复合乳酸菌协同酵母菌发酵对苏打饼干品质特性的影响
2017-06-19闫博文赵建新张均叶范大明郝玉洁管璐静陈卫张灏
闫博文,赵建新*,张均叶,2,范大明,郝玉洁,管璐静,陈卫,3,张灏
1(江南大学 食品学院,江苏 无锡,214122)2(亿滋食品(中国)有限公司,江苏 苏州, 215126) 3(江南大学, 国家重点实验室, 江苏 无锡, 214122)
复合乳酸菌协同酵母菌发酵对苏打饼干品质特性的影响
闫博文1,赵建新1*,张均叶1,2,范大明1,郝玉洁1,管璐静1,陈卫1,3,张灏1
1(江南大学 食品学院,江苏 无锡,214122)2(亿滋食品(中国)有限公司,江苏 苏州, 215126) 3(江南大学, 国家重点实验室, 江苏 无锡, 214122)
开发了一款复合乳酸菌发酵剂,通过协同酵母菌发酵制备苏打饼干,考察其对产品感官风味和营养功能特性的影响。研究结果表明,与单一乳酸菌菌株相比较,复合乳酸菌发酵剂具有良好的产酸速率,可有效降低饼干中丙烯酰胺的含量,且具有较好的消费者感官喜好度,适用于工业化生产。通过对不同发酵剂制得苏打饼干的消化率、维生素含量及γ-氨基丁酸含量(GABA)的分析发现,经复合乳酸菌协同酵母发酵可显著提高苏打饼干的蛋白质消化率、VB1与VB2含量及GABA含量,有助于苏打饼干功能营养特性的提高。
乳酸菌;酵母菌;发酵;苏打饼干;品质;营养
饼干,主要指的是一种以小麦粉为主要原料,经原辅料调粉、成型、烘烤等工艺制备的食品[1]。2014年的饼干产业概况显示[2],全球饼干市场份额的85.5%主要以曲奇类饼干为主,而造成发酵饼干市场占有量较低的原因主要是由于其虽口感松脆,但缺乏如酥性饼干般厚重的香味,产品风味单一。然而随着生活水平的提高,人们对食品的关注点逐渐从温饱转移到营养健康领域,具有高糖高油含量的曲奇类酥性饼干让众多消费者望而却步,这使得具有低油脂低糖分的发酵饼干重新获得了市场的青睐。因此,如何既能保持发酵饼干的营养健康特性,同时还可有效改善其感官风味水平是现今亟待解决的问题。
近年来研究发现,乳酸菌协同酵母发酵制备烘焙产品可有效改善产品风味、质地和营养特性等,可作为一种天然的烘焙改良剂,以改善产品因单一酵母发酵而导致产品风味不足,品质劣变较快等弊端。此外,张均叶[3]等人研究还发现采,用乳酸菌协同酵母发酵可有效降低饼干中所含丙烯酰胺的含量,降低经高温烘焙产品的食品安全隐患,使产品更加营养健康。因此,为了迎合当今市场需求,本研究旨在开发一款适用于发酵饼干的复合乳酸菌发酵剂,使其可有效改善产品风味及营养等品质特性。
1 材料和方法
1.1 材料与试剂
1.1.1 菌株
乳酸乳球菌乳酸亚种XX3,植物乳杆菌SQ1,江南大学食品生物技术中心保藏;即发型活性干酵母,安琪酵母股份有限公司。
1.1.2 主要材料与试剂
低筋小麦粉,益海嘉里食品工业有限公司;起酥油,天津南侨食品有限公司;白砂糖,中粮屯河股份有限公司;食用盐,江苏省盐业集团有限公司;脱脂乳粉,光明乳业股份有限公司。
1.2 仪器与设备
79480-30冻干系统,美国LABCONCO公司;ATLAS MOTOR 150 压面机,意大利MARCATO公司; FE20 pH计,美国METTLER TOLEDO公司;Trace MS 气相色谱-质谱联用仪,美国Finnigan公司;HPLC-1525 高效液相色谱,美国Waters公司; SCINO KT260半自动凯氏定氮仪,丹麦FOSS分析仪器公司。
1.3 试验方法
1.3.1 乳酸菌培养基的配制
(1)乳酸菌液体培养基(MRS)的配制参照周家春[4]的配方并稍作修改,具体为:蛋白胨10 g,牛肉浸膏10 g,酵母提取物5 g,葡萄糖20 g,乙酸钠5 g,柠檬酸氢二铵2 g,K2HPO42 g,MgSO4·7H2O 0.58 g,MnSO4·4H2O 0.19 g,吐温-80 1 mL,调节至pH 6.2~6.4。
(2)MRS琼脂培养基是在MRS培养基配方的基础上,添加1.5%~2.0%的琼脂。
1.3.2 乳酸菌菌粉的制备
(1)取经MRS液体培养基活化的乳酸菌菌液于8 000 r/min、4 ℃冷冻离心10 min,弃上清培养基,并用无菌生理盐水冲洗菌体2~3次并弃上清液;
(2)用无菌脱脂乳液重悬菌体,调节至菌浓度在8×109~1×1010CFU/mL,将乳酸菌脱脂乳液分装于无菌干燥冻干瓶中,冻干。
1.3.3 饼干的制备
酵母发酵饼干按PEDERSEN等[5]的配方修改后制备的,配方如表1所示。

表1 发酵饼干制作配方
(1)将饼干成型面团静置于温度35 ℃、湿度70%的醒发箱内发酵10 h;
(2)发酵完成的面团用压片机压至2 mm厚,并切割成5 cm×5 cm的面片,在上火180 ℃、下火160 ℃下烘烤15 min,冷却后得到酵母发酵饼干的成品。
1.3.4 样品pH及总滴定酸度的测定
面团及饼干的pH和总滴定酸度(Total titratable acid, TTA)测定参照的是国标GB/T 22427.9—2008[6]制订的方法,并稍作修改。其具体的测定方法如下:
(1)称取10.0 g待测样品,置于90 g煮沸去离子水中,溶液于磁力搅拌器上搅拌30 min,静置至溶液恢复室温;
(2)以0.1 mol/L NaOH溶液滴定溶液至pH 8.3,滴定消耗的NaOH溶液体积即为样品的TTA,单位为mL。
1.3.5 丙烯酰胺含量的测定
样品的预处理方法按GB/T 5009.204—2005[7]中的方法并稍作修改:
(1)准确称取粉碎的样品10.0 g,置于90 g无菌去离子水中,溶液于磁力搅拌器上搅拌30 min;
(2)搅拌混匀的混合物以5 000 r/min离心5 min,吸取上清液30 mL于分液漏斗中;
(3)用20 mL正己烷萃取上清液,收集下层水相;
(4)收集的水相以10 000 r/min冷冻离心(4 ℃)30 min,收集上清液并用双层滤纸过滤;
(5)向20 mL滤液中加入7.5 g KBr、0.4 mL氢溴酸和8 mL饱和溴水,避光冷藏放置15 h进行溴化衍生;
(6)向衍生液中逐滴加入硫代硫酸钠溶液至衍生液褪色,向溶液中加入25 mL乙酸乙酯,振荡混匀30 min,溶液静置分层;
(7)收集上层乙酸乙酯层,过0.22 μm有机滤膜,待测;
(8)丙烯酰胺的定性及定量参照HAASE[8]的方法并稍作修改,GC-MS的具体设置为:
色谱条件
色谱柱:HPINNOwax毛细管柱,60 m×0.25 mm×0.25 μm;升温程序:色谱柱温度60 ℃,保留2 min→升温15 ℃/min直到240 ℃→终温保留16 min。
质谱条件
进样口温度:200 ℃,离子源温度:180 ℃;离子源:EI源,70 eV;测定方式:离子监测方式(SIM),选择监测离子(m/z):152、150、108、106;进样方式:恒流1.0 mL/min,无分流进样,载气为氦气(99.999%)。
以不同浓度的丙烯酰胺标准品(0.0~1000.0 μg/L)作为外标,通过样品所测的特征峰峰面积在图1所示的丙烯酰胺标准品标准曲线中的代入值确定样品中丙烯酰胺的含量,标准曲线为:y=122.06x-4 767,R2=0.989 9。

图1 丙烯酰胺标准曲线Fig.1 Standard curve of acrylamide concentration
1.3.6 蛋白质体外消化率的测定
发酵饼干蛋白质体外消化率的测定采用的是两步消化法,具体测定步骤参照张庆[9]的方法。
1.3.7 VB1(硫胺素)与VB2(核黄素)含量的测定
制备发酵饼干中硫胺素与核黄素的水解提取液,具体测定步骤参照LEPORATI等[10]的方法。
以不同浓度的脱氢硫胺素(0~420.0 μg/mL)及核黄素(0~120.0 μg/mL)标准品作为外标,通过样品所测的特征峰峰面积在图2所示的维生素标准曲线中的代入值确定样品中硫胺素与核黄素的含量,标准曲线为:y=0.024x-0.099 7,R2=0.992 6(VB1标准曲线)、y=0. 156 8x-0.512 3,R2=0.994 9(VB2标准曲线)。

图2 脱氢硫胺素与核黄素标准曲线Fig.2 Standard curve of thiochrome and riboflavin concentration
1.3.8 VE含量的测定
发酵饼干中VE含量的测定参照国标GB/T 5009.82—2003[12]的方法并稍作修改,其具体测定方法如下:
(1)样品粉碎并过1 mm筛,称取10.00 g过筛样品于皂化瓶内,向皂化瓶内加入30 mL无水乙醇,将皂化瓶置于磁力搅拌器上搅拌20 min;
(2)向溶液内加入10%抗坏血酸5 mL、内标2.00 mL(维生素E乙酸酯标准品)、50% KOH溶液10 mL并混匀,沸水回流30 min后置于冰水浴中冷却;
(3)皂化完成的溶液入分液漏斗,以50 mL水洗皂化瓶2~3次,再以100 mL乙醚洗皂化瓶2~3次,将水洗液和醚洗液一并入分液漏斗,振荡混匀,静置分层,弃下层水层;
(4)以50 mL水洗分液漏斗乙醚层至水层不显碱性,收集分液漏斗内上层乙醚层提取液;
(5)提取液经5 g无水硫酸钠脱水后入旋蒸瓶,用100 mL乙醚冲洗分液漏斗2~3次,将冲洗液并入旋蒸瓶,55 ℃水浴旋蒸至瓶内约余2 mL乙醚溶液,利用氮吹仪吹干乙醚,立即加入2.00 mL无水乙醇,充分混合并溶解提取物;
(6)乙醇液以5 000 r/min离心5 min,上清液过0.22 μm有机滤膜,待测;
(7)样品中VE的含量测定的具体设置为:
色谱条件
色谱柱:ultrasphere ODS,25 cm × 4.6 mm × 5 μm;流动相:V(甲醇)∶V(水)= 98∶2;流速:1.7 mL/min;UV波长:300 nm;进样量:20 μL。
通过样品所测的特征峰峰面积与内标特征峰峰面积的比值确定样品中VE的含量。
1.3.9 γ-氨基丁酸(GABA)含量的测定
准确称取10.0 g粉碎样品并加入5%三氯乙酸25 mL,高速匀浆30 s;用双层滤纸进行过滤,滤液以10 000 r/min离心10 min;上清液用邻苯二醛进行衍生,衍生完成的溶液上HPLC,使用外标法测定其中GABA的含量。HPLC的具体设置参照LV等[13]的方法。
1.3.10 发酵饼干的感官评价
1.3.10.1 乳酸菌发酵饼干风味特征的确定
乳酸菌发酵饼干的感官评价描述词确定的方法参照VARAPHA等[14]的方法,并稍作修改。
(1)不同发酵剂制备的饼干被切割成5 cm×5 cm的方块,饼干样品均保存于干燥密封无异味的塑料罐内,为了降低贮藏对感官品质的影响,所有样品均在制备完成后72 h以内完成感官评价;
(2)乳酸菌发酵饼干的感官评价小组由15名以前从事过饼干感官评定的专业食品感官人员组成(其中男性6名、女性9名),所有感官评价小;组成员前期需接受1.5 h/天为期4天的关于乳酸菌发酵产品的感官术语的培训;
(3)在评定初期,使用12种不同乳酸菌制备的样品用于感官术语的确定,评定人员被要求尽可能多且针对性地提出单一风味特征的描述性词语(评定人员不需考虑此风味对产品是否存在积极或消极影响);
(4)评价小组成员对每个样品的描述词汇开展讨论直至成员对该词汇在对样品的风味贡献的描述程度上达成一致;
(5)选取12种样品共有的描述词汇,确定为乳酸菌发酵饼干的风味描述词汇表。经过讨论,感官评价小组确定的乳酸菌发酵饼干的风味特征描述词如表所示。
准备具有每一描述词汇风味的典型参考物质,评价小组成员通过讨论确定参考物质的评分,满分10分,分数越高代表参考物质该风味越强烈。
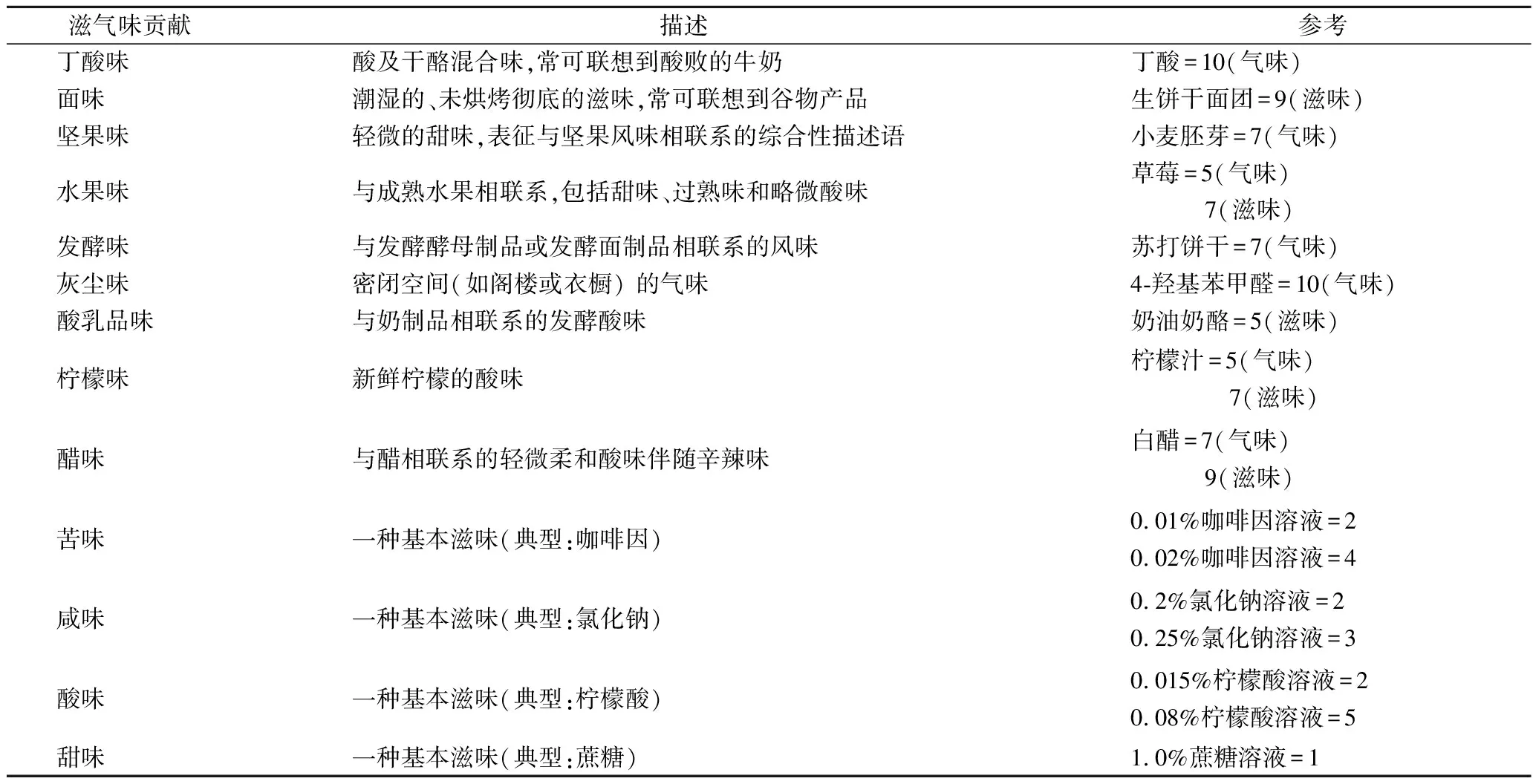
表2 乳酸菌发酵饼干感官特征描述词汇表
1.3.10.2 喜好度评分
(1)20名日常喜好饼干的消费者(其中男性7名、女性13名)组成消费者评价小组,对不同组样品饼干进行喜好度评分;
(2)满分10分,1~10分分别代表:1~2分,非常不喜欢;3~4分,比较不喜欢;5~6分,既不喜欢也不讨厌;7~8分,比较喜欢;9~10分,非常喜欢。
1.3.11 数据处理及分析
本研究中的实验除在方法中另提及外,均执行3次平行实验。利用SPSS Statistics 17.0软件对实验数据进行单因素ANNOVA分析及聚类分析,Tukey HSD被用于确定P≤0.05水平时数据的显著性。
2 结果与分析
本课题组的前期研究发现,乳酸菌协同酵母菌发酵制得饼干产品,当其产品pH值达到pH 4.2~4.4时,产品具有较好的消费者感官喜好度评价。而乳酸乳球菌乳酸亚种XX3虽能显著提高发酵饼干的风味且具有较好的消费者感官评价,但其在发酵过程中酸化速率过慢,不适于工业化生产需求,且其对降低饼干中丙烯酰胺的能力较差。而植物乳杆菌SQ1虽能有效地降低发酵饼干中的丙烯酰胺含量,且酸化速率较快,通常8~10 h即可达到发酵终点,但其对于饼干的风味无显著的改善作用。因此研究采用XX3及SQ1制备复合发酵剂,考察其对发酵饼干的酸化速率、风味及产品营养特性的影响。
2.1 复合乳酸菌对发酵饼干面团产酸速率的影响
为了验证复合乳酸菌协同酵母发酵对面团产酸速率的影响,本研究通过添加不同发酵剂观察其对面团发酵过程中pH和滴定酸度的变化,其结果如图3所示。
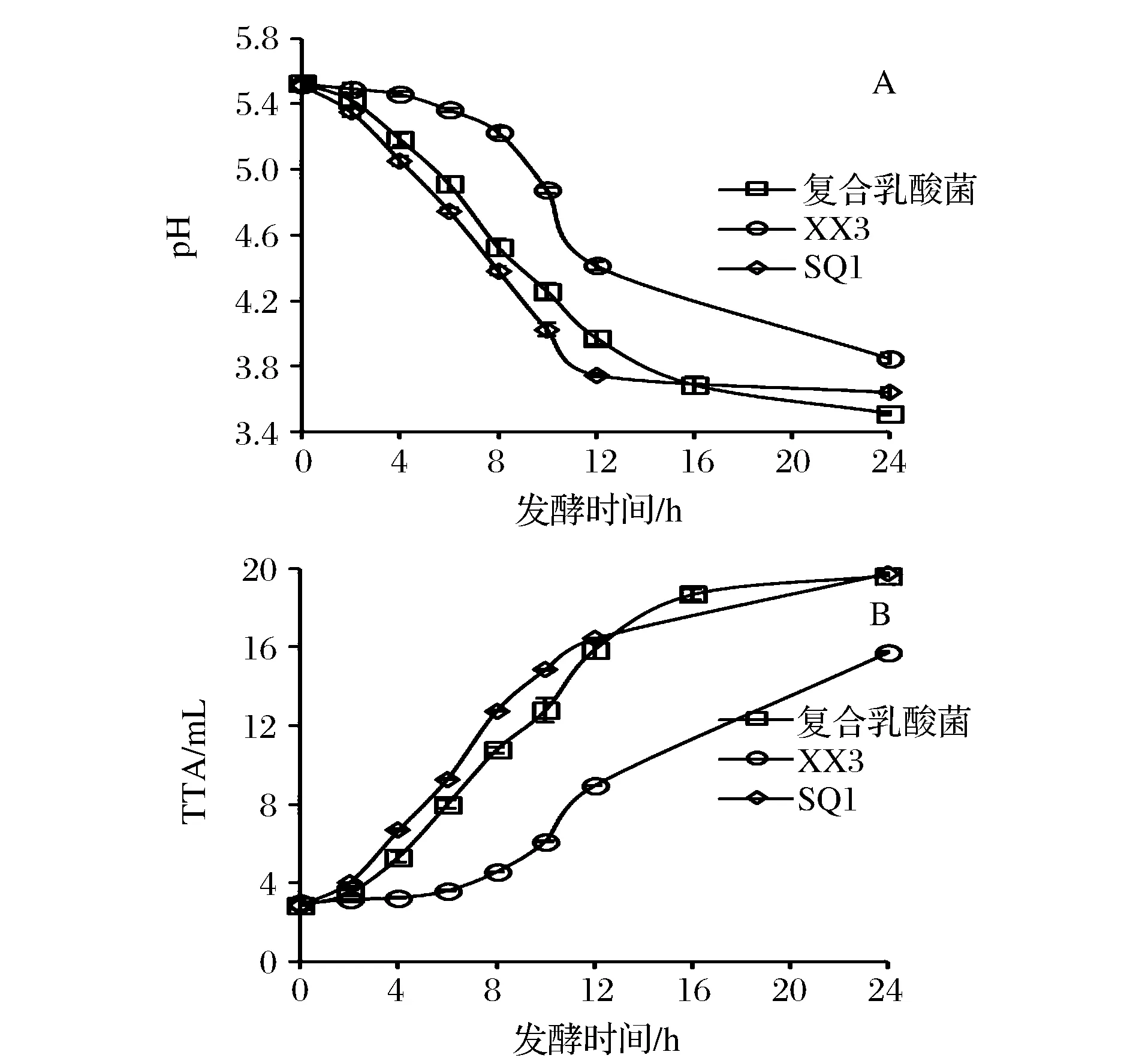
图3 复合乳酸菌面团发酵过程中pH和TTA的变化Fig.3 pH value and TTA profile of biscuit fermented by complex LAB strains
面团pH值随发酵时间的延长而不断下降,发酵24 h,面团pH由pH 5.53降至pH 3.51,而滴定酸度由2.86升至19.64。然而,XX3组在0~6 h发酵过程中,产酸速率较慢,这可能是由于乳酸菌XX3在面团发酵初期处于生长代谢延滞期阶段,菌种自身代谢水平较低,产酸速率较为缓慢。而当发酵时间延长至8 h时,面团中乳酸菌生长繁殖速率明显加快,产酸量也随之上升。复合乳酸菌发酵制得面团,其产酸速率与始终植物乳杆菌SQ1较为接近,而与单一菌株XX3发酵相比,复合菌株发酵面团的酸化速率则存在显著加快的现象。当发酵时间为10 h时,面团pH值已到达发酵终点4.26,且满足市售发酵饼干的实际生产需求,为其工业化应用奠定了基础。
2.2 复合乳酸菌对发酵饼干丙烯酰胺含量的影响
为了验证复合乳酸菌协同酵母发酵对降低饼干中丙烯酰胺含量的影响,采用气相质谱联用技术,通过添加不同发酵剂测定饼干中丙烯酰胺的含量,其结果如图4所示。
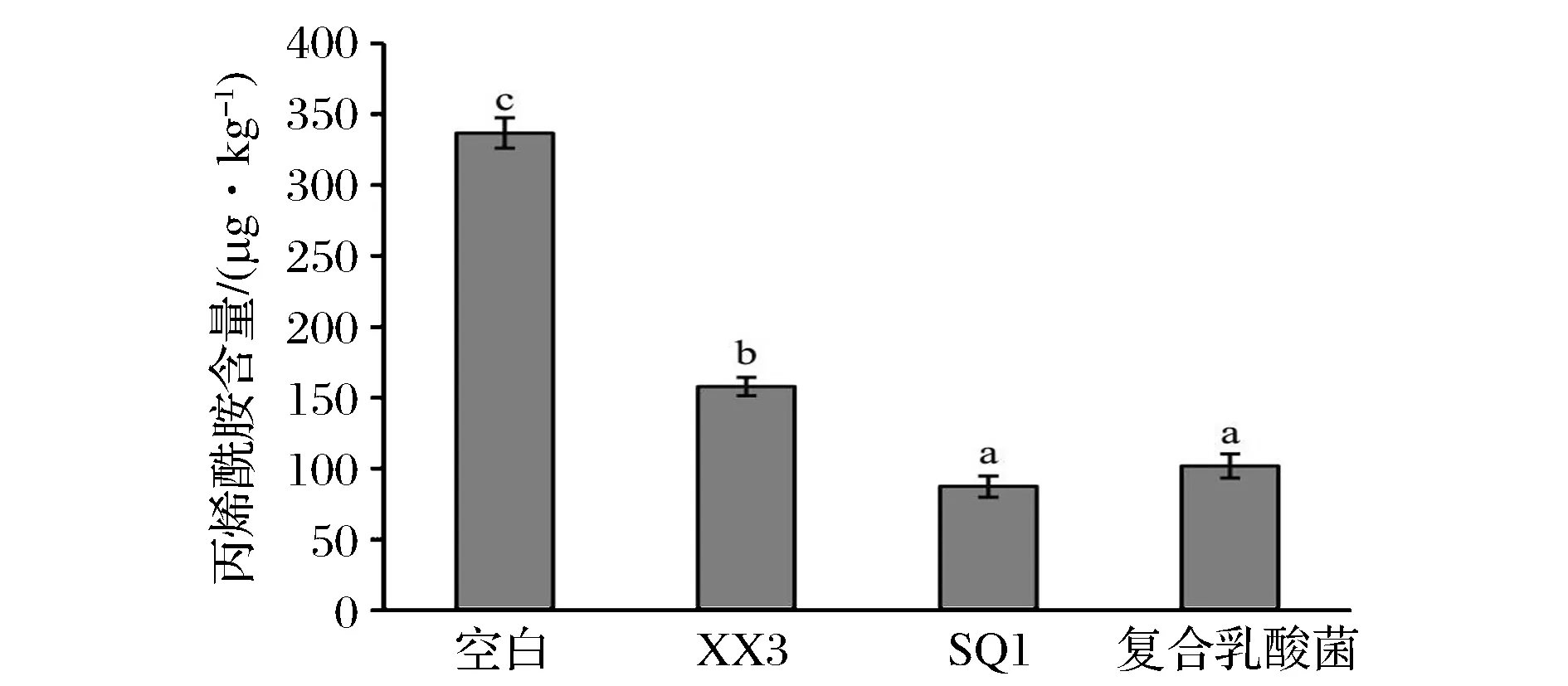
图4 复合乳酸菌对发酵饼干丙烯酰胺含量的影响Fig.4 Effect of fermented by complex LAB strains on a crylamide content of fermented biscuit
结果表明,空白组饼干中丙烯酰胺含量为336.7 μg/kg,而乳酸菌发酵组饼干中丙烯酰胺含量均呈显著降低,说明乳酸菌发酵可显著降低饼干中丙烯酰胺的含量。复合乳杆菌与植物乳杆菌SQ1发酵制备的饼干中丙烯酰胺的含量较低,效果更佳。而乳酸乳球菌XX3组中丙烯酰胺的含量则与其他两组存在显著差异。一方面,可能是由于产品在相同pH值下有机酸的组成及含量差异导致其抑制丙烯酰胺的合成能力有所不同;另一方面,研究表明乳酸菌代谢产生的某些游离氨基酸或氨基酸衍生物(如甘氨酸、蛋氨酸、GABA等)也对丙烯酰胺的形成具有抑制作用,因此,乳酸菌降低发酵饼干中丙烯酰胺含量的效果也与发酵产物中氨基酸及其衍生物的含量和构成有关,且在高温环境下,不同氨基酸与丙烯酰胺合成底物参与美拉德反应的竞争能力不同,进而导致丙烯酰胺生成量存在显著差异。
2.3 复合乳酸菌对发酵饼干描述性感官风味特征的影响
为了进一步说明复合发酵剂对饼干风味特性的影响,采用描述性感官评价方法对发酵饼干的16种风味特征的强度进行了评价分析,并对其进行消费者感官喜好度打分,其结果如图5所示。
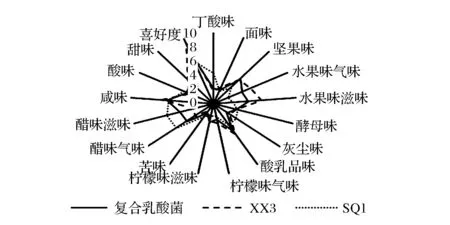
图5 复合乳酸菌发酵饼干风味特征强度及喜好度评价Fig.5 Evaluation of aroma and flavor characteristics and preference of complex LAB strains fermented biscuit
经风味特征强度及感官喜好度评价显示,复合乳酸菌制备的发酵饼干与植物乳杆菌SQ1组相比,消费者感官喜好度打分得到显著提高。其中,与喜好度呈正相关的风味特征中的水果滋气味与坚果味的强度均有显著增加,而与喜好度呈负相关的醋滋气味、丁酸味及灰尘味的强度则呈现显著减弱趋势。消费者认为复合乳酸菌组饼干中坚果及水果味,与XX3组相比较为平淡。但与SQ1组饼干相较,其风味层次丰富,酸味柔和,且具有一定的酪香味。
综上所述,采用以乳酸乳球菌XX3和植物乳杆菌SQ1为主的复合型乳酸菌发酵剂,通过酵母协同发酵方式加工制得饼干,与单一菌株发酵相比,可显著提高面团发酵过程的酸化速率,有效降低产品在加工过程中丙烯酰胺的生成量,且风味浓郁,酸味柔和,更受消费者的喜爱。因此,采用该复合型发酵剂考察其对产品营养特性的影响。
2.4 乳酸菌对发酵饼干营养特性的影响
近年来已有多位学者证实,乳酸菌不仅可改善发酵面制品的风味及质构特性,还对产品的营养功能具有一定提升作用。因此,考察复合乳酸菌发酵对饼干的消化率、维生素含量及γ-氨基丁酸含量的影响,旨在探究复合乳酸菌发酵制得饼干是否具有更佳的营养品质。
2.4.1 乳酸菌发酵对饼干蛋白消化率的影响
IVPD是重要的食品中蛋白质营养的评价指标之一,它反映了蛋白质被人体消化的程度。体外消化率越高,代表蛋白被机体利用的可能性越大。本研究测定了空白组饼干、酵母发酵饼干及复合乳酸菌发酵饼干的蛋白质体外消化率,结果见表3。研究发现,乳酸菌发酵可显著提高小麦蛋白的体外消化率。其中,在胃蛋白酶消化阶段,与未发酵样品相比,经酵母或乳酸菌发酵的面团所制备的饼干蛋白质消化率显著提高,增幅分别为9.76%和18.25%。而经胃蛋白酶和胰蛋白酶处理后,酵母发酵和乳酸菌发酵面团中小麦蛋白的体外消化率与未发酵样品相比,增幅依次为44.30%和57.20%。这主要是由于面团中乳酸菌在发酵过程中大量增殖,在乳酸菌生长代谢作用下,有机酸含量显著提高,pH值迅速下降,进而有效激活谷物内源性蛋白酶[15],促进蛋白质大分子降解成短肽和小分子氨基酸,从而有利于人体的消化和吸收。因此,IVPD的显著增加是酸与酶共同作用的结果。

表3 复合乳酸菌对发酵饼干的体外消化率的影响 %
注:不同字母标示的同一行间具有显著性差异(P≤0.05)。
2.4.2 乳酸菌发酵对饼干中维生素含量的影响
分别测定了未发酵饼干、酵母发酵饼干及复合乳酸菌发酵饼干中硫胺素、核黄素及生育酚的含量,表4显示了复合乳酸菌发酵对饼干中几种维生素的影响。经酵母菌发酵及乳酸菌发酵均能提高饼干中硫胺素及核黄素的含量,而经复合菌组发酵后制备的饼干中硫胺素及核黄素的含量最高,分别为111.88 μg/100 g和76.00 μg/100 g。这主要是由于乳酸菌长时间的发酵能够减少烘焙产品中VB1及VB2的损失[16]所导致的。而由于乳酸菌在发酵过程中酸化作用可导致面团中VE的流失[17],因此,复合乳酸菌发酵饼干中的生育酚含量显著低于空白组及酵母组饼干。LIUKKONEN等[18]在黑麦酸面团焙烤时也出现类似产品中生育酚和三烯生育酚含量降低的结果。
注:不同字母标示的同一行间具有显著性差异(P≤0.05)。
2.4.3 乳酸菌发酵对饼干中γ-氨基丁酸含量的影响
诸多学者已致力于利用筛选的乳酸菌提高食品中的GABA含量[19-20]。本研究测定了未经发酵、酵母发酵和乳酸菌发酵制备的饼干中GABA的含量,结果如表5所示。面团经酵母发酵后,其制备的饼干中GABA含量与未发酵饼干无显著变化,而复合乳酸菌发酵制备的饼干中GABA含量则为1.50 g/100 g,与未发酵饼干及酵母发酵饼干相比分别提高了78.0%和84.8%。BHANWAR等[21]发现将荞麦、苋菜、鹰嘴豆及藜麦的混合粉制成的酸面团在优化条件下发酵,植物乳杆菌C48和乳酸乳球菌乳酸亚种PU1被作为这些谷物、“伪谷物”及豆类混合物的酸面团发酵剂。混合物经酸面团发酵后其中GABA浓度(504 mg/kg)显著高于烘焙酵母发酵的混合物面团。上述结果与本研究中是一致的,乳酸菌提高发酵饼干中GABA含量的原因可能是经乳酸菌发酵制备的饼干中,由面团体系中内源或乳酸菌生物合成的谷氨酸脱羧酶可经α脱羧反应一步催化L-谷氨酸酯形成GABA[22]。

表5 复合乳酸菌对发酵饼干中GABA含量的影响
注:不同字母标示的同一行间具有显著性差异(P≤ 0.05)。
综上所述,复合乳酸菌可显著提高苏打饼干的蛋白质消化率、VB1与VB2含量及GABA含量,有助于发酵饼干中功能营养特性的提高。
3 结论
利用复合乳酸菌协同酵母菌发酵制备苏打饼干,观察其对产品品质和营养特性的影响。结果表明,复合乳酸菌在发酵过程中产酸速率与植物乳杆菌SQ1相近,发酵10 h即可满足产品面团的酸度需求,且可有效降低产品中丙烯酰胺的含量。此外,与植物乳杆菌SQ1组相较,复合乳酸菌发酵制得苏打饼干感官喜好度和描述性感官风味评价均得到了显著提升。通过对饼干的蛋白质消化率、维生素和γ氨基丁酸含量的比较发现,经复合乳酸菌发酵可显著提高苏打饼干的蛋白质消化率、VB1与VB2含量及GABA含量,其中体外蛋白质消化率为(81.54±1.46)%,VB1含量为(111.88±2.49) μg/100 g,VB2含量为(76.00±1.80) μg/100 g,GABA含量为(1.50±0.13) g/100 g,有助于苏打饼干中营养特性的提升。
[1] 饼干. GB/T 20980—2007[S].
[2] MARKETLINE. Global biscuits industry profile[M]. New York: Alalra Store, 2014:20-21.
[3] 张均叶,赵建新,闫博文,等.乳酸菌发酵降低饼干中丙烯酰胺含量机理的初步探究[J].现代食品科技,2015,31(12):277-282.
[4] 周家春.乳酸菌菌种的简便分离和培养[J].食品科学,1998,19(1): 39-41.
[5] PEDERSEN L,KAACK K,BERGSOE M N,et al. Effects of chemical and enzymatic modification on dough rheology and biscuit characteristics[J].Journal of Food Science,2005,70(2): E152-E158.
[6] 淀粉及其衍生物酸度测定:GB/T 22427.9—2008.[S].
[7] 食品中丙烯酰胺含量的测定方法气相色谱-质谱 (GC-MS)法:GB/T 5009.204-2005.[S].
[8] HAASE N U,GROTHE K H,MATTHAUS B,et al.Acrylamide formation and antioxidant level in biscuits related to recipe and baking[J]. Food Additives and Contaminants Part A,Chemistry,Analysis,Control,Exposure and Risk Assessment,2012,29 (8):1 230-1 238.
[9] 张庆.植物乳杆菌燕麦酸面团发酵过程及其面包烘焙特性研究[D].无锡:江南大学,2012.
[10] LEPORATI A,CATELLANI D,SUMAN M,et al.Application of a liquid chromatography tandem mass spectrometry method to the analysis of water-soluble vitamins in Italian pasta[J]. Analytica Chimica Acta,2005,531(1): 87-95.
[11] MIHHALEVSKI A,NISAMEDTINOV I,HALVIN K,et al.Stability of B-complex vitamins and dietary fiber during rye sourdough bread production[J]. Journal of Cereal Science,2013,57 (1): 30-38.
[12] 食品中维生素A和维生素E的测定:GB/T 5009.82-2003.[S].
[13] LV Y,ZHANG H,MENG X,et al.A Validated HPLC method for the determination of GABA by pre-column derivatization with 2,4-dinitrofluorodinitrobenzene and its application to plant GAD activity study[J]. Analytical Letters,2010,43(17):2 663-2 671.
[14] LOTONG V,CHAMBER IV2 E,CHAMBER D H.Determination of the sensory attributes of wheat sourdough bread[J].Journal of Sensory Studies,2000,15: 309-326.
[15] TURK M,CAELSSON N-G,SANDBERG A-S. Reduction in the levels of phytate during wholemeal bread making; effect of yeast and wheat phytases[J]. Journal of Cereal Science,1996,23(3):257-264.
[16] BATIFOULIER F,VERNY M A,CHANLIAUD E,et al. Effect of different breadmaking methods on thiamine, riboflavin and pyridoxine contents of wheat bread[J]. Journal of Cereal Science,2005,42 (1):101-108.
[17] WENNERMARK B H,JEGERSTAD M. Breadmaking and storage of various wheat fractions affect vitamin E[J].Journal of Food Science,1992,57(5):1 205-1 209.
[18] LIUKKONEN K-H,KATINA K,WILHELMSSON A,et al. Process-induced changes on bioactive compounds in whole grain rye[J]. Proceedings of the Nutrition Society,2003,62 (1): 117-122.
[19] DI CAGNO R,MAZZACANE F,RIZZELLO C G,et al.Synthesis of gamma-aminobutyric acid (GABA) byLactobacillusplantarumDSM19463: functional grape must beverage and dermatological applications[J]. Applied Microbiology and Biotechnology,2010,86 (2): 731-741.
[20] LI H,QIU T,HUANG G,et al. Production of gamma-aminobutyric acid byLactobacillusbrevisNCL912 using fed-batch fermentation[J]. Microbial Cell Factories,2010,9: 85.
[21] BHANWAR S,BAMNIA M,GHOSH M,et al. Use ofLactococcuslactisto enrich sourdough bread with gamma-aminobutyric acid[J]. International Journal of Food Sciences and Nutrition,2013,64 (1): 77-81.
[22] CODA R,RIZZELLO C G,GOBBETTI M.Use of sourdough fermentation and pseudo-cereals and leguminous flours for the making of a functional bread enriched of gamma-aminobutyric acid (GABA)[J]. International Journal of Food Microbiology,2010,137 (2-3): 236-245.
Effect of co-culture of lactic acid bacteria and yeast on the quality characteristics of Soda biscuit
YAN Bo-wen1, ZHAO Jian-xin1*, ZHANG Jun-ye1,2, FAN Da-ming1, HAO Yu-jie1, GUAN Lu-jing1, CHEN Wei1,3, ZHANG Hao1
1(School of Food Science and Technology, JiangnanUniversity, Wuxi 214122, China) 2(Mondelez Food China, Inc, Suzhou 215126) 3(State Key Laboratory of Food Science and Technology, Jiangnan University, Wuxi 214122, China)
The co-culture of lactic acid bacteria and yeast improves the quality characteristics of soda biscuit. This paper is aimed at developing soda biscuit starter culture, which is fermented with the co-culture of lactic acid bacteria and yeast, to investigate the effect on the flavor and nutrition of soda biscuit. The results show that compared with the solo lactic acid bacteria fermentation, the acidification rates of dough fermented with the mixed-strains is faster, and the amount of acrylamide of biscuit is lower. Furthermore, it is a kind of favorite products for consumers and is suitable for industrial manufacture. The mixed-strains could also remarkably improve the rate ofin-vitroprotein digestibility, and enhance the amount of VB1, VB2and GABA, it’s good for improving the nutrition of the soda biscuit.
lactic acid bacteria; yeast; fermentation; soda-biscuit; quality; nutrition
10.13995/j.cnki.11-1802/ts.201704016
博士研究生(赵建新教授为通讯作者,E-mail:jxzhao@jiangnan.edu.cn)。
国家自然科学基金资助项目(31471721);“十二五”国家科技支撑计划项目(2014BAD04B03)
2016-06-14,改回日期:2016-11-25
