基于SSR标记的木荷核心种质构建*
2017-07-18杨汉波王帮顺徐肇友周志春
杨汉波 张 蕊 王帮顺 徐肇友 周志春
(1.中国林业科学研究院亚热带林业研究所 浙江省林木育种技术研究重点实验室 杭州 311400;2.浙江省龙泉市林业科学研究院 龙泉 323700)
基于SSR标记的木荷核心种质构建*
杨汉波1张 蕊1王帮顺2徐肇友2周志春1
(1.中国林业科学研究院亚热带林业研究所 浙江省林木育种技术研究重点实验室 杭州 311400;2.浙江省龙泉市林业科学研究院 龙泉 323700)
【目的】 通过对比分析与评价以确定木荷核心种质构建的最适取样策略和比例,并构建木荷核心种质; 在此基础上,进一步建立核心种质分子身份信息,为木荷种质资源的深入研究和加强利用、发掘优异基因资源提供理论依据和核心材料,同时也可为其他多年生木本植物核心种质的构建提供参考。【方法】 利用13对SSR引物,以来自7个省(市)29个地区的754份木荷种质资源为材料,利用M策略(M)、随机取样法(R)、遗传多样性最大化法(SAGD)和等位基因最大化法(SANA)分别构建核心种质。采用等位基因数(Na)、有效等位基因数(Ne)和Shannon’s信息指数(I)等遗传多样性指标进行比较分析来确定最适合的构建方法。【结果】 13对SSR引物共检测到128个等位基因(Na),平均有效等位基因数(Ne)为3.47,Shannon’s信息指数(I)为1.39,表明木荷种质资源具有丰富的遗传多样性。SANA、SAGD和M策略构建的核心种质均优于R策略。SANA和SAGD法抽取的核心种质对原有种质均具有较好的代表性,但等位基因保留率较低。M策略构建的核心种质等位基因(Na)保留比例明显高于其他3种策略所构建的核心种质。根据遗传多样性参数综合考虑,同时考虑抽样数量,M策略构建的核心种质能以最小的取样量、最大程度地保留原有种质的遗传多样性,为最优的取样策略。采用主坐标分析法显示,M策略构建的核心种质能够较全面地代表木荷种质资源的遗传多样性,利用该策略得到的115份木荷核心种质,保留了原有种质15.3%的种质材料,等位基因数(Na)、有效等位基因数(Ne)和Shannon’s信息指数(I)的保留率分别达到93.8%,115.6%和109.9%。依据13对SSR引物的扩增数据,经过多态性谱带的有序编码转换,构建了115份木荷核心种质的特异分子身份信息,置信概率达到99.99%,具有有效性和唯一性。【结论】 M策略是较适宜的构建木荷核心种质的方法,构建的115份核心种质能最大程度代表木荷种质资源的遗传多样性,同时,本研究所采用的方法对其他多年生木本植物核心种质的构建具有重要的参考价值。
木荷; SSR标记; 核心种质; 分子身份信息
植物种质资源是育种工作的重要物质基础,是决定育种效果的关键,世界各国都非常注重对种质资源的调查、搜集、评价、保存和利用(Escribanoetal., 2008)。然而,大量的种质资源在为植物遗传改良和品种选育提供了丰富遗传基础的同时,也给种质资源的搜集保存及研究利用带来了困难(Escribanoetal., 2008)。针对这一问题,Frankel等(1984)首次提出核心种质的概念,Brown(1989)将其进一步发展,核心种质是指以最小数量的种质资源和遗传重复最大程度地代表整个种质资源的遗传多样性,是资源深入研究、优良基因挖掘和新技术应用的核心子集,能够提高种质资源的有效利用率。核心种质的提出为种质资源的有效保护与深入研究、利用开辟了新的途径,发达国家和农业研究中心十分注重对核心种质的研究,使其逐渐成为国际种质资源研究的热点(Belajetal., 2012; 刘娟等, 2015)。利用分子标记的方法,Miyamoto等(2015)从3 203份日本柳杉(Cryptomeriajaponica)优树中选出了349份核心种质; Belaj等(2012)建立了总体比例为10%的橄榄(Canariumalbum)核心种质; 王丽侠等(2014)建立了数目为157份的扁豆(Lablabpurpureus)核心种质。我国从1994年开始提出构建核心种质,现今已经在多种农作物上建立了核心种质,如水稻(Oryzasativa)(Zhangetal., 2011)、小麦(Triticumaestivum)(董玉琛等, 2003)、苎麻(Boehmerianivea)(栾明宝等, 2010)等。而林木核心种质构建工作相对滞后,仅对杏(Armeniacavulgaris)(刘娟等, 2015)、新疆野苹果(Malussieversii)(张春雨等, 2009)、白桦(Betulaplatyphylla)(魏志刚等, 2009)、欧洲黑杨(Populusnigra)(曾宪君等, 2014)等少数树种进行了初步研究。林木等多年生木本植物树体庞大,且资源保存通常以田间保存为主,占地面积大,管理费用高,因此,建立林木核心种质是十分必要和迫切的。
木荷(Schimasuperba)为山茶科(Theaceae)木荷属(Schima)常绿大乔木,是我国东部湿润亚热带常绿阔叶林的主要建群树种,天然散布于东经105°以东、北纬31°以南的广阔地区,是我国南方各省区的珍贵优质阔叶用材和当家生物防火树种,在商品用材林和生态防火林建设中占有重要地位(倪健, 1996; 张萍等, 2004; 张蕊等, 2013)。项目组自2001年开始,先后开展了木荷育种与培育技术的研究,在木荷主产区选择优树1 000余株,已成功嫁接保存了754个优树无性系,为高抗优质新品种选育和种子园营建等木荷遗传研究和育种工作提供了大批量的优质材料。如何从丰富的木荷种质资源中快速、准确地鉴定出木荷育种上迫切需要的优异基因,是木荷育种工作所急需解决的一个重大问题。本研究在项目组前期研究 (张萍等, 2006; 辛娜娜等, 2014; 2015; 杨汉波等, 2016) 的基础上,借鉴其他多年生木本植物核心种质构建的方法,利用SSR分子标记,以754份木荷种质资源为材料,通过对比分析与评价不同构建策略和取样比例构建的核心种质,确定木荷核心种质构建的最适取样策略和比例,并构建木荷核心种质; 在此基础上,进一步建立核心种质分子身份信息,为木荷种质资源的深入研究和加强利用、发掘优异基因资源提供理论依据和核心材料,同时也可为其他多年生木本植物核心种质的构建提供参考。
1 材料与方法
1.1 试验材料
试材取自浙江省龙泉市林业科学研究院上圩基地的木荷基因库(28°03′N, 119°06′E,面积为6.7 hm2,海拔200~300 m,相对湿度79%,年均降雨量1 664.8~1 706.2 mm)。2010年至今,在浙江、福建、江西等木荷主产区选择优树1 000余株,嫁接保存754株。选优林分要求林龄20年、面积1.0 hm2以上,以木荷为主的优良天然林或起源明确的人工林; 优树选择条件为树体高大,干形通直圆满,生长量明显高于附近3~5株同龄优势木等。以当地1~2年生木荷容器苗为砧木,在3—4月份选用带有休眠芽的穗条,采用切接的方法嫁接优树无性系,在圃地集中培育2年生容器嫁接苗。然后根据林木种质资源异地保存库营建要求(株行距为4 m×4 m)将无性系嫁接苗移植至基因库相应位置上。取样种质为浙江(109份)、福建(350份)、江西(168份)、湖南(75份)、广东(32份)、贵州(12份)和重庆(8份)7个省(市)的754份木荷种质资源。2015年6月,对来自上述7个省(市)资源群体全部单株采集其顶端新发枝条上的新鲜嫩叶,将其放入液氮中带回实验室,置于-80 ℃冰箱中保存。
1.2 基因组DNA提取及SSR扩增
采用植物DNA快速提取试剂盒提取木荷基因组DNA,1.0%琼脂糖凝胶电泳检测DNA纯度和完整性,NanoDrop-2000超微量分光光度计测定260 mm/280 mm处的光吸收值来检测其浓度,最后稀释到20 ng·μL-1,-20 ℃保存备用。本研究选用13对条带清晰、多态性强的SSR引物进行PCR扩增(Niuetal., 2012; 辛娜娜等, 2015)。PCR扩增反应在TaKaRa PCR Thermal Cycler Dice(TaKaRa)上进行。PCR扩增反应体系25 μL: 含12.5 μL 2×Taq Plus Master Mix,1 μL 10 μmol·L-1上下游引物和2.5 μL基因组DNA。PCR扩增程序为: 95 ℃ 5 min; 94 ℃ 30 s,53 ℃ 30 s,72 ℃ 45 s(35个循环); 72 ℃延伸10 min。扩增产物在Qsep100TM全自动核酸蛋白自动分析系统上进行电泳分离检测,Qsep100TM能以1~4 bp的分辨率高效区分20~20 000 bp DNA片段 (http://www.sciencemag.org/content/348/6241/1383.1.full)。
1.3 核心种质构建
Frankel等(1984)和Brown(1989)提出可以利用地理来源和表型数据构建核心种质。先按照地理来源分组、后取样的策略也已被许多研究证明是合理有效的(李自超等, 1999; 崔艳华等, 2003; 赵冰等, 2007)。因此,本研究先将754份木荷种质资源按地理来源分为7组,再按照下述方法分别对各组种质进行抽样分别构建核心种质,最后将各组抽取的核心种质进行合并构成全部木荷种质资源的核心种质。
1)M策略(Maximization strategy, M strategy):利用一系列遗传标记的位点来鉴定同时具有低冗余和高丰度等位变异的种质材料,并确定如何在组内合理地分配这些种质材料,分配原则主要建立在一种假设上,即最大化核心种质中的标记位点,就相当于最大化目标位点(Schoenetal., 1995)。通过Core Finder软件(Ciprianietal., 2010)以等位基因数最大化为原则对SSR数据进行分析抽取核心种质。
2)随机取样法(Random sampling, R):总体取样比例设定为10%,15%, 20%,25%和30%,根据随机取样策略构建核心种质(张春雨等, 2009)。
3)采用PowerMarker version 3.25软件(Liuetal., 2005),选取的总体取样比例同2),根据模拟退火算法(simulated annealing algorithm, SA)以等位基因最大化(maximizing allelic richness, SANA)为标准构建核心种质。
4)采用PowerMarker version 3.25软件(Liuetal., 2005),选取的总体取样比例同2),根据模拟退火算法(simulated annealing algorithm, SA)以遗传多样性最大化(maximizing genetic diversity, SAGD) 为标准构建核心种质。
1.4 数据分析
对SSR检测结果进行峰图分析及等位基因的读取,然后利用GenAlEx6.5软件(Peakalletal., 2012)计算等位基因数(Na)、有效等位基因数(Ne)、期望杂合都(He)、观测杂合度(Ho)及Shannon’s信息指数(I),用这些指标来评价核心种质、原有种质和保留种质的遗传多样性,并通过t检验对核心种质与原有种质各遗传多样性指标进行差异显著性检测(Escribanoetal., 2008)。同时运用主坐标分析法(PCoA)对构建的核心种质进行确认(Wangetal., 2007)。将毛细管电泳谱带转化为数字指纹图谱并以此构建木荷核心种质的分子身份信息,依据指纹图谱概率公式P=1/2n(n为等位基因的数目)计算其置信概率(唐源江等, 2015)。
2 结果与分析
2.1 SSR分子标记遗传多样性
采用SSR标记对754份木荷种质资源的遗传多样性进行分析(表1),13对SSR引物共检测到128个等位基因(Na),有效等位基因数(Ne)为3.47,每对引物检测到的等位基因数(Na)的变化范围为4(ss01)~15(ss16),平均为9.85。各引物的期望杂合度(He)变化范围0.54(ss30)~0.83(ss32),平均为0.68。Shannon’s信息指数(I)变化范围0.91(ss01)~1.92(ss32),平均为1.39。结果说明木荷种质资源具有丰富的遗传多样性。
2.2 不同方法构建的核心种质遗传多样性指标的比较
不同方法构建核心种质的遗传多样性指标见表2。结果显示,随着取样比例的增加,各级候选核心种质等位基因数(Na)保留率呈逐渐上升的趋势。各候选核心种质有效等位基因数(Ne)保留率均在95%以上,其中M策略以15.3%取样比例构建的核心种质Ne保留率最高,为115.6%,其次是SANA(10%),为100.9%。Shannon’s信息指数(I)仅M策略构建的候选核心种质高于原有种质,其余均低于原有种质,但其保留率均在97%以上。SANA和R在10%、15%和20%抽样比例下的核心种质与原有种质Nat检验存在显著差异,SAGD在10%和20%抽样比例下的核心种质与原有种质Nat检验也存在显著差异。各候选种质的Shannon’s信息指数(I)、有效等位基因数(Ne)、观测杂合度(Ho)和期望杂合度(He)与原有种质t检验无显著差异。从等位基因保留数目和比例看,M策略构建的核心种质等位基因保留比例为93.8%,明显高于其他3种策略所构建的核心种质。根据这5个遗传参数综合考虑,同时考虑抽样数量,选择M策略抽取115份(15.3%)资源时,构建的木荷核心种质能够以最小的种质份数最大程度地代表整个木荷种质资源的遗传多样性。利用M策略构建木荷核心种质中包含湖南15份、江西24份、福建27份、广东13份、浙江21份、贵州9份及重庆6份共115份种质材料。
采用基于位点优先的M策略构建的木荷核心种质保留了原有种质15.3%的材料,等位基因数(Na)、有效等位基因数(Ne)、观测杂合度(Ho)、期望杂合度(He)、Shannon’s信息指数(I)等遗传多样性指标的保留率分别为93.8%,115.6%%,98.8%,104.7%和109.9%;t检验结果表明,利用M策略构建的木荷核心种质和原有种质的遗传多样性指数差异不显著(表3)。这说明利用M策略构建的核心种质对原有种质进行了有选择性的选取,剔除了冗余种质后,能够很好地体现原有木荷种质资源的遗传多样性。由此认为本文利用M策略构建的木荷核心种质在遗传多样性上能很好地代表原木荷种质资源。同时,对核心种质和保留种质的遗传多样性指标进行比较分析(表3),t检验结果表明在概率0.05水平上,核心种质的Ne、I和He均显著高于保留种质。因此,应优先考虑使用木荷核心种质开展高效的木荷育种工作。采用主坐标分析法(PCoA)对构建的木荷核心种质和整个木荷种质资源进行分析,以确定其代表性(图1),发现木荷核心种质在整个木荷种质资源的主坐标图内均匀分布,表明采用M策略(Core Finder软件)构建的木荷核心种质具有很好的代表性。

表1 木荷754份种质资源遗传多样性参数①Tab. 1 The genetic diversity parameters ofSchima superba germplasm resources
①Na: 等位基因数;Ne: 有效等位基因数;I: Shannon’s信息指数;Ho: 观测杂合度;He: 期望杂合度。下同。Na: Number of different alleles;Ne: Number of effective alleles;I: Shannon’s information index;Ho: Observed heterozygosity;He: Expected heterozygosity. The same below.
2.4 核心种质分子身份信息构建
用13对SSR引物扩增的全部128条多态性谱带构建了115份木荷核心种质的分子身份信息(图2),其带型全部相同的概率为2.9×10-39,置信概率达到99.99%。每份核心种质材料都有唯一分子身份信息,可以比较容易地将115份木荷核心种质材料相互区分鉴别出来。

表2 不同策略构建的核心种质遗传多样性指标比较①Tab. 2 Comparisons of genetic diversity index of core collections constructed by different tactics
①M: M策略; R: 随机取样法; SAGD: 遗传多样性最大化法; SANA: 等位基因最大化法。*表示在α=0.05水平下核心种质与原有种质各项指标的统计检验的显著性。M: Maximization strategy; R: Random sampling; SAGD: Simulated annealing algorithm maximizing the genetic diversity; SANA: Simulated annealing algorithm maximizing the number of alleles. * Significant at 0.05 probability of each index between core collection and original collection.

表3 M策略构建的核心种质、保留种质和原有种质遗传多样性对比①Tab. 3 Genetic diversity comparison and t-test between original collection, reserve collection and core collection constructed by M strategy
①括号内为保留率。t1值表示核心种质与原有种质各遗传参数t检验值,t2表示核心种质与保留种质各遗传参数t检验值。*表示在0.05水平下核心种质与保留种质间存在显著差异。The contents in the brackets are the percentage of reserve.t1means thetvalue of genetic diversity parameters between core collection and initial collection, andt2means thetvalue of genetic diversity parameters between core collection and reserve collection. * means significant difference at 0.05 probability between core collection and reserve collection.
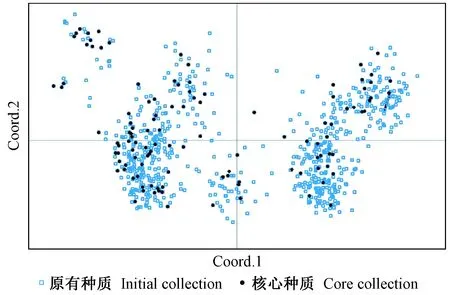
图1 M策略构建的核心种质与原有种质的主坐标Fig.1 Principal coordinates plots of core collection constructed by M strategy and initial collection

图2 13个SSR标记构建的115份木荷核心种质分子身份信息Fig.2 Molecular identity of 115 core collection set up by SSR markers
3 讨论
目前构建核心种质主要基于形态标记和分子标记分析2类(王建成等, 2008)。对树体高大的多年生林木而言,资源的搜集与保存较为困难,要得到可靠的表型数据需要较长的年限(王红霞等, 2013)。分子标记以DNA分子多态性为基础,一般不具有生物功能活性,不会或极少受植物生长期和外界环境的影响,较形态标记更适用于构建核心种质和进行遗传多样性评价(刘新龙等, 2014)。SSR是共显性标记,提供的等位基因信息较其他分子标记更为丰富,采用SSR数据构建的核心种质较其他标记具有较大的优势(张君玉, 2012)。因此,本研究采用13对多态性SSR引物,采用4种不同的鉴别和筛选方法,有效地得到115份木荷核心种质材料,具有重要的理论和实践意义。核心种质的构建中样本量的大小是衡量核心种质是否有效的重要因子之一(杨美等, 2011)。由于物种的特性,核心种质的取样比例应以所研究植物的遗传多样性和遗传结构而定(李自超等, 2003)。迄今为止,在国内外所构建的不同植物核心种质其所占比例一般为原有种质的5%~40%,大多数在5%~15%(Wangetal., 2007; Escribanoetal., 2008)。王萱等(2016)分析了180份银杏(Ginkgobiloba)古树遗传多样性,获得了63份核心种质,占原有种质的35%。Liang等(2014)利用SSR标记获得了55份苹果(Malusdomestica)核心种质,占原有种质的13.2%。Wang等(2014)利用EST-SSR标记建立了占原有材料22.9%(22份种质)的荔枝(Litchichinensis)核心种质。这些结果均表明取样比例应视种质资源的数量和遗传多样性而定。M策略基于等位基因(Na)的最大化,同时兼顾遗传多样性,经Core Finder软件自动生成合理的取样比例,获得的取样比例最为科学、可靠(郭大龙等, 2012)。在本研究中,木荷种质资源共有754份,通过对不同取样比例构建的核心种质进行比较分析,结果表明以15.3%取样比例(M策略)构建的木荷核心种质能够最大程度地代表整个木荷种质资源的遗传多样性。
取样策略是影响构建的核心种质是否有效的另一重要因子(刘娟等, 2015)。目前多数研究者采用分层分组和聚类的方法构建核心种质,但不同的分组和聚类方法构建的核心种质存在较大的差异(Wangetal., 2007; Ciprianietal., 2010)。因此,应对取样策略进行优化以获得最具代表性的核心种质。植物基因多样性是决定其性状的遗传因子,而位点和等位基因频率是种质资源基因多样性的基础,构建的核心种质应以最小的取样比例最大限度地保留原有种质的等位基因多样性(刘娟等, 2015)。本研究中,基于等位基因和遗传多样性优先的取样策略(M、SANA和SAGD策略)构建的木荷核心种质其保留的等位基因数明显多于随机取样策略(R策略),遗传多样性指标也均高于随机取样策略,与前人研究结果 (刘娟等, 2015) 基本一致。因此,基于等位基因和遗传多样性优先的M、SANA和SAGD取样策略构建的木荷核心种质优于R取样策略。M策略构建的核心种质以15.3%的取样比例代表了原有种质93.8%的等位基因多样性,明显优于SANA和SAGD取样策略,也显著高于Frankel等(1984)提出的核心种质应以10%的取样比例代表原有种质70%以上的遗传多样性的标准。依据地理来源对种质资源进行分组取样是取得高代表性核心种质的一种较好的分组策略(Liuetal., 2005)。本研究中,首先根据地理来源对木荷种质资源进行分组取样,在此基础上通过4种不同取样策略(M、R、SANA和SAGD)构建木荷核心种质,最后通过遗传多样性指标进行评价,结果表明先按来源地分组,再利用M策略(Core Finder软件)分组取样是构建木荷核心种质的最优取样策略,构建的木荷核心种质能有效代表整个木荷种质资源的地理分布和遗传多样性,说明保证原有种质不同地理来源种质的相应比例是获得高遗传多样性代表性核心种质的最好方法,也表明在核心种质取样时,应尽量包含所有来源地的种质资源。本研究最终以M策略得到115份木荷核心种质,分别为广东13份(40.6%)、湖南15份(20%)、贵州9份(75%)、江西24份(14.3%)、浙江21份(19.3%)、福建27份(7.7%)和重庆6份(75%); 福建省的种质资源最多,但抽取的核心种质反而较少,这可能与该地区种质资源纯合度较高、亲缘关系较近有关(明军等, 2005),也与李自超等(2003)认为核心种质的取样数量应视种质资源的数量和遗传多样性而定,不能格式化与统一化的观点相符合。
尽管构建的木荷核心种质能够最大程度地代表整个木荷种质资源的遗传多样性,但保留种质仍不能淘汰,以便从保留种质中寻找育种家们需要而在核心种质中无法找到的性状(Ciprianietal., 2010)。对所代表的基因库中种质资源遗传多样性的代表程度是评价核心种质有效性的根本指标; 然而基因库中的材料常常不能完全代表一个种的全部遗传多样性,解决这个问题最有效的方法是使核心种质的大小及内容随时间而变化(明军等, 2005)。本研究原有种质材料比较多,包含中国7个省(市)的754份木荷种质资源,但部分地区的种质资源相对较少(如重庆市、贵州省收集到的种质资源仅分别为8个和12个),因此,为保证木荷丰富的物种多样性,需不断地补充新搜集的木荷优树种质进入木荷核心种质中。本研究对木荷核心种质的研究尚属首次尝试,由于标记数量的限制及表型特征数据的缺乏,在今后的研究中仍需加以补充和完善。
通过DNA分子标记来鉴别品种资源是准确而有效的方法,这在大量的种质资源研究中得到应用(唐源江等, 2015; 李婷婷等, 2016)。由于具有特征条带的材料较少,可以利用2对以上的引物组合来区分材料,若2对引物组合不能完全区分开所有材料时,则可以增加引物组合数直至区分开所有种质材料(唐源江等, 2015)。SSR为共显性标记,是鉴定种质资源的理想标记之一(Zhangetal., 2012),本研究选用13对SSR引物进行组合,首次构建了系统的木荷核心种质分子鉴定、图谱及数字化分子身份信息。利用SSR指纹图谱库构建的115份核心种质分子身份信息,其置信概率达99.99%,说明利用构建的分子身份信息足以将115份木荷核心种质区分开来。有限的引物只能适用于一定数量的样本群体,本研究木荷核心种质达到了115份,随着核心种质数量的增加,就需要增加更多合适的引物进行鉴定(Heckenbergeretal., 2002)。在今后的工作中,应将形态信息与分子数据结合在一起形成更加完整的木荷核心种质指纹图谱,使育种工作者能够借助其分子身份信息更有针对性地开展研究,以大大缩短种质鉴定时间。
(1) 随车辆的前移,结合梁跨中挠度曲线以波动式变化且挠度值逐渐增大,当车辆行驶到试验梁跨中附近时跨中挠度值最大;随剪力连接度的减小,部分连接结合梁的刚度降低,在相同车重和车速下,其跨中最大动挠度值要高于完全连接结合梁。
4 结论
根据SSR标记数据构建木荷核心种质,采用Corefinder软件以等位基因最大化为原则的M策略优于随机取样法(R)、根据模拟退火算法的等位基因最大化(SANA)和遗传多样性最大化(SAGD)为标准的取样策略,是构建木荷核心种质的较适宜方法。按地理来源将754份木荷种质资源划分为7组,在各组内采用M策略,构建了含有115份材料的木荷核心种质,保留了原有种质15.3%的种质材料,等位基因数、有效等位基因数和Shannon’s信息指数的保留率分别达到93.8%、115.6%和109.9%,表明本研究建立的核心种质是有效的。并以13对SSR引物的扩增数据,构建了具有有效性和唯一性的115份木荷核心种质的特异分子身份信息。同时本研究所采用的方法对其他多年生木本植物核心种质的构建具有重要的参考价值。
崔艳华, 邱丽娟, 常汝镇, 等. 2003. 植物核心种质研究进展. 植物遗传资源学报, 4(3): 279-284.
(Cui Y H, Qiu L J, Chang R Z,etal. 2003. Advances in research on core collection of plant germplasm resources. Journal of Plant Genetic Resources, 4(3): 279-284. [in Chinese])
董玉琛, 曹永生, 张学勇, 等. 2003. 中国普通小麦初选核心种质的产生. 植物遗传资源学报, 4(1): 1-8.
(Dong Y C, Cao Y S, Zhang X Y,etal. 2003. Establishment of candidate core collections in Chinese common wheat germplasm. Journal of Plant Genetic Resources, 4(1): 1-8. [in Chinese].)
郭大龙, 刘崇怀, 张君玉, 等. 2012. 葡萄核心种质的构建. 中国农业科学, 45(6): 1135-1143.
(Guo D L, Liu C H, Zhang J Y,etal. 2012. Construction of grape core collection. Scientia Agricultura Sinica, 45(6): 1135-1143. [in Chinese])
李婷婷, 朱锦茹, 邱 帅, 等. 2016. 基于CE-SRAP标记的榉树优良单株指纹图谱构建. 植物遗传资源学报, 17(1): 169-176.
(Li T T, Zhu J R, Qiu S,etal. 2016. Fingerprinting construction for superior individuals ofZelkovaschneiderianabased on CE-SRAP. Journal of Plant Genetic Resources, 17(1):169-176. [in Chinese])
李自超,张洪亮. 1999. 植物遗传资源核心种质研究现状与展望. 中国农业大学学报, 4(5): 51-62.
(Li Z C, Zhang H L. 1999. Status and prospects of core collection in plant germplasm resource. Journal of China Agricultural University, 4(5): 51-62.[in Chinese])
李自超, 张洪亮, 曹永生, 等. 2003. 中国地方稻种资源初级核心种质取样策略研究. 作物学报, 29(1): 20-24.
(Li Z C, Zhang H L, Cao Y S,etal. 2003. Studies on the sampling strategy for primary core collection of Chinese ingenious rice. Acta Agronomica Sinica, 29(1): 20-24. [in Chinese])
刘 娟, 廖 康, 曼苏尔·那斯尔, 等. 2015. 利用ISSR分子标记构建南疆杏种质资源核心种质. 果树学报, 32 (3): 374-384.
(Liu J, Liao K, Mansur N,etal. 2015. Core-germplasm construction of apricot collections in south of Xinjiang by ISSR molecular markers. Journal of Fruit Science, 32(3): 374-384. [in Chinese])刘新龙, 刘洪博, 马 丽, 等. 2014. 利用分子标记数据逐步聚类取样构建甘蔗杂交品种核心种质库. 作物学报, 40(11): 1885-1894.
(Liu X L, Liu H B, Ma L,etal. 2014. Construction of sugarcane hybrids core collection by using stepwise clustering sampling approach with molecular marker data. Acta Agronomica Sinica, 40(11): 1885-1894. [in Chinese])
栾明宝, 陈建华, 许 英, 等. 2010. 苎麻核心种质构建方法. 作物学报, 36(12): 2099-2106.
(Luan M B, Chen J H, Xu Y,etal. 2010. Method of establishing ramie core collection. Acta Agronomica Sinica, 36(12): 2099-2106. [in Chinese])
明 军, 张启翔, 兰彦平. 2005. 梅花品种资源核心种质构建. 北京林业大学学报, 27(2): 65-69.
(Ming J, Zhang Q X, Lan Y P. 2005. Core collection ofPrunusmumeSieb. et Zucc. Journal of Beijing Forestry University, 27(2): 65-69. [in Chinese])
倪 健. 1996. 中国木荷及木荷林的地理分布与气候的关系. 植物资源与环境学报, 5(3): 28-34.
(Ni J. Relationship between geographical distribution ofSchimasuperba, its forest and climate in China. Journal of Plant Resources and Environment, 5(3): 28-34. [in Chinese])
唐源江, 曹雯静, 吴坤林. 2015. 基于SRAP标记的国兰种质资源遗传多样性分析及分子身份证构建. 中国农业科学, 48(9): 1795-1806.
(Tang Y J, Cao W J, Wu K L. 2015. Genetic diversity analysis and molecular identification card construction of Chinese cymbidium germplasms based on SRAP markers. Scientia Agricultura Sinica, 48(9): 1795-1806.[in Chinese])
王红霞, 赵书岗, 高 仪, 等. 2013. 基于AFLP分子标记的核桃核心种质的构建. 中国农业科学, 46(23): 4985-4995.
(Wang H X, Zhao S G, Gao Y,etal. 2013. A construction of the core-collection ofJuglansregiaL. based on AFLP molecular markers. Scientia Agricultura Sinica, 46(23): 4985-4995. [in Chinese])
王建成, 胡 晋, 黄歆贤, 等. 2008. 植物遗传资源核心种质新概念与应用进展. 种子, 27 (5): 47-50.
(Wang J C, Hu J, Huang X X,etal. 2008. New concept and application on core collection of plant germplasm resources. Seed, 27 (5): 47-50. [in Chinese])
王丽侠, 程须珍, 王素华. 2014. 中国饭豆种质资源遗传多样性及核心种质构建. 植物遗传资源学报, 15(2): 242-247.
(Wang L X, Cheng X Z, Wang S H. 2014.Genetic diversity analysis and a core collection construction of rice bean (Vignaumbellata) in China. Journal of Plant Genetic Resources, 15(2): 242-247. [in Chinese])
王 萱, 刘晓静, 邢世岩, 等. 2016. 中国部分古银杏资源遗传多样性的AFLP分析及核心种质的构建. 园艺学报, 43(2): 249-260.
(Wang X, Liu X J, Xin S Y,etal. 2016. AFLP analysis of genetic diversity and a construction of the core collection of partial ancient ginkgo trees in China. Acta Horticulturae Sinica, 43(2): 249-260. [in Chinese])
魏志刚, 高玉池, 刘桂丰, 等. 2009. 白桦核心种质初步构建. 林业科学, 45(10): 74-80.
(Wei Z G, Gao Y C, Liu G F,etal. 2009. Preliminary construction of core collection ofBetulaplatyphyllagermplasm. Scientia Silvae Sinicae, 45(10): 74-80. [in Chinese])
辛娜娜, 张 蕊, 范辉华, 等. 2014.5年生木荷生长和形质性状的家系变异和选择. 林业科学研究, 27(3): 316-322.
(Xin N N, Zhang R, Fan H H,etal. 2014. Family variation and selection of growth and quality characteristics of 5-year-oldSchimasuperbaseedlings. Forest Research, 27(3): 316-322. [in Chinese])
辛娜娜, 张 蕊, 徐肇友, 等. 2015. 木荷1代育种群体遗传多样性分析. 林业科学研究, 28(3): 332-338.
(Xin N N, Zhang R, Xu Z Y,etal. 2015. Genetic diversity among breeding parents ofSchimasuperbarevealed by SSR. Forest Research, 28(3):332-338. [in Chinese])
杨 美, 付 杰, 向巧彦, 等. 2011. 利用AFLP分子标记技术构建花莲核心种质资源. 中国农业科学, 44 (15): 3193-3205.
(Yang M, Fu J, Xiang Q Y,etal. 2011. The core-collection construction of flower lotus based on AFLP molecular markers. Scientia Agricultura Sinica, 44 (15): 3193-3205. [in Chinese])
杨汉波,张 蕊,周志春. 2016. 木荷种子园的遗传多样性和交配系统. 林业科学, 52(12): 66-73.
(Yang H B, Zhang R,Zhou Z C. 2016. Genetic diversity and mating system in a seed orchard ofSchimasuperba. Scientia Silvae Sinicae, 52(12): 66-73. [in Chinese])
曾宪君, 李 丹, 胡彦鹏, 等. 2014. 欧洲黑杨优质核心种质库的初步构建. 林业科学, 50(9): 51-58.
(Zeng X J, Li D, Hu Y P,etal. 2014. A preliminary study on construction of high-quality core collection ofPopulusnigra. Scientia Silvae Sinicae, 50(9): 51-58. [in Chinese])
张春雨, 陈学森, 张艳敏, 等. 2009. 采用分子标记构建新疆野苹果核心种质的方法. 中国农业科学, 42(2): 597-604.
(Zhang C Y, Chen X S, Zhang Y M,etal. 2009. A method for constructing core collection ofMalussieversiiusing molecular markers. Scientia Agricultura Sinica, 42(2): 597-604. [in Chinese])
张君玉. 2012. 利用SSR和SRAP分子标记构建葡萄核心种质. 洛阳: 河南科技大学硕士学位论文.
(Zhang J Y. 2012. Construction of core collections for grape germplasm using SSR and SRAP molecular markers. Luoyang: MS thesis of Henan University of Science and Technology.[in Chinese])
张 萍, 金国庆, 周志春, 等. 2004. 木荷苗木性状的种源变异和地理模式. 林业科学研究, 17(2): 192-198.
(Zhang P, Jin G Q, Zhou Z C,etal. 2004. Provenance difference and geographic variation pattern for seedling trait ofSchimasuperba. Forest Research, 17(2):192-198. [in Chinese])
张 萍, 周志春, 金国庆, 等. 2006. 木荷种源遗传多样性和种源区初步划分. 林业科学, 42(2): 38-42.
(Zhang P, Zhou Z C, Jin G Q,etal. 2006. Genetic diversity analysis and provenance zone allocation ofSchimasuperbain China using RAPD markers. Scientia Silvae Sinicae, 42(2): 38-42. [in Chinese])
张 蕊, 王 艺, 金国庆, 等. 2013. 施氮对木荷3个种源幼苗根系发育和氮磷效率的影响. 生态学报, 33(12): 3611-3621.
(Zhang R, Wang Y, Jin G Q,etal. 2013. Nitrogen addition affects root growth, phosphorus and nitrogen efficiency of three provenances ofSchimasuperbain barren soil. Acta Ecologica Sinica, 33(12): 3611-3621. [in Chinese])
赵 冰, 张启翔. 2007. 中国蜡梅种质资源核心种质的初步构建. 北京林业大学学报, 29(1): 16-21.
(Zhao B, Zhang Q Q. 2007. Preliminary construction of the core germplasm ofChimonanthuspraecoxin China. Journal of Beijing Forestry University, 19(1): 16-21. [in Chinese])
Brown A H D. 1989. Core collection: A practical approach to genetic resources management. Genome, 31(2): 818-824.
Belaj A, Atienza S G, Urdíroz N M,etal. 2012. Developing a core collection of olive (OleaeuropaeaL.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genetics & Genomes, 8(2): 365-378.
Cipriani G, Spadotto A, Jurman I,etal. 2010. The SSR-based molecular profile of 1005 grapevine (VitisviniferaL.) accessions uncovers new synonymy and parentages, and reveals a large admixture amongst varieties of different geographic origin. Theoretical & Applied Genetics, 121(8): 1569-1585.
Escribano P, Viruel M A, Hormaza J I. 2008. Comparison of different methods to construct a core germplasm collection in woody perennial species with simple sequence repeat markers. A case study in cherimoya (Annonacherimola, Annonaceae), an underutilised subtropical fruit tree species. Annals of Applied Biology, 153(1): 25-32.
Frankel O, Brown A. 1984. Plant genetic resources today: a critical appraisal// Holden J H W, Williams J T. Crop genetic resources: Conservation and evaluation. George Allan and Unwin, London, 249-257.
Heckenberger M, Bohn M, Ziegle J S,etal. 2002. Variation of DNA fingerprints among accessions within maize inbred lines and implications for identification of essentially derived varieties. Molecular Breeding, 10(4):181-191.
Liang W, Dondini L, Franceschi P D,etal. 2014. Genetic diversity, population structure and construction of a core collection of apple cultivars from Italian germplasm. Plant Molecular Biology Reporter, 33(3): 1-16.
Liu K, Muse S V. 2005. PowerMarker: an integrated analysis environment for genetic marker analysis. Bionformatics, 21(9): 2128-2129.
Miyamoto N, Ono M, Watanabe A. 2015. Construction of a core collection and evaluation of genetic resources forCryptomeriajaponica(Japanese cedar). Journal of Forest Research, 20(1): 1-11.
Niu H Y, Li X Y, Ye W H,etal. 2012. Isolation and characterization of 36 polymorphic microsatellite markers inSchimasuperba(Theaceae). Journal of Cell Biology, 18(1): 153-166.
Peakall R, Smouse P E. 2012. GenAlEx6.5: genetic analysis in Excel. Population genetic software for teaching and research—an update. Bioinformatics, 28(28): 2537-2539.
Schoen D J, Brown A H D. 1995. Maximizing genetic diversity in core collections of wild crop relatives of crop species//Sharma B. Core Collection of Genetic Resources. Chicester, UK: John Wiley and Sons, 253-259.
Wang J C, Hu J, Xu H M,etal. 2007. A strategy on constructing core collections by least distance stepwise sampling. Theoretical & Applied Genetics, 115(1): 1-8.
Wang Y, Fu J X, Pan L J,etal. 2014. Construction of core collection of lychee by SSR marker. Acta Horticulturae, 1029(1029): 87-92.
Zhang H, Zhang D, Wang M,etal. 2011. A core collection and mini core collection ofOryzasativaL. in China. Theoretical & Applied Genetics, 122(1): 49-61.
Zhang H, Wang H, Guo S,etal. 2012. Identification and validation of a core set of microsatellite markers for genetic diversity analysis in watermelon,CitrulluslanatusThunb. Matsum. & Nakai. Euphytica, 186(2): 329-342.
(责任编辑 徐 红)
Construction of Core Collection ofSchimasuperbaBased onSSR Molecular Markers
Yang Hanbo1Zhang Rui1Wang Bangshun2Xu Zhaoyou2Zhou Zhichun1
(1.ZhejiangProvincialKeyLaboratoryofTreeBreedingResearchInstituteofSubtropicalForestryChineseAcademyofForestryHangzhou311400; 2.LongquanForestryResearchInstitute,ZhejiangProvinceLongquan323700)
【Objective】 The most suitable sampling strategy and proportion for core collection ofSchimasuperbawere defined through a comparative analysis and evaluation, and a core collection ofS.superbawas constructed. Molecular identity of the core collection was further studied, providing theoretical basis and core materials for further research and utilization ofS.superbagermplasms. It also provides a basis for the core collection construction of other woody plants.【Method】 Taking 754 accessions ofS.superbaas materials that were come from seven provinces of China. The methods of M strategy (maximization strategy, M), random sampling (R), simulated annealing algorithm maximizing the genetic diversity (SAGD) and simulated annealing algorithm maximizing the number of alleles (SANA) were used to construct the core collections using 13 SSR primer pairs. The parameters of genetic diversity, such as number of alleles (Na), effective number of alleles (Ne) and Shannon’s information index (I), were used to determine the optimal method.【Result】 128 alleles (Na) were detected in 13 SSR primer pairs, the average ofNewas 3.47. High genetic diversity was revealed in the germplasm resources ofS.superba(I=1.39). Comparative analysis showed that the core collection constructed by SANA, SAGD and M strategies were all better than R strategy. The retained ofNain the core collection was low, although the core collection with SANA and SAGD are better representation of the original collection. The retained ofNain the core collection that was constructed by M strategy was apparently higher than other core collections that were constructed by other three strategies. According to the parameters of genetic diversity, and the number of sampling, the core collection constructed by M strategy could preserve the greatest level of genetic diversity of germplasm resources with the minimum sample size, indicating that M strategy was optimal in terms of establishing the most representative core collection, although SANA and SAGD methods all have good representation. The principal coordinate analysis (PCoA) showed that the core collection constructed by M strategy could more comprehensively represent at the genetic diversity level ofS.superba. The 115 accessions ofS.superbaincludes 15.3% samples of the germplasms, the retention ratio ofNa,NeandIwere 93.8%, 115.6% and 109.9%, and the specific molecular identity for 115 core collections were established with 99.99% probability of confidence. These results demonstrated that the 115 accessions could stand for the initial collection, at the same time a set of unique molecular identity was established for 115 accessions based on 13 SSR loci.【Conclusion】 M strategy is a suitable method for constructingS.superbacore collection. These results demonstrated that the 115 accessions as core collection could represent the original germplasms, at the same time this research method of the construction of core collection would provide an example for other tree species.
Schimasuperba; SSR marker; core collection; molecular identity
10.11707/j.1001-7488.20170605
2016-06-29;
2016-08-24。
“十二五”国家科技支撑计划课题(2012BAD01B04);浙江省竹木农业新品种选育重大科技专项竹木育种协作组项目(2012C12908-6);福建省林木种苗科技攻关五期项目木荷课题;江西省林业厅林业科技创新专项项目(201503)。
S722; S718.46
A
1001-7488(2017)06-0037-10
*张蕊为通讯作者。
