载畜率对内蒙古荒漠草原冷蒿种群资源分配格局的影响
2017-06-27陈万杰杜宇凡王亚婷赵天启赵萌莉
古 琛,陈万杰,杜宇凡,王亚婷,赵天启,赵萌莉
内蒙古农业大学生态环境学院, 呼和浩特 010019
载畜率对内蒙古荒漠草原冷蒿种群资源分配格局的影响
古 琛,陈万杰,杜宇凡,王亚婷,赵天启,赵萌莉*
内蒙古农业大学生态环境学院, 呼和浩特 010019
研究植物的资源分配格局以揭示其对环境变化的响应机制有重要的生态学意义。以内蒙古短花针茅(Stipabreviflora)草原冷蒿(Artemisiafrigida)种群为研究对象,设置4个载畜率水平的随机区组试验,由冷蒿地上、地下及各构件生物量的变化入手,探讨了载畜率对该种群资源分配格局的影响规律,为退化草地的恢复及合理的放牧管理提供参考。结果表明:(1)冷蒿种群的高度随载畜率的增大而显著降低(P<0.05),中度与重度放牧显著降低了该种群的盖度(P<0.05),轻度放牧使该种群密度显著增加(P<0.05);(2)地上、地下生物量及总生物量均随载畜率的增加而显著降低(P<0.05),3a的年际效应及载畜率与年际的互作效应对总生物量、地上和地下生物量的影响差异均显著(P<0.05);(3)生物量分配的总体格局是根茎叶花/果,且各构件的生物量均随着载畜率的增加而减少;(4)各构件的生物量分配比例对载畜率的响应不同,中度与重度放牧显著增大了生物量在根的分配(P<0.05),茎的生物量分配在轻度放牧显著增加,重度放牧显著降低(P<0.05),轻度与中度放牧显著促进了叶的生物量分配(P<0.05),花/果的生物量分配随载畜率的增加而显著降低(P<0.05);(5)随着载畜率的增大,冷蒿的有性繁殖能力减弱,而无性繁殖能力增强。
家畜放牧;短花针茅;生物群系;种群特征;生物量分配比例
自Harper[1]提出植物构件理论以来,以构件生物量为核心研究资源分配格局已成为生态学研究的重要内容。平衡生长假说、最优分配理论、功能平衡假说均表明植物权衡生物量在根、茎和叶等器官中的分配,以获取满足其形态结构和生理活动所需的水、肥、气、热等资源[2- 3]。植物各功能器官的资源在不同生境中的分配始终存在竞争,并通过权衡生长、发育和繁殖等过程以达到理想的分配格局[4- 5],这在一定程度上反映了植物对环境变化的适应,有重要的生态学和进化意义[2,6]。许多学者分别就增温、增雨、遮阴等对植物资源分配的影响进行了研究[7- 8]。那不同放牧强度又是如何影响植物的生物量分配格局的?放牧是影响植物资源分配格局的主要因子之一[9],其所导致的植物种群变化最终都将反应在植物的构件和种群的结构上。放牧同时改变了草地生态系统中水、肥、气、热等环境因子和植物本身的生物因素,对植物资源分配格局影响的机制较为复杂。因此,研究放牧对植物资源分配的影响有重要生态学意义。
冷蒿(Artemisiafrigida)是菊科的一种小半灌木,返青早、枯落晚、营养价值高,耐干旱、耐啃食、耐践踏、且再生能力强[10],是牧场冬春季的主要优良牧草,在过度放牧下仍能维持较高的生长繁殖能力,在畜牧业生产中占有重要地位[11]。冷蒿是内蒙古草原群落的伴生种,甚至优势种,伴随着退化演替的各个阶段,是最可靠的正定量放牧指示植物,同时也是草原退化的阻击者,会对群落的结构与功能产生重要的影响[12- 14]。冷蒿草原可向禾草草原恢复,也可继续退化并最终消失[11]。因此,在生境严酷、稳定性差、生态系统脆弱的荒漠草原[15],研究放牧对冷蒿种群特征及生物量分配格局的影响,有利于荒漠草原的合理利用和退化恢复。
为此,本试验以荒漠草原的冷蒿种群为研究对象,由地上、地下及各构件生物量的变化入手,探讨载畜率对该种群特征及资源分配格局的影响规律,旨在回答以下3个问题:1)冷蒿种群的高度、盖度和密度随载畜率的变化有何种规律?2)不同载畜率下,冷蒿地上、地下及总生物量如何变化,各构件生物量如何变化?3)各构件的资源分配比例随载畜率如何变化?4)冷蒿的繁殖策略是否受载畜率的影响?通过分析种群特征及资源分配格局与载畜率的关系,为退化草地的恢复及草地放牧管理与畜牧业生产提供参考。
1 材料与方法
1.1 试验地概况
试验区位于阴山北麓,内蒙古自治区乌兰察布市西北部的葛根塔拉草原,四子王旗王府一队,内蒙古农牧科学院草原研究所荒漠草原实验基地(41°47′17″ N,111°53′46″ E)。地势平坦开阔,海拔1450 m。
该旗地处中温带大陆性季风气候区,具有干旱、少雨、多风和蒸发量大的气候特征。年均气温3.4 ℃,月平均温度最高月为6—8月,年均气温分别为21.5、24.0、23.5 ℃,≥0 ℃积温为2500—3100 ℃,≥10 ℃的年积温为2200—2500 ℃,无霜期90—115 d。平均日照时数为3117.7 h。年均降水量299.4 mm,且年际变化较大,降水主要集中在5—9月,这段时间的降水量占全年总降水总量的80%以上。年蒸发量约2300 mm远超过降水量,是降水量的7—10倍,致使空气干燥,干旱严重,湿润度0.15—0.30。
草地植被类型为短花针茅+冷蒿+无芒隐子草。草层低矮,平均高度为8 cm,盖度为17%—20%,植被较稀疏。植物群落由20多种植物组成,主要为多年生草本短花针茅(Stipabreviflora)、无芒隐子草(Cleistogenessongorica)、银灰旋花(Convolvulusammannii)、阿尔泰狗娃花(Heteropappusaltaicus)、冰草(Agropyroncristatum)、羊草(Leymuschinensis)、细叶葱(Alliumtenuissimum)、蒙古葱(Alliummongolicum)等;一、二年生草本有猪毛菜(Salsolacollina)、猪毛蒿(Artemisiascoparia)、栉叶蒿(Artemisiapectinata)等。还有灌木小叶锦鸡儿(Caraganamicrophylla)、矮灌木狭叶锦鸡儿(C.stenophylla)、半灌木驼绒藜(Ceratoideslatens)、小半灌木木地肤(Kochiaprostrata)、冷蒿(Artemisiafrigida)等。
土壤为淡栗钙土,土层厚度大约为1 m,但在40—50 cm的地方出现了钙积层,土体坚硬且渗透能力差,有机质含量较低。土壤微生物有好气性细菌、放线菌和霉菌等。
1.2 试验设计和测定方法
1.2.1 样地概况
图1 试验小区和区组示意图Fig.1 Schematic diagram for experimental plot and block CK: 对照control check;LG: 轻度放牧Light grazing;MG: 中度放牧Moderate grazing;HG: 重度放牧Heavy grazing
放牧试验于2003年6月开始,在内蒙古乌兰察布市四子王旗短花针茅草原上,选取地势相对平坦、植被均一的约50 hm2天然草地围封,进行不同载畜率放牧试验。采用完全随机区组设计,将围封的草地分为12个小区,3个区组(3次重复),每个区组中设4个处理(4个载畜率水平)(图1),分别为对照(CK)、轻度放牧(LG)、中度放牧(MG)和重度放牧(HG)。载畜率值依次为0、-0.91、1.82和2.71羊单位 hm-20.5a-1,绵羊只数分别为0、4、8和12只。每个区组内的4个处理完全随机排列,每个处理的面积基本相等为4.4 hm2。放牧使用当地成年蒙古羯羊。从2003年到2015年每年放牧期为半年,6月1日开始,11月30日结束。每日06:00将绵羊赶入各小区让其自由采食,到6:00赶回圈内饮水、休息。
1.2.2 测定方法
每年放牧开始前,在各小区分别围封10个1 m×1 m样方以测定群落特征。试验野外采样于2013—2015年每年8月中旬生物量的最高峰时期进行。在已围封好的样方内分种测定各种群的高度、盖度、密度和生物量。再在每个小区内对冷蒿的单优斑块进行10次重复的随机取样,以保证地上地下完整连接为前提,挖取30 cm×30 cm×40 cm的土柱,地上部分齐地面刈割后,按构件分为茎、叶、花/果3类;地下部分放入根袋带回实验室冲洗干净,挑去其他植物根系及杂物后为地下生物量。将地上部分各构件和地下部分于65 ℃烘干24 h后称其干重。
1.2.3 数据分析方法
试验数据用EXCEL 2013软件进行数据整理、简单分析和做图表,不同处理间冷蒿种群特征及资源分配格局的差异性用SAS 9.4在α = 0.05显著性水平下进行单因素方差分析(one-way ANOVA),处理与年际交互作用的差异性采用双因素方差分析(two-factor ANOVA)进行。
2 结果与分析
2.1 载畜率对冷蒿种群特征的影响
载畜率对冷蒿种群盖度和高度的影响变化趋势一致。中度放牧和重度放牧显著降低了种群的盖度(P<0.05),与对照相比分别降低了57%和83%(图2),轻度放牧与对照相比差异不显著(P0.05)。随载畜率的增加,冷蒿的高度与对照相比分别降低了39%、62%和74%,不同处理间差异均显著(P<0.05)(图2)。轻度放牧使冷蒿的密度显著增大(P<0.05),分别是对照、中度放牧和重度放牧的1.6、2.1和2.4倍(图2)。
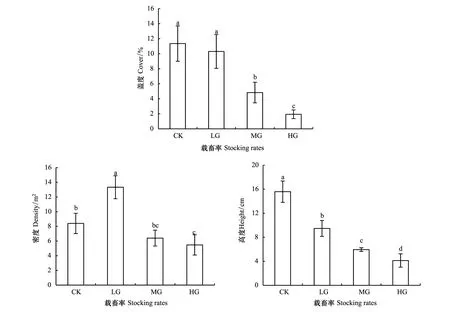
图2 不同载畜率冷蒿的种群特征Fig.2 Population characteristics of Artemisia frigida under different stocking rates同一柱形图中不同字母表示不同处理间差异显著(P<0.05)
2.2 载畜率对冷蒿种群生物量的影响
冷蒿地上、地下生物量随载畜率增大而降低。对照最大,轻度放牧次之,中度放牧与重度放牧最小,除后两者间差异不显著(P0.05)外,其他处理间差异显著(P<0.05)。3a平均地上、地下生物量,对照分别约是轻度放牧、中度放牧、重度放牧的3、7和14.5倍(图3)。总生物量也随着载畜率的增大而显著降低,对照的生物量显著高于其他处理(P<0.05);2013年对照依次是轻度放牧、中度放牧、重度放牧的3.9、10.2和16.8倍,2014和2015年对照分别约是轻度放牧、中度放牧、重度放牧的2、4.6和14倍(图3)。载畜率、年际效应及载畜率与年际的互作效应对总生物量、地上、地下生物量的影响差异显著(表1)。
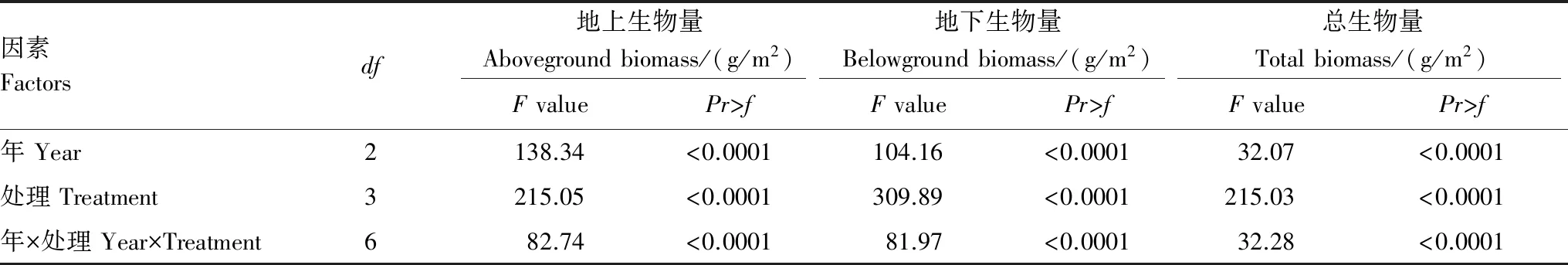
表1 冷蒿生物量双因素方差分析
载畜率对各构件生物量也有显著影响。茎、叶与花/果的生物量均随载畜率的增加而显著降低(P<0.05),对照轻度放牧中度放牧重度放牧。对照茎的生物量依次是轻度放牧、中度放牧、重度放牧的3.1、8.7和17.7倍,叶的生物量分别是2.7、6.8、14.4倍,花/果的生物量是11.0、63.4、190.3倍。在中度放牧和重度放牧,花/果几乎消失(图3)。
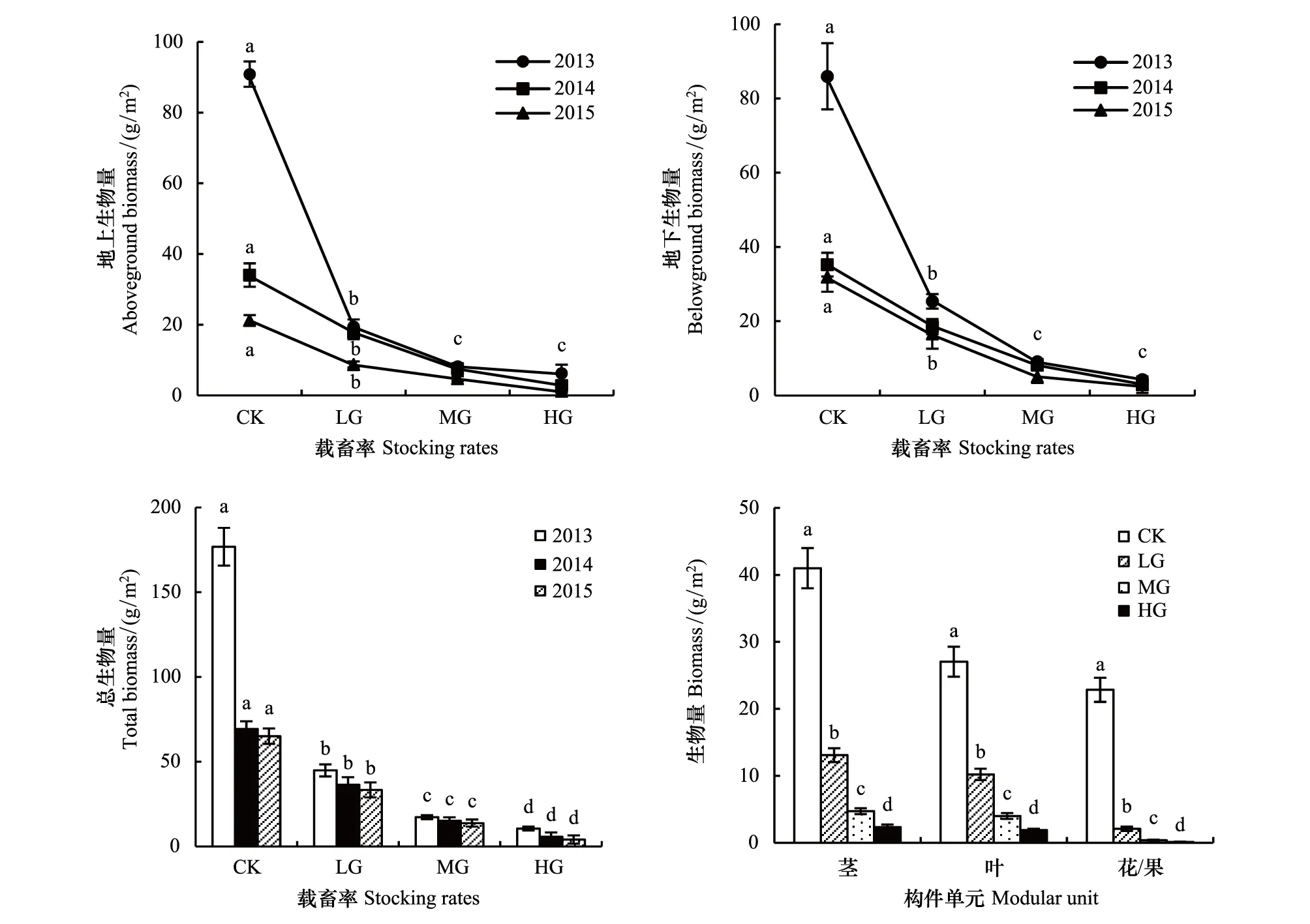
图3 不同载畜率冷蒿生物量Fig.3 Biomass of Artemisia frigida under different stocking rates
2.3 载畜率对冷蒿资源分配的影响
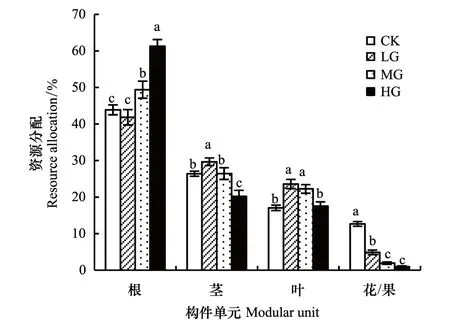
图4 不同载畜率冷蒿资源分配 Fig.4 Resource allocation of Artemisia frigida under different stocking rates
冷蒿生物量总体分配格局是根茎叶花/果。载畜率对冷蒿各构件资源分配的影响不同(图4)。根的分配比例随载畜率的增大而增大,重度放牧为61.3%,最大;中度放牧49.4%,次之;对照与轻度放牧分别为43.9%和41.9%,最小;除后两者间差异不显著(P0.05)外,其他处理间差异显著(P<0.05)。随载畜率的增大,茎的分配比例先增大后减小。轻度放牧最大为29.7%,重度放牧最小为20.2%,与其它处理相比差异显著(P<0.05);对照与中度放牧均为26.4%居中,二者之间差异不显著(P0.05)。
叶的分配比例随载畜率的变化趋势与茎的相似,轻度放牧与中度放牧分别为23.65%和22.35%,显著高于对照(17.1%)和重度放牧(17.6%)(P<0.05)。
花/果的分配比例随载畜率的增加而显著递减(P<0.05),对照最大为12.7%,轻度放牧为4.9%,次之,然后是中度放牧为1.9%,重度放牧最小为1.0%。
3 讨论与结论
放牧是天然草原重要的影响因子和进化驱动力。群落特征与放牧强度紧密相关,当气候条件一致时,家畜放牧对草地生态系统的影响将超越其它环境因素,成为控制草地群落特征的主导因子[12]。
本研究结果表明,冷蒿种群盖度和高度均随载畜率的增大而显著降低,轻度放牧显著增加了冷蒿密度(图2)。冷蒿茎的斜倚程度随放牧强度的增大而逐渐增大,甚至匍匐于地面,高度急剧下降。由于放牧强度的增加,群落中高大禾草短花针茅的比例减少且群落高度降低,减弱了冷蒿因光竞争而伸长生长的趋势。冷蒿通过矮化躲避家畜的采食,是其对过度放牧的适应策略[16]。
放牧对植物最直接的影响就是采食植物的茎叶,降低叶面积指数,干扰植物碳水化合物的合成,影响植物正常的生长发育。放牧使植物光合器官出现小型化,导致个体生物量降低[17]。本研究中,冷蒿地上、地下生物量及总生物量均随载畜率的增加而显著降低(图3)。因为随着载畜率的增大,家畜的采食和践踏对冷蒿的破坏加剧,使其光合面积减少,再生能力下降,光合产物的积累降低。同时放牧家畜的过度践踏,使土壤的紧实度和容重增大,渗透性、含水量、孔隙度降低[18- 19],间接阻碍了根系的伸长与扩展[20]。随着载畜率的增大,冷蒿的地上、地下生物量、总生物量及在群落中的竞争力下降,最终导致整个种群的退化[21]。
植物资源分配是生活史理论研究的核心内容[22- 23]。植物地上、地下生物量的积累与分配主要受到环境中可利用资源的影响[24],根、茎、叶、花/果等构件生物量的分配决定了植株同化能力的大小,个体的生长发育也依赖于各构件单元资源分配的权衡。植物通过生物量分配格局的改变来响应生境的异质性,以提高自身的同化效率[22],更好的获取生长、发育和繁殖所需的水分、温度、光照、养分等资源[2,25]。本研究表明,冷蒿生物量分配的总体格局是根茎叶花/果,且各构件的生物量均随着载畜率的增加而降低(图3)。
杜丽霞、王静等研究认为放牧增加了冷蒿根系的生物量分配[26- 27],本研究也得出相似的结论。中度与重度放牧显著增大了根的生物量分配。放牧家畜对土壤的压实显著促进了根系生物量分配比例的增加[2]。随着放牧强度的增加,冷蒿通过增加根系的生物量分配以确保吸收足够的水分和养分来供给地上部分的生长发育,增加种群适合度,以便更好的适应环境,促进种群的扩散[11]。
Quezada和Gianoli指出植物增加茎的生物量分配以补偿放牧的采食[9],武瑞鑫等也认为披针叶黄华(Thermopsislanceolata)在放牧条件下将更多的资源分配给植株的地上部分,因为放牧家畜不采食该物种而采食其他优良牧草[28]。本试验中,由于家畜对冷蒿的采食和践踏,轻度放牧增加了茎的生物量分配,重度放牧使之显著降低,轻度与中度放牧显著促进了叶的生物量分配。在适度放牧条件下,冷蒿种群叶面积减小,为了缓解因采食而引起的光合作用下降,往往将光合产物优先分配给地上部分,尤其是光合器官叶片和茎[29],以提高对光的截获能力和再生能力,尽快恢复其光合作用[30],是冷蒿对频繁牧食的主要适应机制之一。但过度放牧使冷蒿在茎和叶的生物量分配显著降低。
繁殖策略是植物对环境的生殖适应趋势,是资源向生存、生长和繁殖活动中最适分配的结果。植物在不同生境中的繁殖对策反映了对环境的适应和生殖的潜能。本试验得出花/果的生物量分配随载畜率的增加而显著降低。随着载畜率的增大,冷蒿有性繁殖分配减少,生殖构件显著下降,生殖枝数和种子产量降低,甚至在重度放牧中有性生殖几乎消失。冷蒿以无性生殖为主,不定根和营养枝是其无性生殖器官,且随着载畜率的增大,无性系分株比种子形成的幼苗更易存活,因此冷蒿生殖格局由有性与无性生殖并存向单一的营养繁殖转变[16,26- 27,31]。
放牧的践踏使冷蒿在倒伏的枝条上形成不定根,促进其更新芽的生根及萌蘖,营养枝分化率增大。轻度放牧下,冷蒿通过枝条、不定根、匍匐茎的增加,增强对资源的有效利用,加强在群落中的竞争力,扩大种群的空间分布,优势地位逐渐提高甚至取代原生建群种[11,16,21,31]。
[1] Harper J L. Population Biology of Plants. London: Academic Press, 1977.
[2] Poorter H, Niklas K J, Reich P B, Oleksyn J, Poot P, Mommer L. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytologist, 2012, 193(1): 30- 50.
[3] Freschet G T, Kichenin E, Wardle D A. Explaining within-community variation in plant biomass allocation: a balance between organ biomass and morphology above vs below ground?. Journal of Vegetation Science, 2015, 26(3): 431- 440.
[4] 安珏, 安慧. 宁夏荒漠草原优势植物生长及生物量分配对放牧干扰的响应. 西北植物学报, 2015, 35(2): 373- 378.
[5] 王一峰, 靳洁, 候宏红, 赵博, 曹家豪, 李筱姣. 川西风毛菊花期资源分配随海拔的变化. 植物生态学报, 2015, 39(9): 901- 908.
[6] 姚洁, 曾波, 杜珲, 潘晓娇, 苏晓磊. 三峡水库长期水淹条件下耐淹植物甜根子草的资源分配特征. 生态学报, 2015, 35(22): 7347- 7354.
[7] 马志良, 高顺, 杨万勤, 朱鹏, 吴福忠, 谭波. 遮荫对撂荒地草本群落生物量分配和养分积累的影响. 生态学报, 2015, 35(16): 5279- 5286.
[8] 吕晓敏, 王玉辉, 周广胜, 许振柱, 陈军, 谭丽萍, 刘涛. 温度与降水协同作用对短花针茅生物量及其分配的影响. 生态学报, 2015, 35(3): 752- 760.
[9] Quezada I M, Gianoli E. Counteractive biomass allocation responses to drought and damage in the perennial herbConvolvulusdemissus. Austral Ecology, 2010, 35(5): 544- 548.
[10] 中国科学院内蒙古宁夏综合考察队. 内蒙古植被. 北京: 科学出版社, 1985: 631- 644.
[11] 杨持, 宝音陶格涛, 李良. 冷蒿种群在不同放牧强度胁迫下构件的变化规律. 生态学报, 2001, 21(3): 405- 408.
[12] 李永宏. 内蒙古锡林河流域羊草草原和大针茅草原在放牧影响下的分异和趋同. 植物生态学与地植物学学报, 1988, 12(3): 189- 196.
[13] 李永宏. 内蒙古草原草场放牧退化模式研究及退化监测专家系统雏议. 植物生态学报, 1994, 18(1): 68- 79.
[14] 王炜, 刘钟龄, 郝敦元, 梁存柱. 内蒙古草原退化群落恢复演替的研究I.退化草原的基本特征与恢复演替动力. 植物生态学报, 1996, 20(5): 449- 459.
[15] 古琛, 杜宇凡, 乌力吉, 陈万杰, 薛文杰, 赵萌莉. 载畜率对荒漠草原群落及植物功能群生物量的影响. 生态环境学报, 2015, 24(12): 1962- 1968.
[16] 王静, 杨持, 王铁娟. 冷蒿(Atemisiafrigida)种群在放牧干扰下构件的变化. 生态学报, 2006, 26(3): 960- 965.
[17] 李西良, 刘志英, 侯向阳, 吴新宏, 王珍, 胡静, 武自念. 放牧对草原植物功能性状及其权衡关系的调控. 植物学报, 2015, 50(2): 159- 170.
[18] 戎郁萍, 韩建国, 王培, 毛培胜. 放牧强度对草地土壤理化性质的影响. 中国草地, 2001, 23(4): 41- 47.
[19] 安慧, 徐坤. 放牧干扰对荒漠草原土壤性状的影响. 草业学报, 2013, 22(4): 35- 42.
[20] 董亭, 李群, 赵萌莉, 贾乐, 阿穆拉, 韩国栋, 白玮杰. 放牧对大针茅根系生物量影响的研究. 草地学报, 2011, 19(2): 237- 341.
[21] 李金花, 李镇清, 刘振国. 不同刈牧强度对冷蒿生长与资源分配的影响. 应用生态学报, 2004, 15(3): 408- 412.
[22] Müller I, Schmid B, Weiner J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology, Evolution and Systematics, 2000, 3(2): 115- 127.
[23] 党晶晶, 赵成章, 王继伟. 高寒草地甘肃臭草斑块不同发育阶段茎-叶生物量分配: 异速生长分析. 生态学杂志, 2015, 34(6): 1540- 1545.
[24] Enquist B J, Niklas K J. Global allocation rules for patterns of biomass partitioning in seed plants. Science, 2002, 295(5559): 1517- 1520.
[25] Smith S E. Variation in response to defoliation between populations ofBoutelouacurtipendulavar.caespitosa(Poaceae) with different livestock grazing histories. American Journal of Botany, 1998, 85(9): 1266- 1272.
[26] 杜利霞, 李青丰, 董宽虎. 放牧强度对短花针茅草原冷蒿繁殖特性的影响. 草地学报, 2007, 15(4): 367- 370.
[27] 王静, 杨持, 王铁娟. 放牧退化群落中冷蒿种群生物量资源分配的变化. 应用生态学报, 2005, 16(12): 2316- 2320.
[28] 武瑞鑫, 钟梦莹, 潘多, 邵新庆, 李超群, 刘月华, 位晓婷, 张德里. 青藏高原东缘不同恢复措施对披针叶黄华繁殖构件及资源分配的影响. 草地学报, 2015, 23(1): 34- 40.
[29] Wilsey B J. Urea additions and defoliation affect plant responses to elevated CO2in a C3grass from Yellowstone National Park. Oecologia, 1996, 108(2): 321- 327.
[30] King J, Sim E M, Barthram G T, Grant S A, Torvell L. Photosynthetic potential of ryegrass pastures when released from continuous stocking management. Grass and Forage Science, 1988, 43(1): 41- 48.
[31] 焦树英, 韩国栋, 刘俊清. 荒漠草原地区冷蒿构件及其动态规律对载畜率的响应. 中国草地学报, 2009, 31(4): 80- 84.
Stocking rates affect the resource allocation patterns ofArtemisiafrigidain the inner mongolian desert steppe
GU Chen, CHEN Wanjie, DU Yufan, WANG Yating, ZHAO Tianqi, ZHAO Mengli*
CollegeofEcologyandEnvironmentalScience,InnerMongoliaAgriculturalUniversity,Hohhot010019,China
Studying the patterns of plant resource allocation is of crucial ecological significance, as well as investigating how these patterns respond to environmental change. This study intends to provide a reference for grassland degradation and restoration, as well as grassland grazing management. Therefore, we investigated the biomass distribution ofArtemisiafrigidain theStipabrevifloradesert steppe in Inner Mongolia under different stocking rates. A randomized block experiment that included four different stocking rates was set up in the field. From these treatments, the total biomass, above- and belowground biomass, and biomass from different plant parts were collected and analyzed. Our results indicated that: (1) the height ofA.frigidasignificantly decreased with the increase in stocking rates (P< 0.05), andA.frigidacover significantly decreased by moderate and heavy grazing (P< 0.05), although its density increased remarkably under light grazing (P< 0.05). (2) The above- and belowground biomass and total biomass ofA.frigidasignificantly decreased with increasing stocking rates (P< 0.05), and the influence of 3-year interannual effects, stocking rates, and interannual interactions on total biomass and above- and belowground biomass was significant (P< 0.05). (3) The biomass was differentially distributed inA.frigidasegments, with the highest values measured in the roots, and lower values in the stem, leaf, and flowers or fruits, respectively. Furthermore, the biomass per plant part significantly decreased with increasing stocking rates (P< 0.05). (4) The biomass distribution ratio perA.frigidaplant part was different under various stocking rates, with root biomass distribution increasing with moderate and heavy grazing (P< 0.05), and biomass accumulation in the stem significantly increasing with light grazing and significantly decreasing heavy grazing (P< 0.05). In addition, light and moderate grazing enhanced leaf biomass accumulation (P< 0.05), whereas and fruit biomass significantly decreased with increasing stocking rates (P< 0.05). (5) Finally, increasing stocking rates reduced the sexual reproduction ability ofA.frigida, and promoted its asexual propagation ability.
livestock grazing;Stipabreviflora; steppe biome; population characteristics; biomass allocation ratios
10.5846/stxb201601140089
国家自然科学基金(31460110,31170446);内蒙古农业大学草地资源教育部重点实验室
2016- 01- 14; 网络出版日期:2016- 08- 30
古琛,陈万杰,杜宇凡,王亚婷,赵天启,赵萌莉.载畜率对内蒙古荒漠草原冷蒿种群资源分配格局的影响.生态学报,2017,37(7):2237- 2243.
Gu C, Chen W J, Du Y F, Wang Y T, Zhao T Q, Zhao M L.Stocking rates affect the resource allocation patterns ofArtemisiafrigidain the inner mongolian desert steppe.Acta Ecologica Sinica,2017,37(7):2237- 2243.
*通讯作者Corresponding author.E-mail: nmgmlzh@126.com
