四对结核分枝杆菌毒素-抗毒素系统基因功能的初步研究
2017-06-09刘静仪贾俊楠李卫民张俊杰高基民
刘静仪,贾俊楠,李卫民,张俊杰,高基民
四对结核分枝杆菌毒素-抗毒素系统基因功能的初步研究
刘静仪1,贾俊楠2,李卫民2,张俊杰3,高基民1
目的 探讨结核分枝杆菌毒素-抗毒素(toxin-antitoxin,TA)系统中的4对基因的功能,为研究结核分枝杆菌的传播机制提供基础科学数据。方法 选取结核分枝杆菌TA系统VapBC家族中4对基因,包括VapC4个毒素基因(Rv1720c、Rv2103c、Rv2494和Rv3408)和VapB4个抗毒素基因(Rv1721c、Rv2104c、Rv2493、Rv3407),在大肠杆菌和耻垢分枝杆菌中分别构建严谨型阿拉伯糖诱导质粒体系及乙酰胺诱导穿梭质粒体系观察VapC对细菌生长的抑制作用,及其对应的VapB解除抑制作用。结果 在大肠杆菌和耻垢分枝杆菌的实验结果一致,Rv2103c具有一定抑制作用和 Rv2104c具有消除抑制作用,其余3对毒素-抗毒素基因未见到明显的对细菌生长的抑制和消除抑制的作用。结论 成功构建了VapBC家族在大肠杆菌及耻垢分枝杆菌中的表达体系,并发现了一对TA系统基因对细菌生长的抑制和消除抑制的作用。
结核分枝杆菌;毒素-抗毒素系统;VapBC家族
结核病(tuberculosis,TB),主要由结核分枝杆菌(Mycobacteriumtuberculosis,MTB)引起。据WHO报道,2015年全球约140万人死于结核病,新发病例约1 040万。由此可见,结核病仍是威胁人类健康的重大传染性疾病。另一方面,随着结核病与HIV等免疫缺陷性疾病的并发、耐多药结核病(multidrug-resistant TB,MDR-TB)及广泛耐药结核病(extensively-resistant TB,XDR-TB)的不断增加,对于结核病的诊断及治疗都陷入了瓶颈。结合多学科开展新方向研究势在必行。
全基因组测序技术迅猛发展,已经形成了一个巨大的数据库。充分利用该数据库资源,筛选出有意义的功能基因,从而更有针对性地进行生物学验证,以加快对生命过程的理解速度,已经成为目前研究的热点。毒素-抗毒素系统(toxin-antitoxin system,TAs)最早发现于某些低拷贝质粒,通过分裂后致死效应(the postsegregation killing effect)保证其在细菌中的稳定遗传[1]。其主要组成包括一个稳定的毒素蛋白(toxin)和一个不稳定的抗毒素蛋白(antitoxin),抗毒素蛋白通过与毒素蛋白结合形成复合物,抑制其活性。随着深入研究,有证据显示TAs 在应激状态下并非完全的介导细菌程序性细胞死亡(programmed cell death,PCD)杀灭细菌,而更倾向于抑制细菌生长[2-3]。在饥饿、低氧等应激条件下,结核分枝杆菌可自发的进入生长停滞,临床表现为较长时间的潜伏期,并且现有抗结核药物不能快速有效的消灭休眠状态的结核分枝杆菌。当机体免疫力下降(如营养不良、HIV感染等),休眠结核分枝杆菌可以重新增殖,产生新的传播。因此以结核分枝杆菌的TAs为出发点,研究结核病的休眠机制对探讨某些广泛流行谱系或菌株为何具有高致病性与强传播力非常重要。
结核分枝杆菌中确认或疑似TAs共计79个,涵盖多个家族,包括VapBC(50)、MazEF(10)、YefM/YoeB(1)、RelBE(2)、HigBA(2)、ParDE(2)等[4]。TypeⅡTAs作为结核分枝杆菌TAs中最大的家族[5-6],包括VapBC、MazEF、HigBA、RelBE、ParDE等。国内外文献对MazEF、RelBE的报道较多,但对VapBC的系统研究较少。随着全基因组序列分析在结核分枝杆菌研究中的广泛推广,通过生物信息学分析得到36对可能有功能的TAs基因[7],并有一部分基因的功能在大肠杆菌或结核分枝杆菌中得到了验证,其中VapBC家族的部分毒素基因仅在大肠杆菌中具有抑菌功能,另有一部分在耻垢分枝杆菌中验证为抑菌性[8]。但是联合生物信息学方法筛选可能与谱系传播能力有关的TAs,并在大肠杆菌和耻垢分枝杆菌中构建诱导表达体系验证的工作报道较少。
Supply[9]采用全基因组序列分析在全球范围流行的北京谱系M.TB,结果提示VapC47(Rv3408)、VapC37(Rv2103)具有强烈分化;同时,根据前期工作基础,使用生物信息学研究方法,即proven评分及蛋白空间预测[10],筛选出一个可能与北京谱系传播相关的toxin基因——Rv2494。因此我们尝试在大肠杆菌和耻垢分枝杆菌中同时构建诱导表达系统,以初步鉴定所选取的四个基因是否具有毒素功能。并进一步建立有效的蛋白表达体系。以期为筛选更多谱系相关TAS建立生物信息学与分子生物学研究基础。
1 材料与方法
1.1 材料
1.1.1 菌株和培养基 大肠杆菌E.coliDH5ɑ、BL21(DE3)、BL(DE3)plysS和BW25113感受态细胞分别接种于LB和M9培养基。耻垢分枝杆菌MC2155接种于7H9液体培养基(涂布于7H10固体培养基);大肠杆菌中选用质粒pBAD33、pET 21cc、pET28a,耻垢分枝杆菌中选用质粒pACE、pMV261(均由北京师范大学张俊杰教授课题组惠赠);培养基的抗生素工作浓度分别为,在大肠杆菌中,氨苄青霉素100 u/mL,氯霉素100 u/mL,卡那霉素50 u/mL,潮霉素150 u/mL;耻垢分枝杆菌中,卡那霉素25 u/mL,潮霉素50 u/mL。
1.1.2 主要试剂和仪器HindⅢ、XbaⅠ、NdeⅠ、BamHⅠ、ClaⅠ、EcoRⅠ、BglⅡ等限制性内切酶及T4连接酶购自北京全式金生物技术有限公司;PCR产物纯化试剂盒、DNA胶回收试剂盒、质粒小提试剂盒及包涵体纯化试剂盒均购自北京康为世纪生物科技有限公司。电穿孔仪选用Bio-rad Gene Pulser Xcell。
1.2 方法
1.2.1 VapC与VapB同源蛋白毒性与抗毒性的鉴定
1.2.1.1 大肠杆菌 将4个毒素基因(VapC12、VapC37、VapC38、VapC47)分别克隆入pBAD 33载体[11],转化入E.coliBW25113,将测序鉴定为阳性克隆的pBAD33-VapC工程菌,均匀涂布于含有或不含有0.2%阿拉伯糖溶液的M9固体培养基(氯霉素100 u/mL),37 ℃培养过夜,观察菌体生长情况;将4个毒素-抗毒素共表达基因(VapBC12、VapBC37、VapBC38、VapBC47)分别克隆入pET21cc载体,转化入E.coliBL21(DE),将测序鉴定为阳性克隆的pET21cc-VapBC工程菌,均匀涂布于含有或不含有终浓度为1mM IPTG的LB固体培养基(氨苄青霉素100 u/mL),37 ℃培养过夜,观察菌体生长情况。
1.2.1.2 耻垢分枝杆菌 将4个毒素基因(VapC12、VapC37、VapC38、VapC47)分别克隆入pACE穿梭载体[12],电转入耻垢分枝杆菌感受态(电转条件为2 500 V,25 μF,1 000 Ω,2mm)[13],将测序鉴定为阳性克隆的pACE-VapC工程菌,均匀涂布于含有或不含有0.1%乙酰胺溶液的7H10固体培养基(潮霉素50 u/mL),37 ℃培养2~3 d,观察菌体生长情况;随后,构建带有4个抗毒素基因(VapB12、VapB37、VapB38、VapB47)的pMV261-VapB穿梭质粒,并电转入由上述同源pACE-VapC工程菌制备而成的感受态细胞中,最后将同时带有pACE-VapC与pMV261-VapB的工程菌均匀涂布于含有或不含0.1%乙酰胺的7H10固体培养基(潮霉素50 u/mL、卡那霉素25 u/mL双抗),37 ℃培养2~3 d,观察菌体生长情况。
1.2.2 Rv2103c的蛋白表达与纯化
1.2.2.1 pET28a-Rv2103c重组质粒的构建 将Rv2103c毒素基因克隆入原核表达质粒pET-28a,转化入E.coliBL21(DE)plysS感受态细胞,经T7通用引物测序鉴定为阳性克隆的工程菌重命名为pET28a-Rv2103c。
1.2.2.2 Rv2103c的蛋白表达 将构建成功的pET28a-Rv2103c工程菌以1∶100的比例接种于1 000 mL LB液体培养基(卡那霉素50 u/mL),37 ℃剧烈振荡培养至OD=0.4-0.6,加入终浓度为0.4 mmol/L的ITPG,继续振荡培养8 h,4 ℃ 5 000 r/min离心收菌。
1.2.2.3 Rv2103c的蛋白纯化 将菌体沉淀超声破菌后离心收集沉淀,利用康为世纪his标签包涵体蛋白纯化试剂盒洗脱杂蛋白,回收目的蛋白。
2 结 果
2.1 重组质粒的构建与鉴定 以M.TB标准菌株H37Rv基因组DNA为模板,扩增带有不同酶切位点的VapC、VapB基因目的片段,将扩增产物于2%琼脂糖凝胶电泳鉴定(见图1);挑取转化成功的单菌落扩大培养,以上游与下游引物做菌落PCR鉴定,有目的条带的样品送予北京擎科新业生物技术有限公司测序鉴定(见图2)。

M:100 bp Plus DNA Ladder; 1: Rv1720c (390 bp); 2: Rv2103c (435 bp); 3: Rv2494 (426 bp); 4: Rv3408 (411 bp).图1 toxin基因PCR扩增的目的基因片段Fig.1 PCR amplification of the target toxin gene

图2 pET21cc-VapBC12重组质粒测序结果(Clone Mgr Suite 7)Fig.2 Sequencing result of recombinant plasmid pET21cc-VapBC2(Clone Mgr Suite 7)
2.2 毒素蛋白对大肠杆菌生长的抑制作用 将4个毒素基因分别克隆入pBAD33质粒,转化E.coliBW25113,分别在含有和不含有0.2%阿拉伯糖诱导剂的M9固体培养基上均匀涂布,37 ℃培养过夜。可见pBAD 33-Rv2103c在含有0.2%阿拉伯糖的M9固体培养基上完全抑制菌落生长。在不含有0.2%阿拉伯糖的M9固体培养基上能够正常生长,且未见与pBAD 33对照组菌落存在差异。同时,其抗毒素基因Rv2104c对此种抑菌作用有解除作用(见图3)。

左1,左2:涂布顺序;右1:VapC毒素基因的诱导验证;右2:VapBC毒素-抗毒素基因的诱导验证Left is the diagrammatic presentation,right is the identification of VapC and VapBC,respectively.图3 大肠杆菌中VapC与VapB同源蛋白的毒素与抗毒素功能的鉴定Fig.3 Identification of the functions of toxin VapC and homologous antitoxin VapB in E. coli

左1,左2:涂布顺序;右1:VapC毒素基因的诱导验证;右2:VapB抗毒素基因的诱导验证Left is the diagrammatic presentation,right is the identification of VapC and VapB,respectively.图4 耻垢分枝杆菌中VapC与VapB同源蛋白的毒素与抗毒素功能的鉴定Fig.4 Identification of toxin VapC and homologous antitoxin VapB in M. smegmatis
2.3 毒素蛋白对耻垢分枝杆菌生长的抑制作用 将4个毒素基因分别克隆入pACE质粒,抗毒素基因分别克隆入pMV 261质粒,转化入耻垢分枝杆菌感受态。涂布于含有或不含有0.1%乙酰胺溶液的7H10固体培养基,37 ℃倒置培养2~3 d,观察菌体生长情况(见图4)。可见,在耻垢分枝杆菌中,Rv2103c仍具有毒素作用,其抗毒素基因Rv2104c具有一定抑制作用。
2.4 Rv2103c的表达与纯化 ITPG诱导pET28a-Rv2103c重组菌表达后收菌,将超声破碎后离心所得的上清与沉淀分别SDS-PAGE电泳(见图5)。结果显示,沉淀中与目的蛋白(约15.95 kD)大小接近的位置有一明显条带;大量表达后收集超声破碎后沉淀做包涵体蛋白纯化,得到较高纯度的目的蛋白(见图6)。

M:marker;1:the whole induced bacteria;2:supernatant after centrifugation;3:precipitation after centrifugation图5 小量诱导表达的蛋白可溶性鉴定Fig.5 Protein soluble identification by a small amount of induced expression
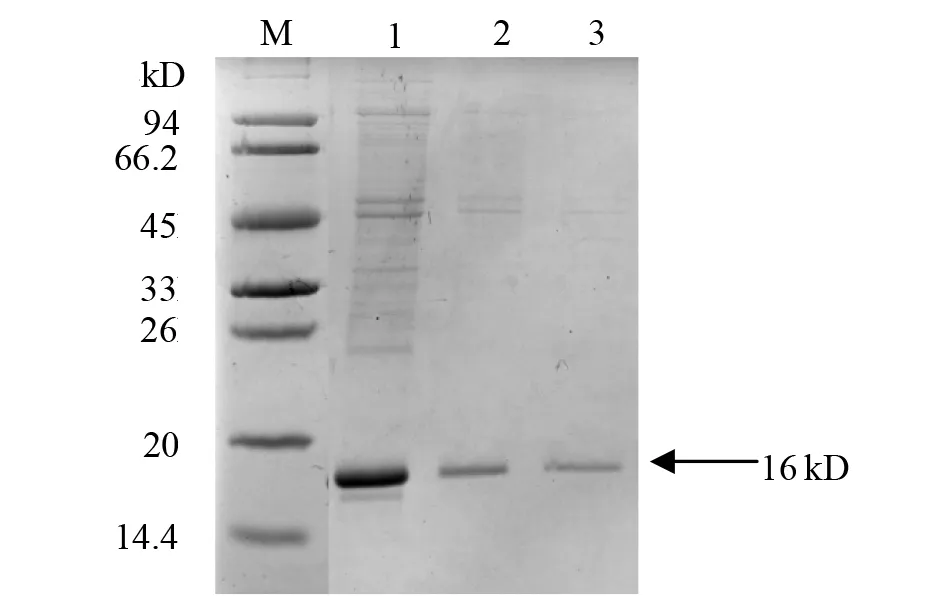
M:marker;1:supernatant after centrifugation;2、3:purification图6 大量诱导表达的蛋白纯化(包涵体)Fig.6 Protein purification by a large amount of induced expression (inclusion protein)
3 讨 论
目前的研究显示,结核分枝杆菌全基因组上共有79个可能的TAs,其中VapBC家族基因所占比例最高(50/79)[5]。前期工作中,我们通过生物信息学技术得到以谱系划分的toxin基因SNP突变位点,我们从中选取了4个具有一定谱系特征,且有不同生物信息学预测的毒素基因进行初步生物学功能验证。结果表明,仅VapC37(Rv2103c)对大肠杆菌的生长具有显著抑制作用。在所选的4个毒素基因中,VapC12(Rv1720c)在大肠杆菌中的毒素验证有过报道[10],与本文研究结果一致,即对大肠杆菌没有明显抑制作用。考虑到大肠杆菌与分枝杆菌性质上存在的差异,我们选取快生长分枝杆菌模式菌株——耻垢分枝杆菌(Mycobacteriumsmegmatis)MC2155,进一步验证所选取的4对毒素抗毒素基因是否具有抑制细菌生长的功能,结果显示仅VapC37(Rv2103c)具有明显的抑菌作用,VapC12(Rv1720c)、VapC38(Rv2494)对耻垢分枝杆菌的生长没有抑制作用,与已有实验报道相符合;但是本实验中VapC38(Rv2494)对耻垢分枝杆菌的生长无抑制作用,与已有实验报道不符[8]。在多次重复实验确认重组质粒构建正确的情况下,我们猜测产生此种差异的原因可能是选用的质粒不同。本实验所用质粒均由北京师范大学张俊杰教授课题组惠赠,参考其前期研究成果,同样存在与相关报道不相符的实验结果。例如,利用pACE构建的毒素基因诱导表达体系中,Rv1942c、Rv1495对大肠杆菌及耻垢分枝杆菌的生长均无明显抑制作用;而将毒素基因整合入pHR100质粒[8]的生物学验证实验中,Rv1942c、Rv1495对大肠杆菌及耻垢分枝杆菌的生长均有明显抑制作用。这提示我们,有必要对已有报道为有功能的毒素基因再次鉴定,以验证是否存在假阳性结果。
前期研究中(data not shown),我们通过对2 347株已知结核分枝杆菌全基因组序列与H37Rv标准株比对,得到107 654个SNP位点,利用生物信息学技术构建系统发育树可将其进一步划分为7个谱系。针对结核分枝杆菌全基因组上已标注的TAs,我们以谱系为单位,对所有79个toxin基因SNP位点进行整理归纳。其中lineage2(东亚谱系)、lineage4(欧美谱系)、lineage3所携带的非同义突变SNP远高于其他谱系。Lineage2、Lineage3、Lineage4是已知的“现代”谱系,带有更多的toxin基因突变位点,符合“现代”谱系高致病性、强传播力的特征。这提示我们探索高致病性“现代”谱系与验证为具有生物学功能的毒素抗毒素系统是否存在相关性,为结核分枝杆菌潜伏感染的检测与诊断,提供新的研究思路。
[1] Engelbergkulka H,Glaser G. Addiction modules and programmed cell death and antideath in bacterial cultures[J]. Microbiology,1999,53(53): 43-70.
[2] Gerdes K,Christensen SK,Løbner-Olesen A. Prokaryotic toxin-antitoxin stress response loci[J]. Nat Rev Microbiol,2005,3(5): 371-382.
[3] Hayes F. Toxins-antitoxins: plasmid maintenance,programmed cell death,and cell cycle arrest[J]. Science,2003,301(5639): 1496-1499.
[4] Sala A,Bordes P,Genevaux P. Multiple toxin-antitoxin systems inMycobacteriumtuberculosis[J]. Toxins,2014,6(3): 1002-1020.
[5] Georgiades K,Raoult D. Genomes of the most dangerous epidemic bacteria have a virulence repertoire characterized by fewer genes but more toxin-antitoxin modules[J]. PLoS One,2011,6(3): 1387-1387.
[6] Sala A,Bordes P,Fichant G,et al. Toxin-antitoxin loci inMycobacteriumtuberculosis[M]// Prokaryotic Toxin-Antitoxins. Berlin,Springer Berlin Heidelberg,2013: 295-314.
[7] Zaychikova MV,Zakharevich NV,Sagaidak MO,et al.Mycobacteriumtuberculosistype II toxin-antitoxin systems: genetic polymorphisms and functional properties and the possibility of their use for genotyping[J]. PLoS One,2015,10(12).
[8] Ramage HR,Connolly LE,Cox JS. Comprehensive functional analysis ofMycobacteriumtuberculosistoxin-antitoxin systems: implications for pathogenesis,stress responses,and evolution[J]. PLoS Genet,2009,5(12): 1000767.
[9] Merker M,Blin C,Mona S,et al. Evolutionary history and global spread of theMycobacteriumtuberculosisBeijing lineage[J]. Nat Genet,2015,47(3): 242-249.
[10] Li XY,Li Y,Zhang Y,et al. The epidemiological characteristics of Beijing lineageMycobacteriumtuberculosisfrom a National Referral Center in China[J]. Biomed Environ Sci,2015,28(7): 539-543.
[11] Guzman LM,Belin D,Carson MJ,et al. Tight regulation,modulation,and high-level expression by vectors containing the arabinose PBAD promoter[J]. J Bacteriol,1995,177(14): 4121-4130.
[12] Xian YB,Cui LD,Zhang JJ. Characterization of toxin-antitoxin systems inMycobacteriumtuberculosis[J]. Acta Microbiologica Sinica,2011,51(2): 214. (in Chinese)
贤一博,崔丽丹,张俊杰.结核分枝杆菌中毒素-抗毒素系统的鉴定[J].中国微生物学报,2011,51(2):214.
[13] Lu YY,Huangpu YM. High efficiency electro-genetic transformation ofMycobacteria[J]. J Beijing Med Univ,2000,32: 178-181. (in Chinese)
路艳艳,皇甫永穆.电穿孔法对分枝杆菌进行高效基因转化[J].北京大学学报医学版,2000,32(2):178-181.
Function of four pairs of genes in toxin-antitoxin system ofMycobacteriumtuberculosis
LIU Jing-yi1,JIA Jun-nan2,LI Wei-min2,ZHANG Jun-jie3,GAO Ji-min1
(1.ZhejiangProvincialKeyLaboratoryforTechnologyandApplicationofModelOrganisms,SchoolofLaboratoryMedicineandLifeScience,WenzhouMedicalUniversity,Wenzhou325035,China; 2.NationalTuberculosisClinicalLaboratoryofChina,CapitalMedicalUniversityAffiliatedBeijingChestHospital,Beijing101149,China; 3.KeyLaboratoryofCellProliferationandRegulationBiology,MinistryofEducation,SchoolofLifeSciences,BeijingNormalUniversity,Beijing100875,China)
We discussed the function of four pairs of genes in the toxin-antitoxin system ofMycobacteriumtuberculosis,providing theoretical foundation and scientific basis for studying the transmission mechanism ofMycobacteriumtuberculosis. Four pairs of genes which belong toVapBCfamily,including fourVapCgenes (Rv1720c,Rv2103c,Rv2494,Rv3408) and fourVapBgenes (Rv1721c,Rv2104c,Rv2493,Rv3407) were chosen. We constructed a serial of arabinose-induced hybrid plasmid system inEscherichiacoliand a serial of acetamide-induced hybrid plasmid system inMycobacteriumsmegmatisrespectively,in order to observe the potential inhibition effect ofVapCand the release inhibition of homologousVapB. Results showed that only one toxin gene(Rv2103c) showed the function of bacteriostasis in bothE.coliandM.smegmatisand the homologous antitoxin gene(Rv2104c) could release the inhibition of growth. We built the inducible systems ofVapBCfamily in bothE.coliandM.smegmatisrespectively and found only a pair of toxin and antitoxin genes(Rv2103c,Rv2104c) had the function of inhibition and release for the growth of bacteria. And two pairs of toxin genes(Rv1720c,Rv2494) did not have the function of inhibition for the growth of bothE.coliandM.smegmatis. Whereas,another toxin geneVapC47(Rv3408) also did not have the bacteriostastic activity,only this result was not consistent with the existing literature. We speculated that the reason for this kind of difference may be the different inducible systems we used. Cause the other three results were consistent with all existing literature and the doubtful result also appeared in other reports,so our protocol could be confirmed as reliable,and we would use it to build inducible systems and make further functional identification of certain toxin and antitoxin genes that we are interested in.
Mycobacteriumtuberculosis; toxin-antitoxin system; VapBC family
Gao Ji-min,Email: jimingao64@163.com
10.3969/j.issn.1002-2694.2017.05.005
国家自然科学基金(81273144);北京市自然科学基金B类重点项目(KZ201510025024)
高基民,Email:jimingao64@163.com
1.温州医科大学检验医学院生命科学学院浙江省模式生物技术与应用重点实验室,温州 325035; 2.首都医科大学附属北京胸科医院国家结核病临床实验室,北京 101149; 3.北京师范大学生命科学学院,教育部细胞增殖及调控生物学重点实验室,北京 100875
R378.9
A
1002-2694(2017)05-0413-05
2016-12-09 编辑:刘岱伟
Supported by the National Natural Science Foundation of China (No. 81273144) and the Beijing Natural Science Foundation Program and Scientific Research Key Program of Beijing Municipal Commission of Education (No. KZ201510025024)
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
杂志排行

中国人兽共患病学报的其它文章
- 中国西北四省(区)结核分枝杆菌分离株一线药物耐药状况及其影响因素分析
- 美国《Emerging Infectious Diseases》2017年第3期有关人兽共患病论文摘译
- MicroRNA通用探针法的建立及布鲁氏菌病患者血浆microRNA-146a的检测
- 龟头包皮炎患者淋病奈瑟菌分离与鉴定及其耐药机制研究
- Identification of non-tuberculosis mycobacteria speciesof clinical isolates from patients clinically diagnosed with tuberculosis in Fujian Province,China
- 耐万古霉素肠球菌临床菌株毒力基因携带情况及差异