中国元古宙主要生物演化事件
2017-06-07杜汝霖
杜汝霖
DU Ru-lin
河北地质大学, 河北 石家庄 050031
Hebei GEO University, Shijiazhuang, Hebei 050031
中国元古宙主要生物演化事件
杜汝霖
DU Ru-lin
河北地质大学, 河北 石家庄 050031
Hebei GEO University, Shijiazhuang, Hebei 050031
元古宙是地球地质发展历史第二阶段,在地质发展史和生物演化史上占有极重要的地位,是早期生物演化有重大转折的时代。生物演化是地质演化最灵敏的指示者,地质每次突变都会使环境、物种和生态发生相应改变事件。元古宙共经历了真核生物出现,宏体生物出现,多细胞组织分化和底栖生物出现,后生动物出现及其生物辐射爆长等事件,从而使低级、简单生物的隐生宙发展演化转折到更高级复杂繁盛显生宙的新时代。
元古宙;生物演化;桑树鞍藻;龙凤山藻
一、前言
元古宙(Proterozoic Eon)是地球地质发展历史第二阶段,距今从25亿~6亿年。它在地质发展史和生物演化史上,都占有极为重要的位置。它是早期生物演化过程中有重大转折的时代,它是有机界开始繁盛的时代,是低等菌藻类植物占统治的时代,也是后生动物开始出现的时代。生物演化是地质演化最灵敏的指示者,由于元古宙地质历史上发生过多次突变事件,每次事件都引起了地质环境的显著改变和生物物种的替换以及大范围生态系统的重建,从而形成生物演化事件一幕接一幕的演变特征,元古宙时期共有五次重大演化事件,而每一次生物演化事件都标志着一个时代的变革发生。从而促进生物面貌不断变化和向前发展。
我国是世界上元古宙化石异常丰富的国家之一,半个世纪以来,我国在元古宙生物起源和演化的研究上,取得了令世人瞩目的丰硕成果,这些最早的化石记录它极大改变和提前了人们对早期生物界起源和演化面貌的认识。
二、距今18亿~19亿年间的真核生物出现事件
在加拿大安大略省,年龄为19亿年左右的冈弗林特组硅质叠层石及燧石层中,发现了大量多样的细菌和蓝藻微生物化石,其中有些种属被一些学者认为是原核细胞与真核细胞间的过渡类型,即原始的真核细胞(见图1)。
距今18亿年前真核生物出现的较可靠的证据,来自中国燕山地区长城系底部微古植物化石。1980年代初期,罗其玲等在常州沟组或串岭沟组页岩相中(同位素年龄在1 850~1 750百万年间)发现了个体较大的梭形、三角形、片状等形态复杂、种类繁多属真核生物的藻类微体化石[1](图版I)。陈晋镳等在串岭沟组和团山子组发现了较小的宏观藻类化石残片[2],张昀对加拿大安大略省西部的冈弗林特铁建造的黑色燧石中发现了可能是世界最早出现的单细胞真核生物化石进行了描述[3](见图1)。杜汝霖、胡华斌、李培菊、裴天明等在河北宣化串岭沟组宣龙式铁矿层中找到了和加拿大冈弗林特铁建造的相当层位和相同的化石类型(均产有底栖型的丝状体和球状体微化石)和产有共同的微化石分子、冈弗林特藻(Gunflintia sp)和休仑藻(Huroniospora sp),化石体大小和直径也较为相似,同位素年龄在18亿~19亿年[4](图版II)。阎玉忠在串岭沟组发现了大量裂梭藻Schizo f usa化石,推论其为真核生物的原始种类[5]。
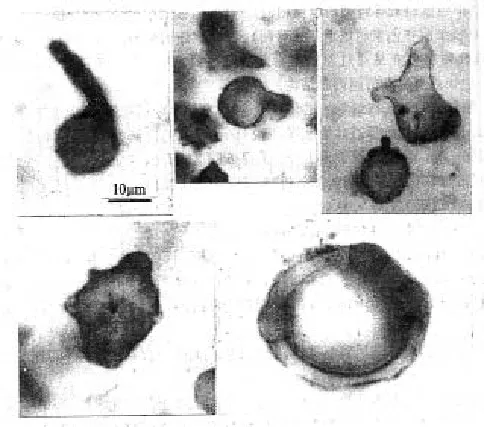
图1 中元古代早期的单细胞真核生物化石
真核细胞的出现是生物演化史上一个重大的转折和突破,因为真核生物的有性生殖会导致生殖机制的变异,不仅使生物有可能向多样化变化,还加速了生物进化的步伐,并有可能朝着机能和器官不断分化和完善的方向发展。
三、距今14亿~15亿年间的宏体生物出现的事件
自从美国古生物学家华可脱(C. D. Walcott)于1899年将发现在美国亚利桑那州的一套未变质地层中有小圆盘状炭质宏体后生植物化石命名为Chuaria circularis(丘阿尔藻)化石[6]后,在世界各地曾较普遍发现了这类化石。该化石具清晰圆形、卵圆形,边缘或有1~2个同心圆边缘带,个体大小不一,最小直径2mm,最大可达10mm,一般在5~6mm,大多具黑色炭质薄膜,其生物分类位置未定,多数学者认为是一种单细胞球形或椭球形浮游生物体压平后,在岩层中遗留下痕迹。同位素年龄时限在600 ~ 1 100百万年。在我国辽宁、河北、湖北、安徽、河南、陕西等地多有发现(见图版Ⅳ,图10~图11)。但这类宏体藻类化石一直未获较大突破。直到1983年以后中国才发现有最早较大形的化石出现。
杜汝霖、田立富等于1983年—1986年在天津蓟县桑树鞍村附近属长城系高于庄组五段下部的泥质白云岩夹碳质页岩中发现了宏体化石[7]。化石属单体,棕褐色带状含有机质薄膜,不分支。一般具两种形态,一种为近椭圆形的旋曲状,直径约8~18 mm,旋带成一圈套一圈形式,疏密不均可有1圈套5圈,旋带宽度1~2 mm,表面无纹饰,另一种为弯曲状和直线状。也有一些为上述两种的过渡形态或任意盘绕。所有带状化石体的宽度都较均匀一致,两侧边缘平行,有明显的外缘。带状体的对称性好,有时两种化石体常互相叠绕或共生。化石定名为桑树鞍藻(Sangshuania),化石同位素年龄在1 434~1 400百万年间[8]。它不仅是我国目前所知最早的宏体化石(见图版Ⅲ),而且也是目前所知世界上最早的宏体化石,它使宏观生物最早发现的时间提前了2~4亿年。
根据化石两种形态不同共分出两个新种:一个种是桑树鞍桑树鞍藻(Sangshuania sangshuanensis)旋曲状,另一种是线形桑树鞍藻(Sangshuania linearis)直线或弯曲状(均见图版Ⅲ)。该化石就其保存状态、大小、特征和时代,与瓦尔特(M. R. Walter)等1976年在北美蒙大拿洲贝尔特超群(Belt Super-group)的格里森组(Greyson Fm.)中发现和研究的大型炭质化石Grypania spiralis是很相似的[9]。化石年龄在1 450~1 400百万年间。据M. R. Walter的深入研究,认为该生物化石可能是具真核细胞的藻类实体化石,与褐藻或红藻及绿藻有关。
从中国长城系高于庄组宏体化石的丰度高、个体大、形态多样、边缘清楚等特征分析、证明该类生物已演化到相当高的水平,由此推断宏体生物的起源,至少应提前到距今15亿年以前。亚洲和北美在14亿年前形成的地层中都发现这一相同类型的宏体化石,证明了早期生物在演化阶段上的全球性和同步性。
四、8.5亿~9亿年宏观多细胞组织分化和底栖生物出现事件
1979年—1985年杜汝霖、田立富在河北省怀来县新保安镇附近的龙凤山南坡地层属于青白口系长龙山组二段下部页岩里,在与大量丘阿尔藻类化石共生中,终于找到了非常奇特的藻类化石[10]。该化石成单体、个体较大,一般在5~20mm,含量丰富,保存完好,具稳定对称和扩展型的形态特征。化石体由叶状体、似叶柄(拟茎)及假根(固着器)三部分组成(见图版Ⅳ和图2)。
叶状体呈圆盘状或卵圆状等,长度在3~15mm,似叶柄(拟茎)长度变化大,一般10~20mm,似叶柄宽度1~2mm,假根(固着器)一般成柄状或茎块状,一般大小在2~6mm。这类化石与以前国内外发现的藻类生物完全不同,从未发现,因此定名为龙凤山藻(Longfengshania),1985年当时已发现有10个属,28个种,1个新属,4个新种,因此,1986年建立了以龙凤山藻为代表,包括多种共生藻类的龙凤山生物群[11]。关于龙风山生物群的年龄时限,经同位素年龄测定为850~900百万年。
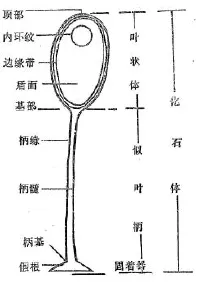
图2 化石体构造图
龙凤山藻其形态和构造与具根、茎、叶分化的高等植物很相似,并已具有一定的生理功能,如营养、繁殖和固着等功能。生物组织器官的分化,反映了细胞机构的复杂和细胞群的分工与分化。因此,可以作为宏观多细胞真核生物出现的重要证据,这是多细胞体型还表明它已具有有性生殖能力。简言之,这是多细胞化和生物组织分化水平的一次大提高,这一演化事件奠定了地球上一切高级生命产生和发展的基础。
龙凤山藻类生物不仅是最早具器官分化的宏体生物,而且也是目前所知世界最早出现的宏观底栖藻类植物。生物原植体中拟柄和假根的出现为海生植物固着、吸收和疏导营养创造了条件,是海生植物向陆生植物演化发展的决定性的一步。从龙凤山藻的复原图中可见该生物在水中保持直立的状态(见图3),这非常有利于气体交换、水分和营养的运输以及进行光合作用。因此,这种生物也非常适应向陆地演化。从此对一个光秃秃的大陆开始逐渐披上绿色的新装。需要指出的是龙凤山藻类化石除在怀来龙凤山发现外,还在河北兴隆、北京昌平、湖北秭归、云南昆明等地陆续发现了多种龙凤山藻类化石,而且还在加拿大麦肯齐山小达尔宏观生物群中发现了龙凤山藻化石[12],其层位和时代大致相近,这也再次证明生物演化阶段的全球性和同步性。
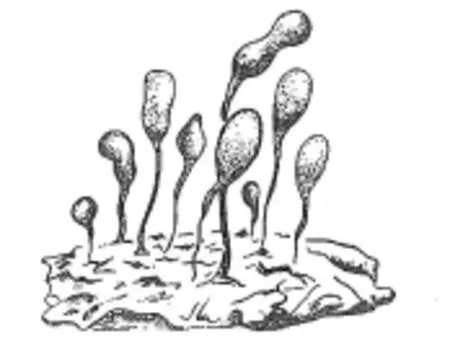
图3 龙凤山藻原植体生态复原臆想图
五、距今8.5亿~7亿年间的后生动物出现的事件
后生动物的出现是生物进化中的又一重大的事件,后生动物从什么时间开始出现,目前尚有不同的争论。近些年来,在我国华北地区的苏皖北部和东北辽南地区的震旦纪早期或更早的时期中,发现有早于7亿年的可靠的后生动物化石,这个属于前伊迪卡拉动物群,在世界其他地区还很少见,所以有其独特的意义。
郑文武等1981年—1986年在皖北淮南地区相当青白口系上部的刘老碑组和相当震旦系下部的九里桥组中发现了含量较丰富、类型多样、保存完好的蠕虫类动物化石[13]。汪贵翔对这类化石进行了深入的研究,主要分子有须腕动物门的淮南中华皱节虫、叶文金古线虫,环节动物门的震旦安徽虫、百瓜山淮远虫、小淮远虫、安徽古吻虫等。刘老碑组仅有淮南中华皱节虫,该组Rb-Sr法年龄值为8.5~7.4亿年。九里桥组化石很丰富,类型很多,其K-Ar法年龄值为7.38亿年左右,都大于7亿年。
陈孟莪等1982年—1991年在辽南复县震旦系下部长岭子组中发现了属于须腕动物的化石。主要分子有皱节虫(未定种)、寒武皱节虫、叶文金古线虫、辽南古线虫等[14],长岭子组的同位素年龄值为7.23亿年,与淮南九里桥组可以对比,郑文武将这些后生动物及与其共生的其他门类组合称之为淮南生物群[13](见图版Ⅴ及图版Ⅵ之4、5)。
淮南生物群如上所述是较原始的动物群落,因此它更接近于后生动物出现时的类型。一些人认为这些前伊迪卡拉动物群也是伊迪卡拉动物群的先驱分子,在自此以后1亿年左右的时间内形成的地层中,世界各地都大量的发现种类繁多、机体演化较高级的伊迪卡拉动物群和白海动物群,在中国相当的动物群为庙河生物群、西陵峡动物群和高家山动物群。
六、7亿~6亿年后生生物适应辐射爆发式增长事件
在7亿年左右时间内世界曾有三次重大冰期活动,其中第二冰期活动在全世界广泛分布(中国为南沱冰期),气候巨变,促进了生物适应辐射爆发式增长。因此在世界性大冰期之后各地广泛地突然出现大量繁多的后生动物,这在生物演化史上无疑是一次重大的生物事件,它生动地显示了后生动物演化上的重要转折和急剧变化,正在孕育着新的突破与飞跃。它也预示着地质历史的隐生宙阶段即将结束,显生宙的历史即将开始。
世界上最早发现这些多样的后生动物是1947年—1949年南澳大利亚弗林德斯山脉及伊迪卡拉地区,化石含量丰富,分属于腔肠动物门,水母类、水螅类、海鳃类、环节动物的多毛类、原始甲壳类以及分类位置未定的门类等,总称伊迪卡拉动物群(Ediacara Fauna),目前已采到1 500多件标本,成为世界最丰富、最有代表性的典型地区之一。自此以后,在世界各地已有多处出现相似的生物群落,我国在湖北三峡、秭归、陕西宁强、云南昆明等也都发现了这一类的化石。种类最多的是湖北秭归庙河生物群,其次还有湖北三峡的西陵峡生物群和陕西宁强的高家山生物群。
庙河生物群(丁莲芳等)。晚震旦世陡山沱期,700~650百万年,它是末元古宙世界性冰期以后的第一个生物群[15],它包括有腔肠动物的类水母、海绵动物、蠕虫动物及遗迹化石,还有各种演化较高级的宏体藻类,如褐藻、绿藻、红藻、硅藻及微体藻类、刺球类,它们有细胞结构和组织分化。庙河生物群是由生物类型比较单调和比较原始的状况转向多门类、较高级后生生物的产物,是它们对环境适应辐射的典范,此时物种分异度猛增,出现了140属322个种,就生物门类繁多和含量丰富来说,完全可以和伊迪卡拉生物群相媲美(见图4),这是一种由长期缓慢增长到爆发式增长的突变,是地质演化史上一次重大海洋生物大爆发事件。
西陵峡生物群(邢裕盛等)[16]、高家山生物群(张录易等)[17](见图版Ⅵ)。晚震旦世灯影期,650~600百万年,它是庙河生物群的延伸,物种由爆发式的发生而转入稳定水平上的发展,化石有腔肠动物的海腮类Charnia、海绵骨针、蠕虫类及大量的遗迹化石、褐藻等。西陵峡生物群和高家山生物群与庙河生物群前后相接,其持续时间相当于南澳Ediacara动物群生存的时间。

图4 庙河生物群化石类型图(据丁莲芳,1996)
从上述生物演化事件可以清楚地观察到生物界在演化的进程中,生物愈简单、低级,则演化愈慢;而愈趋于复杂、高级时则演化愈快。演化速度是以加速度的方式在前进着,也就是说它们各个发展阶段所占据的时间间隔是随着演化进程的推移而越来越短。它们不断地向着适合自己生存的环境扩张和迁移,按着自然演化的进程发生和发展。从末元古宙诸多生物群的特征来看,这些生物群既有联系,又有显著差别。尤其是在末元古宙全球最后一次冰期以后,海平面广泛上升,生物占领了更广阔的生态领域。同时较温暖的气候也提供了一个生物生存发展的有利条件。于是使得以宏体藻类、腔肠动物、蠕虫类、海绵类为代表的庙河生物群及其延伸的生物群应运而生,反映了这时期的生物已向更高级的阶段发展。也就是向生物门类更多、生态更为多样和机体分化程度更高的阶段发展。这不仅对末元古宙全球生物地层对比和研究有重要的现实意义,而且对末元古宙生物演化历史的研究也具有重要的理论意义。
附图:
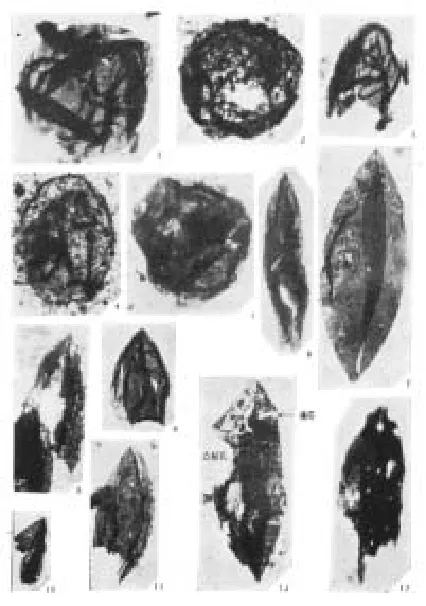
图版I

图版Ⅱ

图版Ⅲ
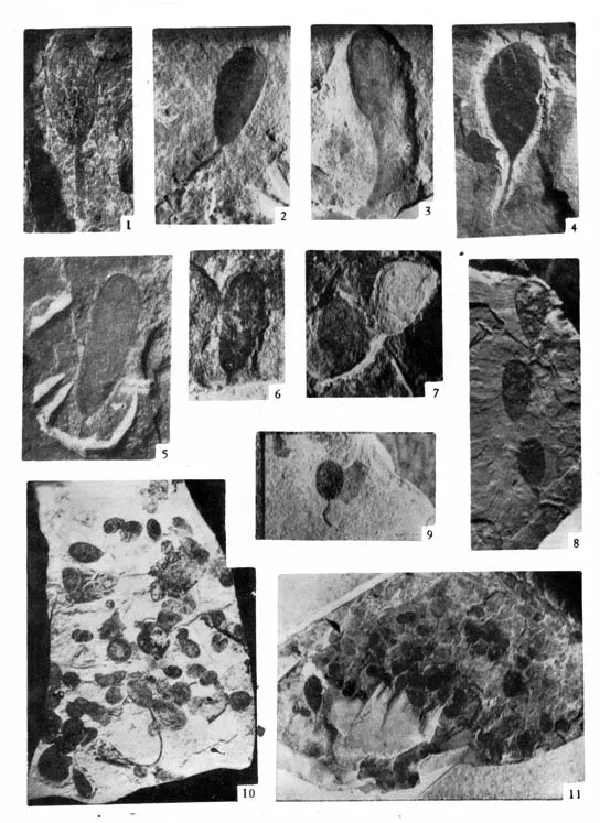
图版Ⅳ

图版Ⅴ

图版Ⅵ
图版说明:所有照片未加任何润饰,除特别注明为岩石磨制薄片和放大倍数外,全部为浸解法所获。化石照像放大倍数均为400倍,化石标本保存在天津地质矿产研究所。
参考文献:
[1] 罗其玲, 张月琳, 孙淑芬. 燕山地区长城系底部的真核生物化石[J]. 地质学报, 1985(1): 12-16.
[2] 陈晋镳, 等. 蓟县震旦亚界的研究[M]//. 中国震旦亚界.天津:天津科学技术出版社, 1980: 56-94.
[3] 张昀. 生物进化[M]. 北京: 北京大学出版社, 1998: 74-78.
[4] 杜汝霖, 胡华斌, 李培菊, 等. 冀西北长城系宣龙式铁矿生物成矿作用[M]. 北京: 科学出版社, 1999: 60-67.
[5] 阎玉忠. 蓟县串岭沟组裂梭藻[C]//中国地质科学院天津地质矿产所.中国地质科学院天津地质矿产研究所所刊, 1983(6): 1-6.
[6] WALCOTT C D. Precambrian Fossiliferous Formation[J]. Bulletion of the Geological Society of America, 1899,10: 235-242.
[7] 杜汝霖, 田立富, 李汉棒. 蓟县长城系高于庄组宏观生物化石的发现[J]. 地质学报, 1986(2): 115-200.
[8] 中国科学院贵阳地球化学研究所. 从燕山地区震旦系地层同位素年龄论中国震旦地质年表[J]. 中国科学, 1977(2):151-161.
[9] WALTER M R, OEHLER J H, OEHLER D Z. Megascopic Algae 1300 Million Years Old from the Belt Supergroup Montana: A Reinter Pretation of Walcott’s Helminthoidichnites[J]. Journal of Paleontology, 1976, 50 (5): 872-881.
[10] 杜汝霖, 田立富. 燕山青白系宏观藻类龙凤山藻属的发现和初步研究[J]. 地质学报, 1985(3): 183-190.
[11] 杜汝霖, 田立富. 我国晚前寒武纪“龙凤山生物群”[J]. 河北地质学院学报, 1986(3-4): 299-304.
[12] Hofmann H J. The Mid-Proterozoic Little Dal Macrobiota, Mackenzie Mountains, North-West Canada [J]. Palaeontology, 1985, 28(2): 331-354.
[13] 郑文武. 皖北震旦系中Chuaria等化石的发现及其地质意义[C]//中国地质科学院天津地质矿产研究所.中国地质科学院天津地质矿产研究所刊.北京:地质出版社.1980:
[14] 陈孟莪. 辽南晚前寒武纪地层中的宏体化石之地质意义的探讨[J]. 地质科学, 1991(2):120-128.
[15] 丁莲芳, 李勇, 胡夏嵩,等. 震旦纪庙河生物群[M]. 北京:地质出版社, 1996: 1-145.
[16] 邢裕盛, 段承华, 梁玉左, 等. 中国晚前寒武纪古生物[M]. 北京:地质出版社, 1985: 182-192.
[17] 张录易. 陕西宁强晚震旦世晚期高家山生物群的发现和初步研究[C]//中国地质科学院西安地质矿产研究所.中国地质科学院西安地质矿产研究所文集(13).西安:陕西科技出版社, 1986: 86-87.
(责任编辑:杜 敏)
Main Biological Evolution Events of Proterozoic Eon in China
The Proterozoic Eon is the second phase of the geological history of the Earth, which occupies a very important position in the geological history and the biological evolution history, and is a major turning point in the early biological evolution. Biological evolution is the most sensitive indicators of geological evolution, geological mutations will cause corresponding changes in the environment, species and ecological each time. Proterozoic Eon has experienced a series events such as the emergence of the eukaryotes, the emergence of the macro organisms, the emergence of the multi-cell tissue differentiation and the benthic organisms, and the emergence of metaphysical animals as well as their biological radiation burst, so that prompt the low-level, simple creatures in Cryptozoic Eons to develop and transit to a more advanced and complex new era in the Phanerozoic Eons.
Proterozoic Eon; Biological evolution; Sangshuania; Longfengshania
Q913.1
A
1007-6875(2017)01-0001-12
��日期:2017-01-11
10.13937/j.cnki.hbdzdxxb.2017.01.001
杜汝霖(1929—),男,辽宁沈阳人,河北地质大学(原河北地质学院)教授,长期从事前寒武纪地层和古生物方面的科学研究,开展了大量系统的开创性的工作,2001年荣获第七次李四光地质科学奖。
