茶树锰超氧化物歧化酶基因CsMSD克隆及其表达分析
2017-07-18赖钟雄金琦芳郭玉琼常笑君
王 仲,赖钟雄,金琦芳,郭玉琼,常笑君
(1.贺州学院 食品与生物工程学院,广西 贺州 542899;2.福建农林大学 园艺植物生物工程研究所,福建 福州 350002)
茶树锰超氧化物歧化酶基因CsMSD克隆及其表达分析
王 仲1,赖钟雄2,金琦芳1,郭玉琼2,常笑君2
(1.贺州学院 食品与生物工程学院,广西 贺州 542899;2.福建农林大学 园艺植物生物工程研究所,福建 福州 350002)
为了解茶树CsMSD在干旱胁迫下的表达规律,采用RT-PCR和RACE技术,对茶树干旱胁迫响应相关重要基因锰超氧化物歧化酶基因CsMSD进行克隆和生物信息学分析,并采用实时荧光定量PCR技术,对其在不同程度干旱胁迫下的表达规律进行分析。结果显示,茶树CsMSD核苷酸序列全长1013bp,开放阅读框长693bp,共编码230个氨基酸;CsMSD蛋白分子量为25.47kD,理论等电点pI=6.38,存在9个磷酸化位点,亚细胞定位于线粒体中,且蛋白质序列中具有MSD特征结构域;qPCR分析干旱胁迫下茶树叶片中CsMSD的表达变化情况发现,CsMSD在干旱胁迫下的相对表达量与对照组相比显著上调。研究认为铁观音茶树CsMSD在应对干旱胁迫中发挥着重要的作用。
铁观音茶树;CsMSD;基因克隆;干旱胁迫;qPCR
植物体内超氧化物歧化酶(Superoxide dismutase,SOD)是一类与清除活性氧有关的金属酶,对保护细胞免受氧化损伤具有十分重要的作用,是植物防御系统中第一个清除活性氧的关键酶[1]。1969年,McCord等[2]首次发现了SOD在生物体内能够歧化超氧阴离子。植物中SOD依据活性中心金属辅因子不同主要有三类:CuZnSOD、FeSOD和MnSOD[3]。不同类型SOD在植物细胞中的亚细胞定位不同,MnSOD可能存在于线粒体和过氧化物酶体中。其中线粒体是细胞内产生超氧阴离子的主要场所,MnSOD可能在保护细胞免受超氧阴离子损伤中发挥重要作用[4]。
干旱是我国茶园生产的重要限制因素,一方面我国不同地区降水分布不均;另一方面茶园多地处丘陵或山地等,水资源不足而灌溉设施不够完善。茶树中抗旱生理研究[5]表明,对茶树超氧化物歧化酶、丙二醛、可溶性糖、过氧化物酶和游离脯氨酸与茶树抗旱指数进行灰色关联度分析,其中超氧化物歧化酶与茶树抗旱的关联程度最大。超氧化物歧化酶活性可以作为茶树抗旱能力强弱的重要指示标志。目前,茶树干旱胁迫下超氧化物歧化酶相关研究主要集中在酶活性的变化上[6,7],而对其基因表达层面的研究较少。鉴于此,本研究以铁观音茶树叶片为试验材料,通过同源克隆获得茶树超氧化物歧化酶基因家族重要成员CsMSD基因序列,并检测其在干旱胁迫下的表达规律。以期为进一步研究CsMSD基因在茶树抵御干旱胁迫中的作用奠定基础。
1 材料与方法
1.1 试验材料及试剂
以铁观音茶树叶片为材料,提取总RNA用于后续试验。Tripure Isolation Reagent购于 Roche,SMART RACE反转录试剂盒购于Clontech公司,SYBR ExScriptTM试剂盒、SYBR Premix Ex TapTM试剂盒、pMD18-T载体、DH5α 感受态细胞为Takara公司产品。
1.2 试验方法
1.2.1 茶树叶片总RNA提取及cDNA合成
参照赵姗姗等[8]的RNA提取和检测方法,按照SMART RACE说明书合成第一链cDNA。
1.2.2 茶树CsMSD基因克隆引物设计及PCR扩增
根据实验室已有的铁观音茶树转录组数据,设计基因保守区或者ORF上下游引物,采用RT-PCR扩增获得其保守序列。获得CsMSD保守区序列后,进一步设计3’端和5’端RACE引物,进行巢式PCR扩增获得各成员的末端序列。试验引物委托厦门闽博公司合成,试验测序委托上海博尚进行。用于CsMSD基因克隆的所有引物序列相关信息及用途见表1。
配置25μl的PCR反应体系,其中12.5μl的Tap Mix,1μl的 cDNA 模板,10μM/L 的上下游引物各1μl,9.5μl的H2O。PCR扩增反应程序设定为:94℃预变性 3min;94℃变性 30s,退火 30s,72℃延伸,共35个循环;最后72℃延伸8min;根据引物特性设定相应的退火温度,根据扩增产物长度调整延伸时间。
1.2.3 生物信息学分析
使用DNAMAN翻译CsMSD最大开放阅读框的蛋白质,使用NCBI进行CsMSD的氨基酸同源性分析,使用ExPASy ProtParam在线工具预测CsMSD蛋白的基本理化性质,使用Softberry ProtComp 9.0预测亚细胞定位情况,使用Chlorop1.1预测其是否存在信号肽,使用Tmpred预测CsMSD蛋白的跨膜结构,使用SOPMA分析CsMSD蛋白二级结构,使用NetPhoS对CsMSD蛋白进行磷酸化位点分析,使用COILS Server软件对CsMSD蛋白卷曲螺旋结构进行预测,使用PredictProtein在线软件对CsMSD蛋白功能位点进行预测与分析,采用Mega 6近邻相接法(Neighbor-Joining,NJ)对来自不同植物的 MSD氨基酸序列构建系统进化树。

表1 茶树CsMSD基因克隆相关引物信息
1.2.4 实时荧光定量表达分析
刘玉英等[9]研究7个茶树品种在干旱胁迫下叶片中SOD活性的变化规律,结果均呈现“先升高后下降”的变化趋势。综合其他相关研究,得出茶树SOD活性在干旱胁迫10天左右时达到最高,因此对铁观音茶树盘栽苗进行干旱胁迫处理,胁迫10天后下午进行取样。干旱胁迫处理为:对照CK正常供水,土壤含水量/田间持水量为80(±5)%;T1轻度干旱胁迫,土壤含水量/田间持水量为60(±5)%;T2中度干旱胁迫,土壤含水量/田间持水量为40(±5)%;T3重度干旱胁迫,土壤含水量/田间持水量为20(±5)%。提取经不同处理的铁观音茶树叶片总RNA,经反转录成cDNA作为模板,采用相应的引物进行qPCR扩增。
参考孙美莲等[10]筛选的内参引物序列,选取18S作为内参基因,检测CsMSD基因在不同干旱胁迫程度下的相对表达量。在CsMSD基因的保守区设计特异性引物进行实时荧光定量qPCR扩增,其中上游引物序列为 5’-GCGTGCTCCCAAACATCT AAG-3’,下游引物序列为 5’-TTACAAGGCTCTGG ATGGGTG-3’,扩增温度为 62℃。
2 结果与分析
2.1 茶树锰超氧化物歧化酶基因CsMSD克隆
采用各对引物分别进行PCR扩增,扩增得到保守区序列500bp以上(图1)。进一步在保守区上设计引物进行5’端RACE、3’端RACE扩增,将相应产物进行回收、连接和转化,选取阳性克隆子进行测序。将测序所得序列在NCBI数据库进行BLASTn比对,比对结果显示与数据库中的可可、开心果、油茶等植物的MSD基因序列相似性较高。表明获得的基因序列为铁观音茶树锰超氧化物歧化酶的基因序列,将其命名为CsMSD(GenBank登录号KP189419)。分析该基因序列及其编码的氨基酸序列(图2),表明其核苷酸序列全长1013bp,开放阅读框(ORF)长693bp,共编码230个氨基酸。

图1 茶树CsMSD基因保守区扩增产物电泳图

图2 茶树CsMSD基因序列及其编码的氨基酸序列
2.2 茶树CsMSD基因编码蛋白的生物信息学分析
茶树CsMSD蛋白基本理化性质分析显示,茶树CsMSD蛋白的原子组成为C1159H1787N301O336S5,分子量为25.47kD,理论等电点pI=6.38,带负电的氨基酸(Asp+Glu)23个,带正电的氨基酸(Arg+Lys)21个,总平均亲水性为-0.269,且性质较稳定,说明该蛋白为稳定的亲水酸性蛋白。分析CsMSD蛋白质的保守结构域(图3),结果表明蛋白质序列中具有保守的Sod_Fe_N和Sod_Fe_C特征结构域。结合CsMSD氨基酸序列中具有MnSOD特有的5个氨基酸残基(图2),说明该蛋白为茶树MnSOD蛋白。

图3 茶树CsMSD蛋白质的保守结构域
茶树CsMSD蛋白亚细胞定位预测结果表明,CsMSD最可能定位于线粒体中。信号肽预测结果显示蛋白质信号肽剪切位置分值均较低,表明茶树CsMSD不存在信号肽。蛋白质在合成后往往需要进行跨区域运输再行使其功能,在跨膜运输过程中需要蛋白质中含有特定的跨膜结构。对CsMSD进行跨膜结构预测(图4),结果显示CsMSD蛋白序列含有一个从内到外的22个氨基酸长度的跨膜结构,位于CsMSD第6-27位氨基酸,分值为739。据此推测CsMSD跨膜结构更倾向于N-端在内的从内到外的跨膜方式。
蛋白质的磷酸化位点往往也是蛋白质的功能区域。对CsMSD进行磷酸化位点分析,结果显示CsMSD存在9个磷酸化位点,其中4个酪氨酸、3个苏氨酸和2个丝氨酸磷酸化位点(图5)。
对CsMSD蛋白的二级结构和三级结构进行预测,结果显示其二级结构主要由37.0%α-螺旋、20.4%延伸链、9.6%β-转角和33.0%无规则卷曲结构构成。二级结构预测结果表明其具有较高程度的α-螺旋,和较低程度的β-转角,这可能与其蛋白质的功能有关。预测CsMSD蛋白质的卷曲螺旋结构(图6)发现,当Windows值为14和21时,CsMSD均可能在蛋白质序列中形成卷曲螺旋结构;而当Windows值为28时,则不存在卷曲螺旋结构。并进一步预测了CsMSD蛋白质的三维结构模型(图7)。
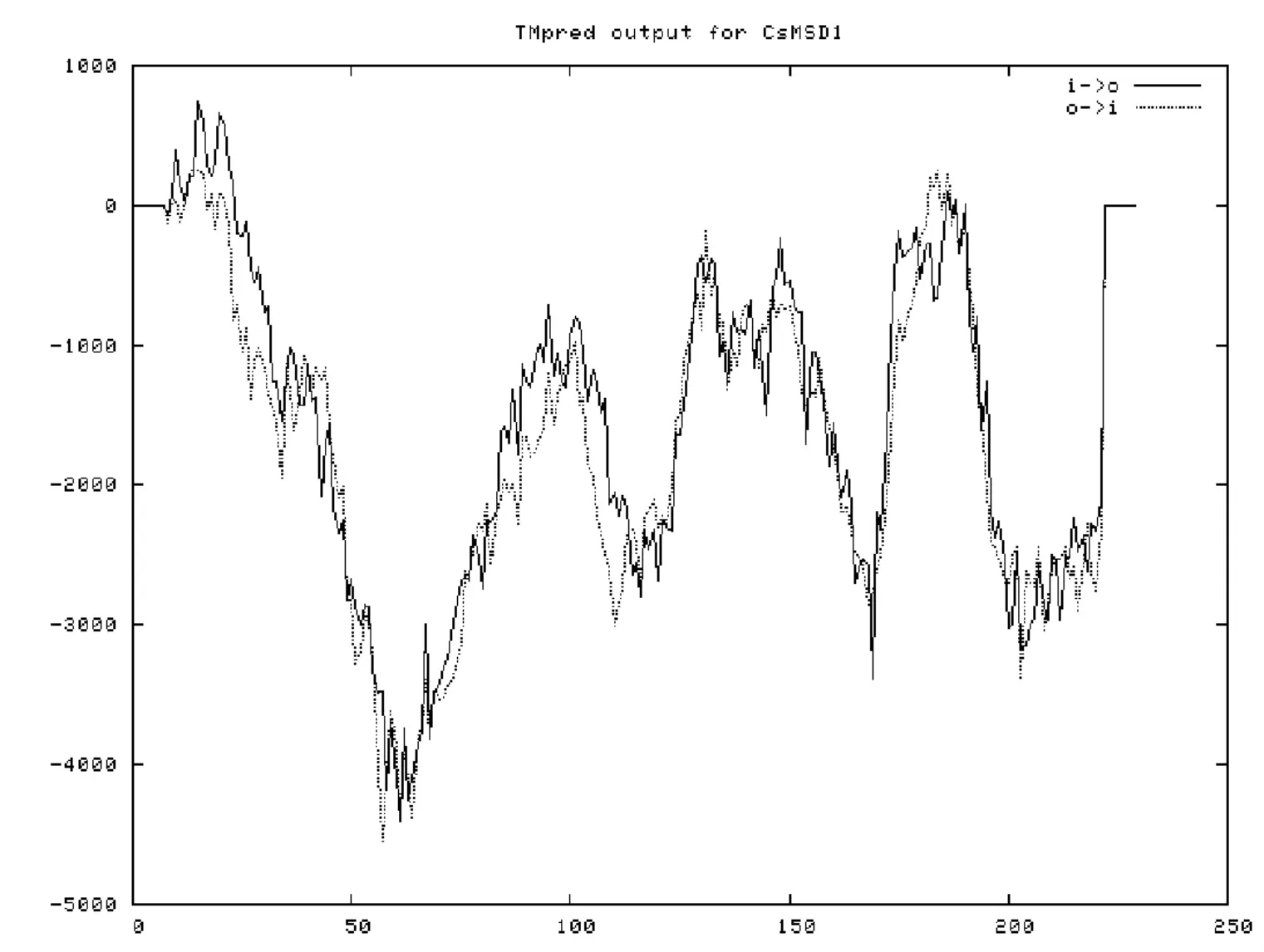
图4 茶树CsMSD蛋白跨膜结构预测

图5 茶树CsMSD蛋白磷酸化位点分析

图6 茶树CsMSD卷曲螺旋结构预测

图7 茶树CsMSD三维结构预测
2.3 茶树CsMSD蛋白质系统进化分析
通过MSD蛋白质序列多重比对分析发现茶树CsMSD与油茶、烟草、可可等植物的MSD氨基酸序列同源性较高。进一步选取具有代表性的植物中已知MSD氨基酸序列,使用邻近归并法构建MSD系统进化树(图8),结果显示茶树MSD氨基酸序列与油茶的MSD亲缘关系和进化距离较近,可能在进化的过程中面临着相近的进化压力。
2.4 茶树CsMSD在不同程度干旱胁迫下的表达情况

图8 MSD系统进化树分析
通过实时荧光定量PCR检测不同程度干旱胁迫情况下铁观音茶树嫩叶中CsMSD基因的相对表达情况(图9),结果显示铁观音茶树中CsMSD基因在轻度和中度干旱胁迫下表达量均轻微上调,相对表达量均为对照组的2倍左右;在重度干旱胁迫下,表达量上调幅度较大,达到对照的6.3倍。在轻度和中度干旱胁迫下CsMSD的表达差异不大,两者之间的相对表达量未达到显著差异水平,但与对照相比均达到显著差异水平;在重度干旱胁迫下,茶树CsMSD基因的相对表达量显著上调,与对照和其他处理间的表达水平差异均达到显著差异水平。推测在重度干旱胁迫下茶树CsMSD对清除叶片细胞中的活性氧发挥着重要作用。茶树CsMSD基因可能在干旱胁迫下通过上调表达来提高超氧化物歧化酶活性,实现调控体内活性氧平衡,参与铁观音茶树应对干旱胁迫。

图9 铁观音茶树CsMSD基因在不同程度干旱胁迫下的表达情况
3 讨论与结论
SOD活性与植物的耐氧化性、耐旱性之间呈正相关,且耐旱性随着耐氧化性增强而增强[11]。而SOD活性受到相应SOD基因的表达调控。植物转基因试验[12,13]发现,过表达 SOD 基因能显著增强转基因植物的SOD活性。MnSOD转基因试验[14-17]结果表明,转基因植株的MnSOD活性和总SOD活性均会发生显著提高;MnSOD转基因高表达植株的抗氧化能力会明显高于对照。柽柳、水稻等作物的转基因试验更直接表明,MnSOD基因具有提高转基因作物抗干旱能力的作用[18,19]。本研究通过实时荧光定量PCR测定发现铁观音茶树叶片中CsMSD的相对表达量在干旱胁迫下显著上调,这与白车轴草中MnSOD基因转录水平的相关研究结果一致[20]。推测在干旱胁迫中铁观音茶树通过调控MnSOD上调表达来提高自身MnSOD的活性,进一步增强自身的抗氧化能力,从而增强自身的耐旱性,达到应对干旱胁迫的目的。
小麦中MnSOD基因研究[21]结果表明,MnSOD与作物对多种非生物胁迫的抗性有关。MnSOD基因在应对NaCl、Na2CO3、NaHCO3和紫外线等其他类型的胁迫中发挥重要作用[22]。银杏中MnSOD基因表达调控研究[23]结果表明,其表达调控还受到ABA、IAA、蔗糖和低温等的诱导。仙客来转基因试验[15]发现,MnSOD能够提高植株对高温胁迫的耐受性。拟南芥MnSOD转基因试验还发现,MnSOD基因除了能提高植株各种非生物胁迫下总SOD活性,还能提高抗氧化酶CAT和POD的活性[24]。据此推测茶树MnSOD不仅能在应对干旱胁迫中发挥重要作用,而且在茶树应对其他种类非生物胁迫中也能发挥积极作用。
亚细胞定位分析显示茶树MnSOD蛋白最有可能定位于线粒体中,与银杏、茄子等[23,24]植物中的结果一致。推测茶树中MnSOD主要负责维持线粒体内的活性氧平衡。而植物体内的活性氧还参与植物器官生长发育的调控,其中SOD能够通过影响植物体内H2O2的含量进而调控木质化次生壁形成和纤维分化等生理过程[25]。水稻中相关试验[26]结果还表明,在水稻叶片的不同发育阶段SOD基因的表达水平有所不同。推测茶树中MnSOD基因在植株相应的生长发育过程中发挥着重要作用。
[1]马旭俊,朱大海.植物超氧化物歧化酶(SOD)的研究进展[J].遗传,2003,02:225-231.
[2]McCord J M,Fridovich I.Superoxide dismutase:an enzymatic function for erythrocuprein (hemocuprein)[J].J Biol Chem,1969,244:6049-6055.
[3]胡根海,喻树迅,范术丽,等.编码棉花胞质铜锌超氧物歧化酶基因的克隆与表达分析[J].中国农业科学,2007,08:1602-1609.
[4]王秋悦,邹亚学,唐家明,等.含锰超氧化物歧化酶基因结构及其转录调控[J].河北科技师范学院学报,2012,04:28-35.
[5]王小萍,王云,唐晓波,等.茶树抗旱相关生理性状的灰色关联度分析[J].茶叶科学,2013,06:512-516.
[6]王小萍,王云,唐晓波,等.干旱胁迫对茶树生理指标的影响[J].西南农业学报,2014,03:1037-1040.
[7]吴芹,张光灿,裴斌,等.3个树种对不同程度土壤干旱的生理生化响应[J].生态学报,2013,12:3648-3656.
[8]赵姗姗,郭玉琼,潘一斌,等.铁观音茶树叶片总 RNA 提取方法研究[J]. 龙岩学院学报,2015,33(2):78-81.
[9]刘玉英,王三根,徐泽,等.不同茶树品种干旱胁迫下抗氧化能力的比较研究[J].中国农学通报,2006,04:264-268.
[10]孙美莲,王云生,杨冬青,等.茶树实时荧光定量PCR分析中内参基因的选择[J].植物学报,2010,45(5):579-587.
[11]汪本勤,陈曦,向成斌.SOD活性增高的拟南芥晚花突变体具有增强的非生物胁迫耐受性[J].植物学通报,2007,05:572-580.
[12]刘明求,刘齐元,丁小维,等.转基因SOD,POD高表达烟草幼苗抗热性研究[J].云南农业大学学报,2005,05:620-623.
[13]刘飞虎,梁雪妮,周玮,等.外源SOD和APX基因在转基因烟草中的表达与遗传[J].植物生理学通讯,2006,05:877-881.
[14]韩利芳,张玉发.烟草MnSOD基因在保定苜蓿中的转化[J].生物技术通报,2004,01:39-42+46.
[15]陈莉,周连霞,马锋旺,等.转MnSOD基因仙客来植株的获得及其对高温胁迫的抗性[J].西北农林科技大学学报(自然科学版),2008,03:155-160.
[16]付畅,张恒,刘关君,等.西伯利亚蓼PsMnSOD基因的耐盐碱性鉴定及其在盐碱胁迫下的表达模式[J].分子植物育种,2009,05:861-866.
[17]杜娟,朱祯,李晚忱.外源超氧化物歧化酶基因MnSOD在玉米中的过量表达及抗逆性的提高[J].植物生理与分子生物学学报,2006,01:57-63.
[18]王丙锋,杨传平,王玉成,等.柽柳MnSOD基因的克隆及功能验证[J].分子植物育种,2007,05:709-714.
[19]Wang F Z,Wang Q B,Kwon S Y,et al.Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase[J].Journal of Plant Physiology,2005,162(4):465-472.
[20]Zhang Y,Li Z,Peng Y,et al.Clones of FeSOD,MDHAR,DHAR Genes from White Clover and Gene Expression Analysis of ROS-Scavenging Enzymes during Abiotic Stress and Hormone Treatments.[J].Molecules,2015,20(11):20939-20954.
[21]Kaouthar F,Ameny F K,Yosra K,et al.Responses of transgenic Arabidopsis,plants and recombinant yeast cells expressing a novel durum wheat manganese superoxide dismutase TdMnSOD,to various abiotic stresses[J].Journal of Plant Physiology,2016,198:56-68.
[22]班巧英,刘桂丰,姜静,等.紫杆柽柳MnSOD基因在非生物胁迫中的作用[J].植物生理学通讯,2008,02:211-214.
[23]程华,李琳玲,许锋,等.银杏锰型超氧化物歧化酶GbMnSOD基因的克隆与表达[J].园艺学报,2009,09:1283-1290.
[24]徐龙,陈火英,蒋明敏,等.茄子MnSOD基因的克隆及表达分析[J].植物生理学报,2016,10:1537-1545.
[25]林植芳,刘楠.活性氧调控植物生长发育的研究进展[J].植物学报,2012,01:74-86.
[26]Nath K,KumarS,PoudyalR S,etal.Developmentalstage-dependentdifferentialgene expression ofsuperoxide dismutase isoenzymes and their localization and physical interaction network in rice (Oryza sativa,L.)[J].Genes&Genomics,2014,36(1):1-11.
Cloning and Expression Analysis of Manganese Superoxide Dismutase Gene inCamellia Sinensis
WANG Zhong1,LAI Zhong-xiong2,IN Qi-fan1,GUO Yu-qiong2,JCHANG Xiao-jun2
(1.School of Food and Bioengineering,Hezhou University,Hezhou Guangxi 542899;2.Institute of Horticultural Biotechnology,Fujian Agriculture and Forestry University,Fuzhou Fujian 350002)
The Manganese Superoxide Dismutase Gene,which plays important roles in response to drought stress,was cloned from Camellia sinensis cv.Tieguanyin.The gene sequence(GenBank accession number,KP189419)was found to be 1013bp,containing a 693bp open reading frame which encodes 230 amino acids.The deduced protein molecular weight was 25.47 kD and its theoretical isoelectric point was 6.38.The bioinformatics revealed that the protein did have 9 phosphorylation sites,and that its subcellular cells located in mitochondria.The protein sequences had conserved functional domains related to manganese superoxide dismutase.QPCR analysis indicated that the expression level of CsMSD,when the tea trees was under varied drought stresses,was significantly higher than the control group,which suggested that CsMSD might play important roles during the processes of tea plant responding to the drought stresses.
Camellia sinensis cv.Tieguanyin;CsMSD;gene cloning;drought stress;qPCR
S571.1
A
1673—8861(2017)02—0149—08
[责任编辑]刘丽英
2017-04-06
王仲(1991-),男,湖南衡阳人,硕士。主要研究方向:茶类资源挖掘与创新利用。
郭玉琼(1974-),女,福建永春人,福建农林大学副教授,博士。主要研究方向:茶树生物技术与茶叶生物化学。
福建省2015年重大科技专项项目(2015NZ0002-1)。
