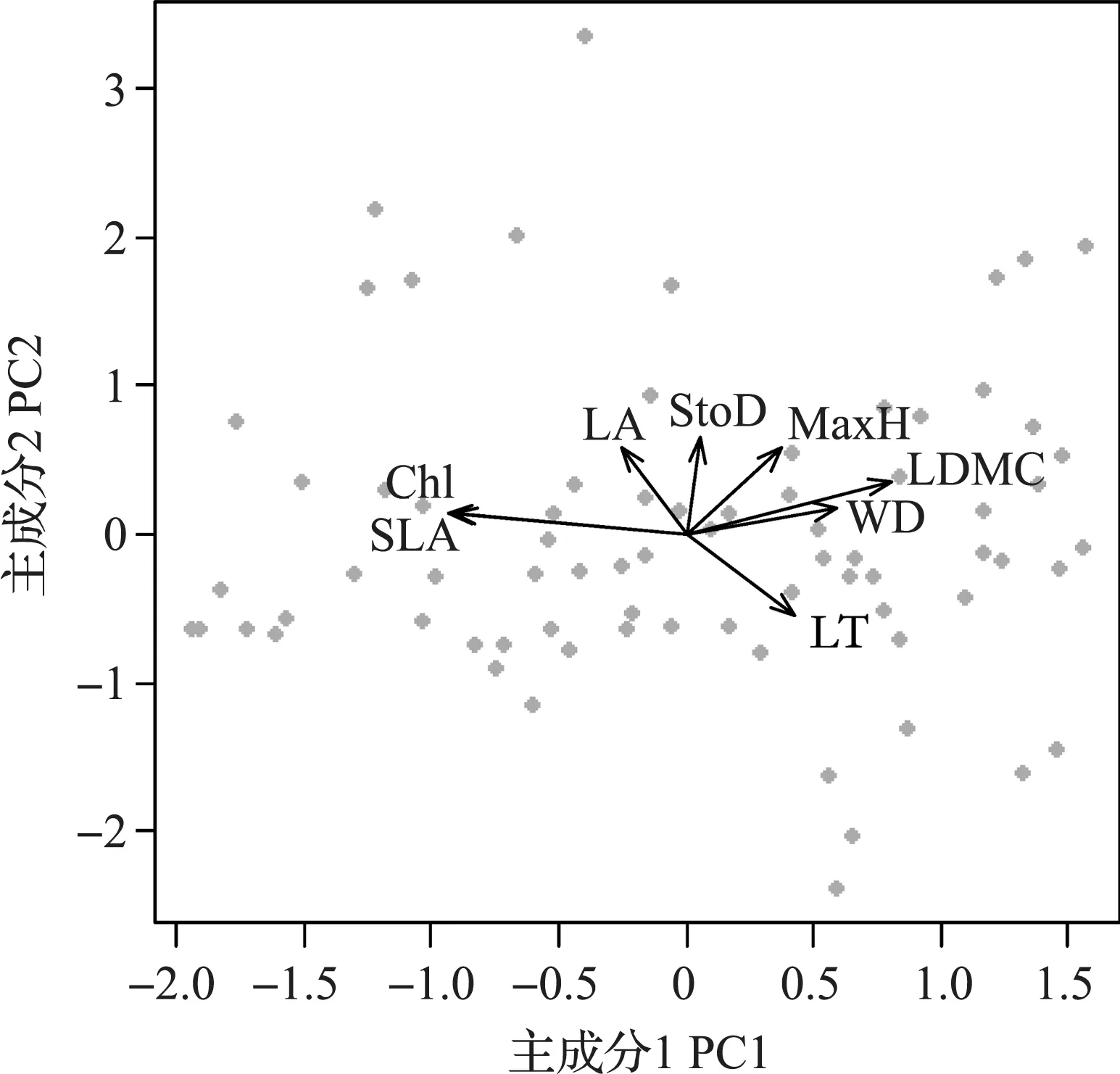
图2 各性状间的主成分分析结果Fig.2 Results of principle component analysis of the study traits 注 Chl表示叶绿素含量;LT表示叶厚度;StoD表示气孔密度;LA表示叶面积;SLA表示比叶面积;LDMC表示叶干物质含量;WD表示木质密度;MaxH表示最大高度. 灰色实心圆表示物种.
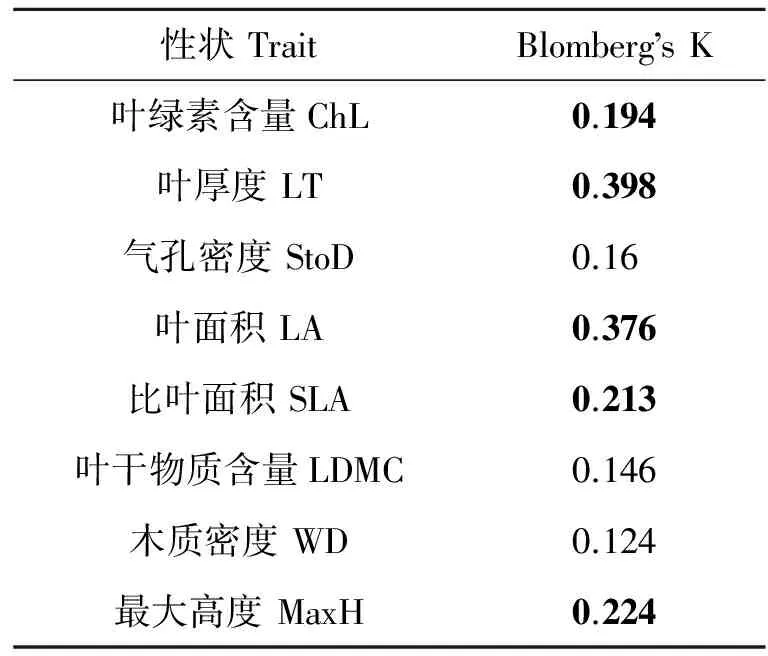
表2 各性状的系统发育信号
注 黑体代表数值的p<0.05.
Pearson检验表明,叶绿素含量与叶面积、SLA正相关,而与除气孔密度外的其他性状负相关;叶厚度与SLA负相关;叶面积与最大高度正相关,而与木质密度负相关.SLA与叶干物质含量、木质密度和最大高度负相关;叶干物质与木质密度和最大高度正相关.气孔密度与所有的性状指标均无相关性.使用PIC方法去掉系统发育关系对性状的影响后,大多数性状间的相关性仍然显著,少数性状间的相关性发生变化.例如,叶绿素含量与叶面积、SLA和最大高度之间、SLA与木质密度之间,以及叶干物质含量与最大高度之间的相关性消失;气孔密度与SLA和木质密度呈现正相关关系(见表3).
使用PIC方法去掉系统发育的影响后,进行Pearson检验发现,8个性状及其PC1和PC2与物种多度均相关性不强(见表4).
3 讨 论
研究发现千岛湖地区75种木本被子植物的功能性状分化主要发生在叶绿素含量和比叶面积,以及气孔密度的分化方向上.其中,叶绿素含量和比叶面积与叶光合能力(例如Amass)及叶寿命(leaf lifespan)紧密相关,为传统的叶经济谱策略(leaf economics spectrum)研究的重要性状[7-8,29-30],该类性状的差异反映了植物生态位需求在环境中光照等梯度上的分化.气孔密度属于叶的水分性状(hydraulic traits)之一[22],与植物水分输导和利用效率紧密相关[31-32],该类性状的差异主要反映植物的生态需求在水分等梯度上的分化情况[31,33].REICH等[9]推测以上两类性状之间应存在紧密联系,因为两类性状共同为光合作用等过程服务[8,34].而LI等[22]对中国低纬度不同区域的热带和亚热带森林中85个优势种的综合分析发现,该两类性状是可以独立进化的,并推测其原因可能是:与水分性状和传统的经济谱性状(economics traits)相关的叶结构间相互独立,且该独立性可影响叶功能;与水分性状和非水分性状相关的叶结构进化历史迥异;两类性状的独立进化可使植物能够适应广阔的环境条件范围.本研究针对千岛湖地区亚热带森林内的木本被子植物类群,发现该两类性状也是相对独立的(见图2、表3).同时,发现千岛湖地区常见木本被子植物的功能性状分化主要集中在水分和经济谱性状的分化上,表明环境资源梯度可能对本研究地区植物的种间生态位分化发挥着重要作用,支持假设1.该结果也与在其他地区所进行的多个研究结果一致[7,9,35].
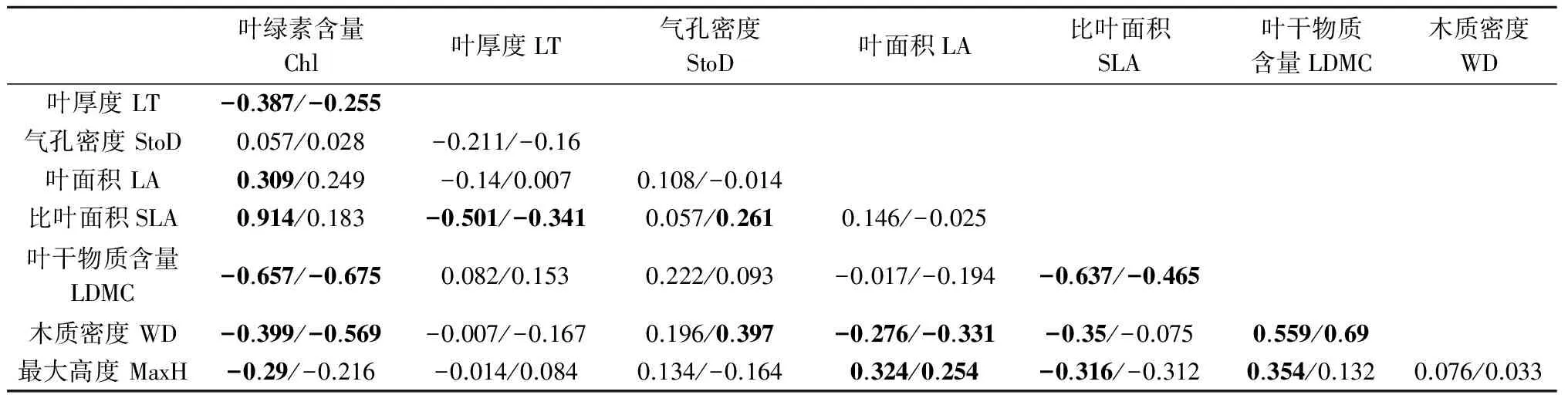
表3 性状间相关性
注 /左侧和右侧分别为PIC处理前后的相关性值;黑体代表数值的p<0.05.

表4 系统发育独立对比方法调整后物种多度与各性状间的相关性
注 主成分1和2分别表示8个性状经主成分分析所得的主成分轴1和轴2上的数值;黑体代表数值的p<0.05.
本研究所涉及的千岛湖地区75 种木本被子植物的叶面积、比叶面积、叶厚度、叶绿素含量和最大树高这5 个性状均检测到显著的系统发育信号,说明它们在进化过程中均受到系统发育关系一定程度的限制 (见表2). 同时,曹科等[12]对于古田山亚热带常绿阔叶林中157 个物种叶功能性状的分析结果也表明,存在显著的系统发育信号. 这2个研究结果表明,千岛湖和古田山所在的浙江省中亚热带地区木本被子植物功能性状的进化可能广泛地受到系统发育关系的限制.
以往研究发现,叶经济谱不同性状之间存在相关性[8,36-37]. 本研究也发现叶经济谱性状间的相关性广泛存在. 例如,叶厚度、叶绿素含量和SLA等性状间的显著相关性 (见表3). 该结果支持假设2,也与以往在世界其他地区进行的相关研究结果一致[7,36,38-39]. 值得注意的是,当使用PIC方法控制系统发育关系影响后,发现原本并无相关性的气孔密度和SLA之间变得显著正相关了,表明水分性状和经济谱性状的进化历史具有相关性[8,40],同时也表明系统发育关系可能会掩盖性状进化历史之间的关系,因此在分析性状间的进化史关系时有必要去掉系统发育的影响[22],尤其是已明确了性状 (例如,本研究中的SLA)进化受到系统发育关系限制时. 另外,还发现PIC控制系统受发育关系影响后,叶绿素含量与其他3个性状间的相关性消失 (见表3),表明其相关性可能受到系统发育关系的影响较大,而进化历史的影响可能较小. 木质密度和SLA间的相关性消失,在一定程度上反映了水分性状与经济谱性状在进化上的独立性. 而最大高度与叶干物质含量的相关性消失,也体现了两者进化史的独立性. 另外,SLA与气孔密度的正相关性变得显著,体现两者在进化上具有协调性,与预测一致. 而气孔密度与木质密度却呈现相同的变化趋势,与预测相反,进一步研究有助于了解其原因. 以上结果部分支持假设3.
物种多度在一定程度上反映了物种在群落中的适合度[14,41]. 多度较大的物种其适合度可能较高,多度较小的物种其适合度可能较低[12,14]. 因此,千岛湖地区的植物功能性状与其多度无相关性,表明群落内物种的资源策略差异可能对植物的适合度并不存在较大影响,不支持假设4. 然而,曹科等[12]发现古田山亚热带常绿阔叶林样地内木本植物的比叶面积、木质密度、氮含量与物种多度呈极显著正相关,磷含量与物种多度呈较为显著的正相关,叶面积与物种多度呈较显著的负相关. 本研究与曹科等[12]结果的差异性,可能与千岛湖和古田山森林群落所处演替阶段不同以及片段化等因素有关. 古田山样地内亚热带常绿阔叶林没有受到片段化影响,且已处于演替后期[42]. 该群落内适合度较高 (即多度较大)的植物可能主要为耐阴植物,该类植物应主要为能够在林内低资源水平下完成生活史的物种[43], 通常具有保守的经济策略 (conservative economics strategy),可表现为具有较低的SLA、叶氮磷含量和较高的木质密度等. 曹科等[12]的结果也与这些预测相符,表现为物种多度随SLA和叶氮磷含量的升高而降低,而随木质密度的增大而提高. 然而,千岛湖地区的植物群落类型主要为片段化次生马尾松林[15],为干扰后演替早期阶段森林,林内环境资源水平可能较高[44],导致耐阴植物在林内的更新优势较小[45]. 同时,片段化导致森林斑块受到外界环境影响通常较完整森林更大,因此片段化森林内的资源水平异质性大[46-47],使得不同耐阴性植物都能在该类型森林中更新,进而导致千岛湖地区的森林群落内植物功能性状与物种多度间缺乏相关性.
理论上,森林演替进程中,林内资源水平会降低,导致耐阴植物的适合度升高而不耐阴植物的适合度降低,进而导致耐阴植物逐渐取代不耐阴植物[48]. 田云等[49]近期对千岛湖岛屿次生马尾松林内不同功能群落植物幼苗动态的对比研究结果也表明,耐阴植物幼苗有取代不耐阴植物幼苗的趋势. 对于演替进程中,千岛湖地区片段化的岛屿次生马尾松林群落内植物功能性状如何影响植物适合度,进而影响物种多度,需要长期的跟踪监测.
浙江淳安县新安江开发总公司对项目研究工作提供了支持,野外实验过程中得到了南歌、骆杨青、戎褔仁、吴奕如等同学和当地村民的大力帮助,在此致以最衷心的感谢!
[1] MCGILL B J, ENQUIST B J, WEIHER E, et al. Rebuilding community ecology from functional traits[J]. Trends in Ecology & Evolution, 2006,21(4):178-185.
[2] ADLER P B, FAJARDO A, KLEINHESSELINK A R, et al. Trait-based tests of coexistence mechanisms[J]. Ecology Letters,2013,16(10):1294-1306.
[3] FARGIONE J, TILMAN D. Niche differences in phenology and rooting depth promote coexistence with a dominant C4 bunchgrass[J]. Oecologia,2005,143(4):598-606.
[4] KRAFT N J B, GODOY O, LEVINE J M. Plant functional traits and the multidimensional nature of species coexistence[J]. Proceedings of the National Academy of Sciences,2015,112(3):797-802.
[5] DIAZ S, CABIDO M N I. Can grazing response of herbaceous plants be predicted from simple vegetative traits[J]. Journal of Applied Ecology,2001,38(3):497-508.
[6] VIOLLE C, NAVAS M L, VILE D, et al. Let the concept of trait be functional[J]. Oikos,2007,116(5):882-892.
[7] WRIGHT I J, REICH P B, WESTOBY M, et al. The worldwide leaf economics spectrum[J]. Nature,2004,428(6985):821-827.
[8] REICH P B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto[J]. Journal of Ecology,2014,102(2):275-301.
[9] REICH P B, WRIGHT I J, CAVENDERBARES J, et al. The evolution of plant functional variation: Traits, spectra, and strategies[J]. International Journal of Plant Sciences,2015,164(S3):143-164.
[10] AUBIN I, MUNSON A D, CARDOU F, et al. Traits to stay, traits to move: A review of functional traits to asses[J]. Environmental Reviews,2015,24(2):1-23.
[11] KRAFT N J B, ACKERLY D D. Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest[J]. Ecological Monographs,2010,80(3):401-422.
[12] 曹科,饶米德,余建中,等.古田山木本植物功能性状的系统发育信号及其对群落结构的影响[J].生物多样性,2013,21(5):564-571. CAO K, RAO M D, YU J Z, et al. The phylogenetic signal of functional traits and their effects on community structure in an evergreen broad-leaved forest[J]. Biodiversity Science,2013,21(5):564-571.
[13] 刘晓娟,马克平.植物功能性状研究进展[J].中国科学:生命科学,2015,45(4):325-339. LIU X J, MA K P. Plant functional traits-concepts, applications and future directions[J]. Science in China: Earth Sciences,2015,45(4):325-339.
[14] SHIPLEY B, BELLO F D, CORNELISSEN J H C, et al. Reinforcing loose foundation stones in trait-based plant ecology[J]. Oecologia,2016,180(4):1-9.
[15] YU M, HU G, FEELEY K J, et al. Richness and composition of plants and birds on land-bridge islands: Effects of island attributes and differential responses of species groups[J]. Journal of Biogeography,2012,39(6):1124-1133.
[16] SUTTON F M, MORGAN J W. Functional traits and prior abundance explain native plant extirpation in a fragmented woodland landscape[J]. Journal of Ecology,2009,97(4):718-727.
[17] HANSKI I. Habitat fragmentation and species richness[J]. Journal of Biogeography,2015,42(5):989-993.
[18] FAHRIG L. Rethinking patch size and isolation effects: The habitat amount hypothesis[J].Journal of Biogeography, 2013,40(9):1649-1663.
[19] 徐高福,章德三,张立功,等.千岛湖森林资源动态变化分析[J].林业调查规划,2005,30(2):25-28. XU G F, ZHANG D S, ZHANG L G, et al. Forest resources dynamics in the Thousand Island Lake[J]. Forest Inventory and Planning,2005,30(2):25-28.
[20] 苏晓飞,袁金凤,胡广,等.千岛湖陆桥岛屿植物群落结构的边缘效应[J].应用生态学报,2014,25(1):77-84. SU X F, YUAN J F, HU G, et al. Edge effect of the plant community structure on land-bridge islands in the Thousand Island Lake[J]. Chinese Journal of Applied Ecology,2014,25(25):77-84.
[21] PÉREZ-HARGUINDEGUY N, GARNIER E, LAVOREL S, et al. New handbook for standardized measurement of plant functional traits worldwide[J]. Australian Journal of Botany,2013,61(3):167-234.
[22] LI L, MCCORMACK M L, MA C, et al. Leaf economics and hydraulic traits are decoupled in five species-rich tropical-subtropical forests[J]. Ecology Letters,2015,18(9):899-906.
[24] QIAN H, JIN Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure[J]. Journal of Plant Ecology,2016,9(2):233-239.
[25] ZANNE A E, TANK D C, CORNWELL W K, et al. ZANNE et al. reply[J]. Nature,2015,521(7552):6-7.
[26] BLOMBERG S P, JR T G. Tempo and mode in evolution: Phylogenetic inertia, adaptation and comparative methods[J]. Journal of Evolutionary Biology,2002,15(6):899-910.
[27] KEMBEL S W, COWAN P D, HELMUS M R, et al. Picante: R tools for integrating phylogenies and ecology[J]. Bioinformatics,2010,26(11):1463-1464.
[28] R Core Team. R: A language and environment for statistical computing[EB/OL]. https://www.r-project.org/(R Foundation for Statistical Computing),2016.
[29] CASSON S A, HETHERINGTON A M. Environmental regulation of stomatal development[J]. Current Opinion in Plant Biology,2009,13(1):90-95.
[30] 刘畅,孙鹏森,刘世荣.植物反射光谱对水分生理变化响应的研究进展[J].植物生态学报,2016,40(1):80-91. LIU C, SUN P S, LIU S R. A review of plant spectral reflectance response to water physiological changes[J]. Chinese Journal of Plant Ecology,2016,40(1):80-91.
[31] SACK L, SCOFFONI C. Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future[J]. New Phytologist,2013,198(4):983-1000.
[32] 王瑞丽,于贵瑞,何念鹏,等.气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例[J].生态学报,2016,36(8):2175-2184. WANG R L, YU G R, HE N P, et al. Altitudinal variation in the covariation of stomatal traits with leaf functional traits on Changbai Mountain[J]. Acta Ecologica Sinica,2016,36(8):2175-2184.
[33] FRANKS S J, WEIS A E. Climate change alters reproductive isolation and potential gene flow in an annual plant[J]. Evolutionary Applications,2009,2(4):481-488.
[34] BLONDER B, VIOLLE C, BENTLEY L P, et al. Venation networks and the origin of the leaf economics spectrum[J]. Ecology Letters,2011,14(2):91-100.
[35] FRESCHET G T, CORNELISSEN J H C, VAN LOGTESTIJN R S P, et al. Evidence of the ‘plant economics spectrum’ in a subarctic flora[J]. Journal of Ecology,2010,98(2):362-373.
[36] REICH P B, ELLSWORTH D S. From tropics to tundra: Global convergence in plant functioning[J]. Proceedings of the National Academy of Sciences,1997,94(25):13730-13734.
[37] LORANGER J, SHIPLEY B. Interspecific covariation between stomatal density and other functional leaf traits in a local flora[J]. Botany-botanique,2010,88(1):30-38.
[38] FORTUNEL C, FINE P V A, BARALOTO C. Leaf, stem and root tissue strategies across 758 Neotropical tree species[J]. Functional Ecology,2012,26(5):1153-1161.
[39] OSNAS J L, LICHSTEIN J W, REICH P B, et al. Global leaf trait relationships: Mass, area, and the leaf economics spectrum[J]. Science,2013,340(6133):741-744.
[40] SHIPLEY B, VU T T. Dry matter content as a measure of dry matter concentration in plants and their parts[J]. New Phytologist,2002,153(2):359-364.
[41] CHENG J, MI X, NADROWSKI K, et al. Separating the effect of mechanisms shaping species-abundance distributions at multiple scales in a subtropical forest[J]. Oikos,2012,121(2):236-244.
[42] 祝燕,赵谷风,张俪文,等.古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构[J].植物生态学报,2008,32(2):262-273. ZHU Y, ZHAO G F, ZHANG L W, et al. Community composition and structure of Gutianshan forest dynamics plot in a mid-subtropical evergreen broad-leaved forest, east China[J]. Journal of Plant Ecology,2008,32(2):262-273.
[43] 宋永昌.中国东部常绿阔叶林生态系统退化机制与生态恢复[M].北京:科学出版社,2007. SONG Y C. Degradation Mechanism and Ecological Restoration of Evergreen Broad-Leaved Forest Ecosystem in East China[M]. Beijing:Science Press,2007.
[44] LAURANCE W F, PéREZ-SALICRUP D, DELAMNICA P, et al. Rain forest fragmentation and the structure of Amazonian Liana communities[J].Ecology,2001,82(1):105-116.
[45] POORTER L, BONGERS F. Leaf traits are good predictors of plant performance across 53 rain forest species[J]. Ecology,2006,87(7):1733-1743.
[46] BENDER D J, CONTRERAS T A, FAHRIG L. Habitat loss and population decline: A meta-analysis of the patch size effect [J]. Ecology,1998,79(2):517-533.
[47] COOK W M, YAO J, FOSTER B L, et al. Secondary succession in an experimentally fragmented landscape: Community patterns across space and time[J]. Ecology,2005,86(5):1267-1279.
[48] MOLINO J F, SABATIER D. Tree diversity in tropical rain forests: A validation of the intermediate disturbance hypothesis[J]. Science,2001,294(5547):1702-1704.
[49] 田云,金毅,王志平,等.千岛湖岛屿马尾松林不同耐阴性植物幼苗动态研究[J].浙江大学学报:理学版,2016,43(4):70-78. TIAN Y, JIN Y, WANG Z P, et al. Seedling dynamics of shade tolerant and intolerant woody plants in the Masson pine forests on islands of the Thousand Island Lake[J]. Journal of Zhejiang University:Science Edition,2016,43(4):70-78.
Differentiation in leaf and branch traits of angiosperms and their relationships between species abundance in the Thousand Island Lake Region.
YU Jingjing1, JIN Yi1, ZHENG Shilu1, HU Guang2, LIU Jinliang1, YUAN Jinfeng1, LIU Jiajia1, YU Mingjian1
(1.CollegeofLifeSciences,ZhejiangUniversity,Hangzhou310058,China; 2.SchoolofCivilEngineeringandArchitecture,ZhejiangSci-TechUniversity,Hangzhou310018,China)
Niche differentiation is a major determinant of species coexistence, and hence understanding how species niche differentiates is beneficial to our understanding of the mechanisms underlying species diversity maintenance. Since interspecific plant functional trait differentiation represents the niche differentiation to some extent, we have studied the differentiation of eight functional traits related to water conductance, resource acquisition and utilization in 75 common woody angiosperm species in the Thousand Island Lake Region. We find that the overall interspecific functional trait differentiation is mainly associated with interspecific differentiation in specific leaf area (SLA), chloroplast concentration (Chl) and stomata density (StoD) by principle component analysis (PCA). Five traits (i.e., chloroplast concentration (Chl), leaf thickness (LT), leaf area (LA), specific leaf area (SLA) and maximum height (MaxH) are constrained by phylogeny (i.e.,p<0.05 for Blomberg’s K) to some extent. A great number of significant pairwise trait correlations are detected, and after PIC adjustment, these significant correlations mostly remaine. We also find that the single trait and the principle component 1 (PC1) and principle component 2 (PC2) values of all traits are not related to species abundance. Our results suggest that the functional trait differentiation of angiosperm species in the fragmented successional Masson pine forest of the Thousand Island Lake Region are constrained by both phylogeny and evolutionary history; And, the functional traits differentiate in major environmental resource axes (i.e., irradiance and water); But they may not relate to species abundance in the successional stage of fragmented forests.
functional trait; phylogeny; fragmented landscape; Masson pine forest

附表 千岛湖样地内75种木本被子植物名录

物种Species拉丁名Scientificname科Family杨梅Myricarubra(Lour.)Sieb.etZucc.杨梅科乌楣栲CastanopsisjucundaHance壳斗科苦槠C.sclerophylla(Lindl.)Schott.壳斗科青冈栎Cyclobalanopsisglauca(Thunb.)Oerst.壳斗科石栎Lithocarpusglaber(Thunb.)Nakai壳斗科麻栎QuercusacutissimaCarr.壳斗科白栎Q.fabriHance壳斗科短柄枹Q.serrataThunb.var.brevipetiolata(A.DC.)Nakai壳斗科山油麻Tremacannabinavar.dielsiana(Hand.⁃Mazz.)C.J.Chen榆科榔榆UlmusparvifoliaJacq.榆科小构树BroussonetiakazinokiSieb.etZucc.桑科山腊梅ChimonanthusnitensOliv.腊梅科樟Cinnamomumcamphora(Linn.)Presl樟科乌药Linderaaggregata(Sims)Kosterm.樟科山胡椒L.glauca(Sieb.etZucc.)Bl.樟科山橿L.reflexaHemsl.樟科豹皮樟Litsearotundifolia(Nees)Hemsl.var.oblongifolia(Nees)Allen樟科山鸡椒L.cubeba(Lour.)Pers.樟科檫木Sassafrastzumu(Hemsl.)Hemsl.樟科长圆叶鼠刺IteaoblongaHand.⁃Mazz.虎耳草科枫香LiquidambarformosanaHance金缕梅科

(续附表)
2016-07-22.
国家自然科学基金资助项目(31361123001,31570524,31500382).
余晶晶(1990-),ORCID:http:∥orcid.org/0000-0002-4721-2495,女,硕士研究生,主要从事生物多样性和植物生态学研究.
*通信作者,ORCID:http:∥orcid.org/0000-0001-8060-8427,E-mail:fishmj202@hotmail.com.
10.3785/j.issn.1008-9497.2017.04.009
Q 948
A
1008-9497(2017)04-437-09
Journal of Zhejiang University(Science Edition), 2017,44(4):437-445
