狨猴B2m基因沉默位点在细胞水平的验证
2017-06-09邓怡晨向志光滕永康刘云波
邓怡晨,张 晨,向志光,滕永康,刘云波*
(1.中国医学科学院医学实验动物研究所,北京 100021;2.北京大学生命科学学院,生物膜及膜生物工程国家重点实验室,北京大学麦戈文脑研究所,北京 100871)
研究报告
狨猴B2m基因沉默位点在细胞水平的验证
邓怡晨1,张 晨2,向志光1,滕永康1,刘云波1*
(1.中国医学科学院医学实验动物研究所,北京 100021;2.北京大学生命科学学院,生物膜及膜生物工程国家重点实验室,北京大学麦戈文脑研究所,北京 100871)
目的 在细胞水平筛选狨猴B2m基因的有效沉默靶点,并进行验证。方法 查询人源B2m验证过的有效siRNA靶位点序列,与狨猴B2m基因序列进行同源性比较,选择匹配靶点合成shRNA序列。将体外合成的2条干扰序列分别与慢病毒载体 FUGW-TDT连接,构建FUGW-TDT-shb2m干扰表达质粒,在聚乙烯亚胺(polyethylenimine,PEI)介导下转染293T细胞,转染后48h,用实时荧光定量法检测转染细胞中B2m基因mRNA的水平。结果 筛选出2个与狨猴完全同源的B2m沉默靶位点,分别位于B2mmRNA 的290~310 bp,665~685 bp;B2m两个靶点在转录水平的沉默效率分别为(46.54±7.91)%(P< 0.05)和(83.22±4.37)%(P< 0.0001),差异有显著性。结论 成功构建成FUGW-TDT-shb2m重组质粒;在细胞水平筛选得到2个有效的B2m基因沉默靶点;为后续有关介导狨猴B2m基因沉默的研究奠定了基础。
基因沉默;狨猴;β2-微球蛋白;短发夹RNA
β2-微球蛋白(beta2-microglobulin,B2M)是一种内源性低分子量血清蛋白质,广泛存在于血浆、尿液、脑脊液、唾液以及初乳中。它是人细胞表面主要组织相容性复合体Ⅰ类分子(major histocompatibility complex class I molecule,MHCI)的β链,由99个氨基酸组成,无多态性且高度保守,是CD8分子的结合区域[1]。
已有研究表明B2m基因缺陷小鼠在胸腺和外周淋巴器官几乎缺乏所有成熟的CD4-8+T细胞,导致免疫功能缺陷[2]。目前免疫缺陷疾病动物模型的研究主要是使用大鼠和小鼠作为动物平台,而非人灵长类动物模型在生物医学研究中的应用较少。与啮齿类动物相比较,非人灵长类动物与人类亲缘关系和生理生化指标更加接近,因此,研究建立非人灵长类动物模型对于相关疾病的研究有重要意义[3]。
狨猴作为一种较小的新大陆猴,与实验室内其他常用的灵长类实验动物,如狒狒、猕猴相比较,由于其体型较小(成年体重300~500 g)、易在实验室内笼养、便于实验操作、具有较高的繁殖率、对人类细胞因子或激素具有交叉反应性、具有独特的行为和认知特性,其在生物医药科学和神经科学方面引起广泛关注[4, 5]。本文从细胞水平筛选及验证狨猴B2m基因的沉默靶位点,为进一步探究B2m基因在狨猴体内的功能奠定基础。
1 材料和方法
1.1 材料
293T细胞来至ATCC细胞库;E.coliDH5α感受态细胞购自北京全式金公司;细胞培养基 DMEM 购自Hyclone;胎牛血清(FBS)、胰蛋白酶为Invitrogen-Gibco公司产品;无内毒素质粒小提试剂盒购自康为世纪公司;逆转录试剂盒购购自TaKaRa公司;PCR试剂盒购自TaKaRa公司;shRNA合成由 Invitrogen 公司提供;引物合成由北京擎科新业生物技术有限公司提供;Trizol试剂由 Invitrogen 公司提供;Xho I和Xba I 购自New England Biolabs公司。
1.2 实验方法
1.2.1 shRNA序列设计
从broad institute 网站中(http://www.broadin stitute.org/rnai/public/)查询人源B2m基因有效siRNA靶位点序列,通过与狨猴B2m基因序列进行同源比对,选择完全匹配的两条siRNA序列,设计shRNA序列[6, 7],由Invitrogen 公司合成。
1.2.2 B2m-RNA干扰慢病毒载体的构建和鉴定
慢病毒载体FUGW-TDT含有Ubi启动子(Ubiquitin promoter)调控的红色荧光蛋白(tandem dimer tomato,TDT)报告基因,可与U6启动子调控的RNA干扰片段共表达[8]。FUGW-TDT 载体经Xho I和Xba I双酶切线性化后回收,将两条shRNA分别连入载体FUGW-TDT的U6启动子下游,即构建重组质粒[9]。连接产物转化至E.coliDH5α 感受态细胞中,涂布于50 μg/mL氨苄青霉素的LB平板上,37℃培养12 h,挑取单菌落,接种至含有50 μg/mL氨苄青霉素的LB液体培养基中,37℃培养不超过16 h,进行菌液鉴定(菌液PCR及测序引物均为F: 5’ -AGGAAGATGGCTGTGAGG- 3’;R: 5’-GCCTTGTATCGTATAAGC- 3’)。选择阳性克隆菌液进行质粒抽提,经过质粒测序(北大仪器中心),将成功插入目的片段的质粒命名为FUGW-TDT-shb2m。本实验的阴性对照为未导入shRNA的FUGW-TDT空质粒。
1.2.3 细胞培养及转染
用含有10%胎牛血清的DMEM培养液,在6孔板中培养293T细胞,每孔加入5×105个细胞,置5% CO2培养箱中37℃常规培养。24 h内细胞密度达70%~80%时进行转染。在转染试剂聚乙烯亚胺(polyethylenimine,PEI)的介导下,按PEI∶质粒=3∶1的比例配制转染混合物,将重组质粒和空载质粒转进细胞[10]。实验设为重组质粒组和空载质粒组,其中,重组质粒组包括FUGW-TDT-shb2m-1和FUGW-TDT-shb2m-2两个,空载质粒组为阴性对照,每个质粒做3个复孔。48 h后观察荧光并进行流式分选,收集带有红色荧光的细胞。
1.2.4 RT-PCR法检测B2m基因mRNA表达水平
采用实时荧光定量PCR法分析各组转染细胞中B2m基因mRNA的表达情况[11]。按照 Invitrogen Trizol Reagent说明书分别提取每组细胞总RNA,然后进行逆转录反应,逆转录条件为: 37℃ 15 min,85℃ 5 s,得到的cDNA用于进行实时荧光定量PCR反应。B2m基因引物为:上游5’-GGCTATCCAGC GTACTCCAAA-3’下游5’- CACGGCAGGCATACTC ATCTT -3’,扩增长度为248 bp。内参GAPDH的引物为:上游5’- TGACTTCAACAGCGACACCCA -3’,下游5’- CACCCTGTTGCTGTAGCCAAA -3’,扩增长度为121 bp。两条引物均由primer5软件设计得来。PCR反应程序:94℃预变性5min,94℃变性30 s,60℃退火30s,72℃延伸30 s,40个循环,72℃延伸10 min。
1.3 统计学方法
采用GraphPad Prism 5.0 软件进行统计学分析,实验数据以平均值±标准差(±s)表示,组间比较采用t检验分析。以P< 0.05表示差异有显著性。
2 结果
2.1 狨猴B2m基因shDNA寡核苷酸序列
根据狨猴B2m基因序列,依照shRNA的设计原则和broad institute网站中人源B2m基因有效siRNA靶位点序列,与狨猴B2m基因序列进行同源性比较,选择完全匹配的两个靶点(图1)合成shRNA序列,共两条核苷酸序列见表1。
2.2 B2M慢病毒沉默载体构建
在靶点序列的正反向组合中间添加一个loop环结构(序列为CTCAAGAGA)以便形成shRNA发夹结构。另外在正义链模板的 5’端添加tcga,可与XhoI酶切后的粘性末端互补,在反义链模板的 5’端添加ctag,可与XbaI酶切后的粘性末端互补,使其能与空的干扰质粒连接,由此合成靶序列的oligoDNA(图2A)。之后将oligoDNA连接在U6启动子的下游,成功构建FUGW-TDT-b2m的shRNA载体(图2B)。

图1 狨猴B2m基因序列同源性比对分析Fig.1 Homology analysis of marmoset B2m gene sequence
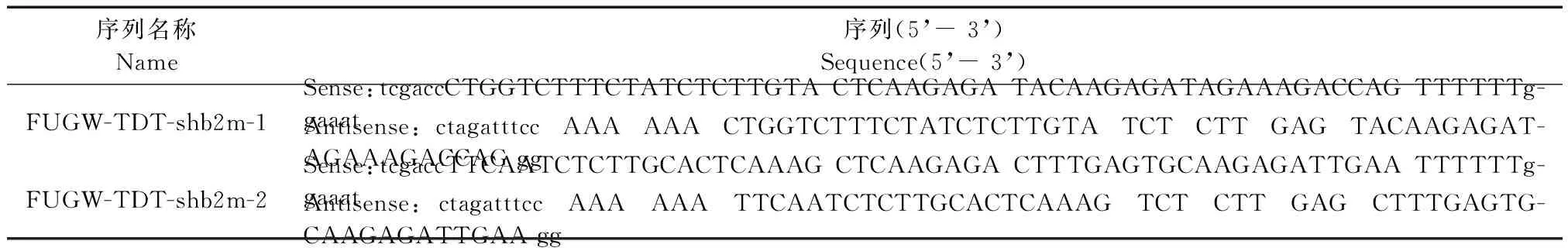
序列名称Name序列(5’-3’)Sequence(5’-3’)FUGW-TDT-shb2m-1Sense:tcgaccCTGGTCTTTCTATCTCTTGTACTCAAGAGATACAAGAGATAGAAAGACCAGTTTTTTg-gaaatAntisense:ctagatttccAAAAAACTGGTCTTTCTATCTCTTGTATCTCTTGAGTACAAGAGAT-AGAAAGACCAGggFUGW-TDT-shb2m-2Sense:tcgaccTTCAATCTCTTGCACTCAAAGCTCAAGAGACTTTGAGTGCAAGAGATTGAATTTTTTg-gaaatAntisense:ctagatttccAAAAAATTCAATCTCTTGCACTCAAAGTCTCTTGAGCTTTGAGTG-CAAGAGATTGAAgg
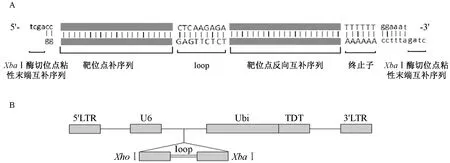
注:(A)shRNA-1的双链寡核苷酸DNA序列结构;(B)FUGW-TDT-shb2m慢病毒载体。图2 结构示意图Note.(A)Double-stranded DNA oligonucleotide sequence structure of shRNA-1;(B)FUGW-TDT-shb2m lentiviral vector.Fig.2 The frame of FUGW-TDT-shRNA
2.3 293T细胞中B2M丰度的检测
选取293T细胞提取总RNA后逆转录为cDNA,经普通PCR,电泳检测细胞中B2m基因的丰度(图3)。结果显示293T细胞中B2m基因的表达,说明能够选用293T细胞于基因抑制效率检测。

注:(M)Marker;(1)293T。图3 B2m含量检测Note.(M)Marker;(1)293T.Fig.3 Detection of B2m abundance
2.4 慢病毒表达载体FUGW-TDT的XhoI和XbaI双酶切结果
将慢病毒表达载体FUGW-TDT经XhoI和XbaI双酶切后,产物于1%的琼脂糖凝胶电泳,结果(如图4)表明:酶切片段大小10600 bp左右,与预期相符合。然后将其酶切产物采用胶纯化回收试剂盒回收目的片段,用于后续的连接反应。
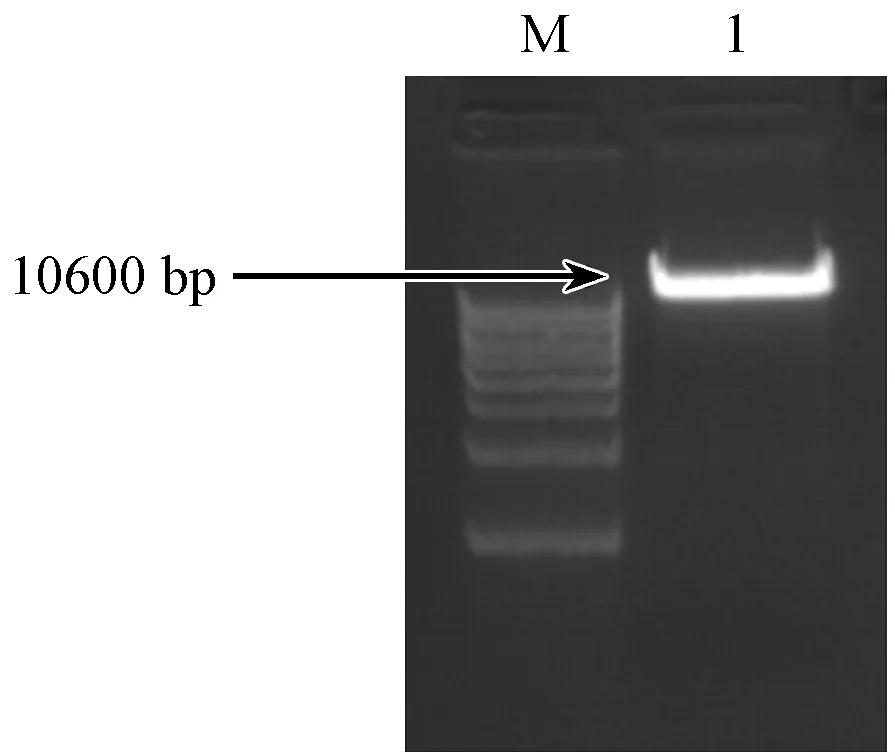
注:(M)DNAMarker;(1)经XhoI和XbaI双酶切线性化后的质粒载体。图4 FUGW-TDT载体的双酶切电泳结果Note.(M)DNA Marker;(1)Linearized plasmid XhoI and XbaI double digestion.Fig.4 Electrophoresis for production of FUGW-TDT vector by double digestion
2.5 菌液PCR鉴定阳性克隆
B2m基因shRNA的寡核苷酸序列,经退火形成双链DNA,与经XhoI和XbaI双酶切后的FUGW-TDT载体连接,连接产物转化大肠杆菌DH5α,每个平板挑取3个重组阳性克隆,进行菌液PCR鉴定阳性克隆。重组细菌克隆的PCR产物351 bp(插入片段为61 bp),而经双酶切后没有插入片段的FUGW-TDT空载体PCR产物为290 bp。作为对照(图5),鉴定结果与预期相符。证明B2m基因的shRNA已经定向连入慢病毒表达载体中。
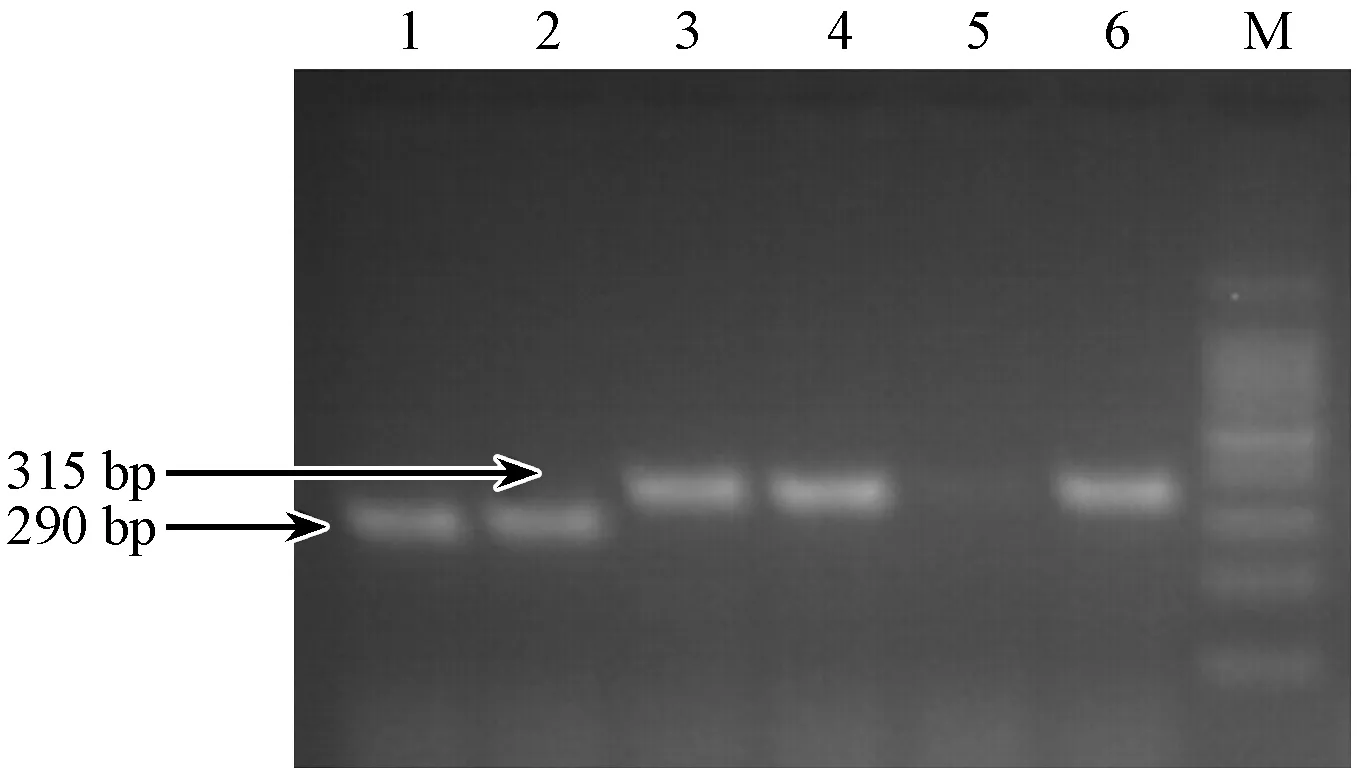
注:(M)DNAMarker;(1~3)FUGW-TDT-shb2m-1;(4~6)FUGW-TDT-shb2m-2。图5 FUGW-TDT-shb2m载体菌液PCR鉴定结果Note.(M)DNA Marker;(1~3)FUGW-TDT-shb2m-1;(4~6)FUGW-TDT-shb2m-2.Fig.5 Results of PCR FUGW-TDT-shb2m identification
2.6 RT-PCR检测293T细胞B2m基因的表达
RT-PCR检测结果显示,实验组中2个插入shRNA序列的重组质粒对应的B2m基因mRNA表达量都明显低于对照组,其中,FUGW-TDT-shb2m-1组与对照组相比,其沉默效率为(46.54±7.91)%(P< 0.05),FUGW-TDT-shb2m-2组与对照组相比,其沉默效率为(83.22±4.37)%(P< 0.0001),实验批数n=3,差异有显著性(图6)。
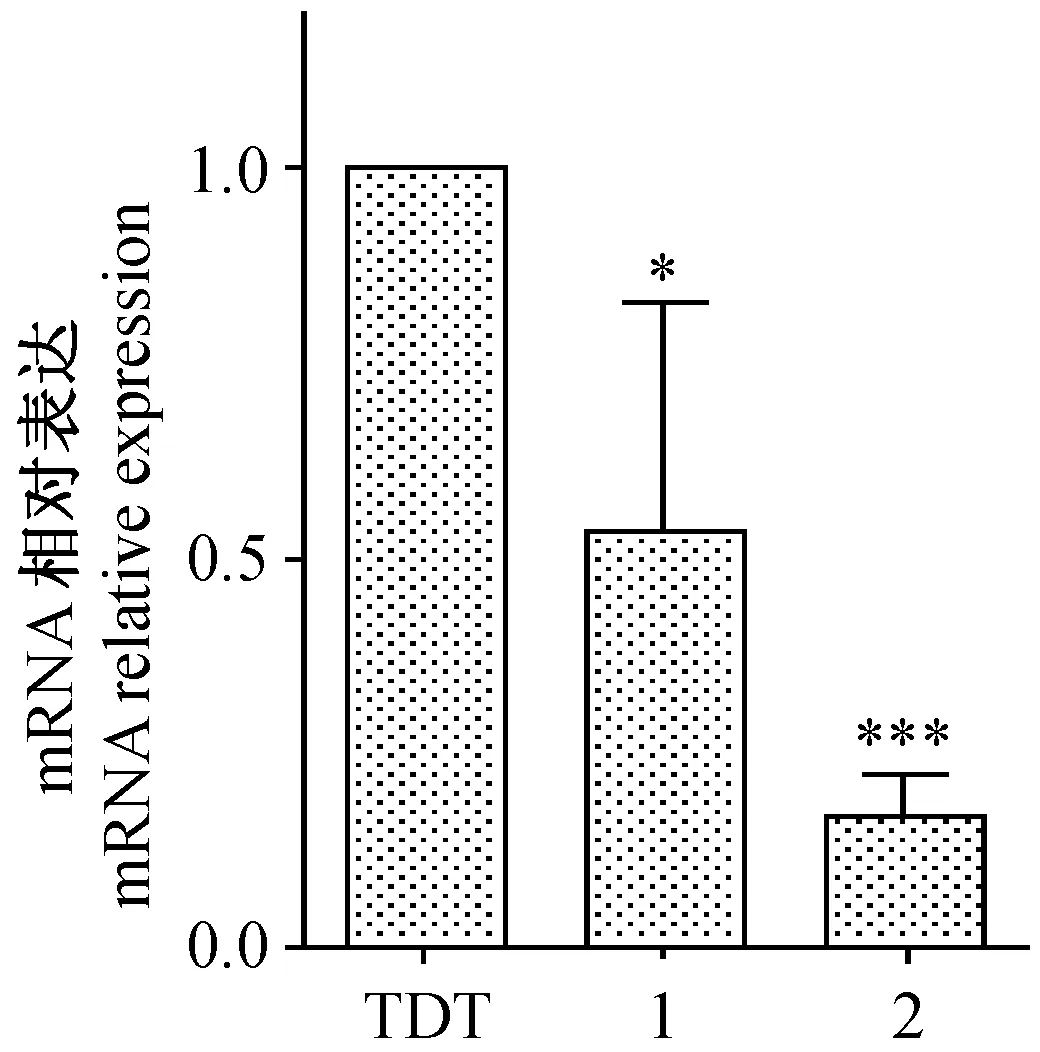
注:(TDT)对照组FUGW-TDT;(1)FUGW-TDT-B2m-1;(2)FUGW-TDT-b2m-2;与对照组FUGW-TDT相比,*P<0.05,***P< 0.0001。图6 SYBR-Green法 Real-time PCR检测B2m基因mRNA的表达(n=3)Note.(TDT)Control group FUGW-TDT;(1)FUGW-TDT-b2m-1;(2)FUGW-TDT-B2m-2; Compared with control group FUGW-TDT,*P<0.05,***P< 0.0001.Fig.6 Detection of B2m mRNA by SYBR-Green Real-time PCR
3 讨论
很多种类的免疫缺陷动物模型已经建立起来,在免疫学、遗传学、肿瘤学、自身免疫性疾病、微生物学等诸多方面发挥重要作用[12]。实验室常用的免疫缺陷动物包括裸小鼠、SCID小鼠等,但相对于啮齿类动物,非人灵长类动物在遗传、进化、生理等方面与人类高度相似,被认为是最理想的疾病治疗和预防药物研究的模型,并已广泛应用于评估抗病药物和疫苗的安全及药代工作[13, 14]。
狨猴现在已经成为重要的研制人类疾病模型的动物,在基因修饰动物研究方面有很多优势条件,比如相对成熟的辅助生殖技术和与人类进化的亲缘性。近年来,狨猴在神经科学方面及帕金森、肝炎等疾病动物模型方面具有重要地位[15]。但是, 狨猴疾病模型无法替代旧大陆猴在构建HIV、结核病等人类疾病模型的独特优势, 所以应根据课题的需求和具备的条件,采用相应的动物品种[16]。
B2m是通过非共价键与MHCI类分子的α链胞外段相互作用,不参与MHCI类分子同抗原多肽的结合,但对维持MHCI类分子天然构型的稳定性及其表达起关键作用。有文献报道B2m基因缺陷小鼠表现为免疫缺陷,且B2m基因缺陷小鼠在无病原体环境下培养,可正常发育及繁殖[17],为探究狨猴B2m基因沉默后是否有免疫降低甚至缺陷的现象,首先从细胞水平上筛选沉默效率高的B2m基因靶点。本文中,将筛选的B2m基因沉默靶点插入带红色荧光的慢病毒质粒(FUGW-TDT)后转染HEK293T细胞,通过real-timePCR方法分析,结果表明在RNA干扰作用下,转染的细胞中B2m基因在转录水平比对照组显著降低,说明RNA干扰靶点有效,可以经过病毒包装后进行狨猴胚胎和个体水平上验证实验,为构建狨猴免疫缺陷模型奠定基础。
[1] Chiou SJ,Chen CH. Decipher beta2-microglobulin: gain- or loss-of-function (a mini-review)[J]. Med Sci Monit Basic Res, 2013, 19: 271-273.
[2] Koller BH,Marrack P,Kappler JW,etal. Normal development of mice deficient in beta 2M, MHC class I proteins, and CD8+ T cells. 1990[J]. J Immunol, 2010, 184(9): 4592-4595.
[3] Van Rompay KK. Evaluation of antiretrovirals in animal models of HIV infection[J]. Antiviral Res, 2010, 85(1): 159-175.
[4] Sasaki E. Prospects for genetically modified non-human primate models, including the common marmoset[J]. Neurosci Res, 2015, 93: 110-115.
[5] Kishi N,Sato K,Sasaki E,etal. Common marmoset as a new model animal for neuroscience research and genome editing technology[J]. Dev Growth Differ, 2014, 56(1): 53-62.
[6] Grinev VV. [Design and quality control of short interfering RNAs][J]. Mol Biol (Mosk), 2012, 46(6): 827-845.
[7] Zhao H,Zhang JY,Xu WH,etal. [Construction and identification of lentiviral vector for RNA interference targeting STUB1 gene][J]. Zhejiang Da Xue Xue Bao Yi Xue Ban, 2010, 39(6): 623-627.
[8] Cheng L,Shu R,Song ZC,etal. [Reconstruction and expression of recombinant lentiviral vector for human amelogenin in 293T cell line][J]. Shanghai Kou Qiang Yi Xue, 2008, 17(1): 45-50.
[9] 苏静芬,张晨,刘云波, 等.恒河猴p53基因沉默靶点在细胞水平的验证[J].中国比较医学杂志, 2014, 24(8): 7-10.
[10] Longo PA,Kavran JM,Kim MS,etal.Transient mammalian cell transfection with polyethylenimine (PEI)[J].Methods Enzymol, 2013, 529: 227-240.
[11] Alfonso J,Pollevick GD,Castensson A,etal. Analysis of gene expression in the rat hippocampus using Real Time PCR reveals high inter-individual variation in mRNA expression levels[J].J Neurosci Res, 2002, 67(2): 225-234.
[12] Mosier DE. Small animal models for acquired immune deficiency syndrome (AIDS) research[J]. Lab Anim Sci,1996, 46(3): 257-265.
[13] Schiel N,Souto A.The common marmoset: An overview of its natural history, ecology and behavior[J].Dev Neurobiol, 2017,77(3):244-262.
[14] Watakabe A,Sadakane O,Hata K,etal. Application of viral vectors to the study of neural connectivities and neural circuits in the marmoset brain[J].Dev Neurobiol,2017,77(3):354-372.
[15] Hashikawa T,Nakatomi R,Iriki A. Current models of the marmoset brain[J].Neurosci Res,2015, 93: 116-127.
[16] Chan AW.Progress and prospects for genetic modification of nonhuman primate models in biomedical research[J].ILAR J,2013, 54(2): 211-223.
[17] Bernal M,Ruiz-Cabello F,Concha A,etal. Implication of the beta2-microglobulin gene in the generation of tumor escape phenotypes[J]. Cancer Immunol Immunother, 2012, 61(9): 1359-1371.
Validation of the silencing site of marmosetB2mgene at the cellular level
DENG Yi-chen1,ZHANG Chen2,XIANG Zhi-guang1,TENG Yong-kang1,LIU Yun-bo1*
(1.Institude of Medical Laboratory Animal Science,Chinese Academy of Medical Sciences, Beijing 100021,China;2.State Key Laboratory of Membrane Biology, School of Life Sciences; PKU-IDG/McGovern Institute for Brain Research, Peking University,Beijing 100871)
Objective To screen and determine the effective silencing targets of β2- microglobulin(B2m)gene at the cellular level in marmoset.Methods By homology comparison of the b2m gene in human and theB2mgene in marmoset, choose homology small hairpin RNA(shRNA)sequences targeting marmosetB2mgene were designed, We choose homology small hairpin RNA(shRNA)sequences targeting designedB2mgene to make homology analysis, and insert into lentivirus-based gene silencing constructs FUGW-TDT. The vectors were transfected into HEK293T cells induced by polyethylenimine(PEI). The suppression ofB2mmRNA was detected by real-time PCR. Results Two gene-silencing sequences were screened that lied in 290~310 bp and 665~685 bp of the marmosetB2mmRNA, and have statistical significance in the silencing rate:(46.54±7.91)% (P< 0.05) and(83.22±4.37)%(P< 0.0001). Conclusions Two effective silencing target sequences are screened at cellular level, which can be further used in studies on gene silencing in marmoset.
Gene silence; Marmoset;B2m; shRNA
国家科技支撑计划(2014BAI03B01)。
邓怡晨(1991-),女,硕士研究生,研究方向:实验动物学。E-mail: yichendeng007@126.com
刘云波,教授,研究方向:实验动物质量控制。E-mail: yunboliu@126.com
R-33
A
1671-7856(2017) 05-0037-05
10.3969.j.issn.1671-7856. 2017.05.010
2016-12-30
