水稻化感品种和非化感品种对稗草伴生的响应差异
2017-06-05孙备苏本卿郭伟王国骄周斌
孙备,苏本卿,郭伟,王国骄,周斌
1. 沈阳农业大学农学院,辽宁 沈阳 110866;2. 辽宁省气象科学研究所,辽宁 沈阳 110166
水稻化感品种和非化感品种对稗草伴生的响应差异
孙备1,苏本卿1,郭伟1,王国骄1,周斌2
1. 沈阳农业大学农学院,辽宁 沈阳 110866;2. 辽宁省气象科学研究所,辽宁 沈阳 110166
利用作物自身的化感作用控制农田杂草是生态安全条件下农田杂草综合防治的有效途径,但在水稻Oryza sativa和稗草Echinochloa crusgalli的种间关系研究中化感作用对资源竞争的影响常被忽略。通过研究田间稗草伴生条件下,水稻化感品种和非化感品种生物量及形态特征的变化,结合水稻和稗草植株养分含量及根区土壤养分含量,分析水稻化感品种和非化感品种对稗草竞争的响应差异。稗草混种后,水稻化感品种根系生物量提高、叶面积增大;非化感品种生物量、株高、叶面积显著下降。与水稻化感品种混种,稗草株高、生物量、叶面积与单种对照差异不显著;而与非化感水稻混种,稗草株高、生物量、叶面积显著升高。水稻化感品种和稗草混种,水稻植株磷含量和根区土壤铵态氮含量增加,稗草根区土壤养分含量降低;而水稻非化感品种和稗草混种,水稻根区土壤铵态氮含量和植株氮含量下降,稗草茎叶氮含量增加,根区土壤养分含量与单种差异不显著。水稻化感品种对稗草的相对总产量和养分竞争比例显著高于非化感品种。可见,水稻化感品种和非化感品种对伴生稗草的响应和影响不同,水稻和稗草伴生系统中化感品种较非化感品种对稗草更具有竞争优势,二者形态特征及其对稗草根区土壤养分影响的差异,影响了其自身对伴生稗草的竞争能力。
化感作用;形态特征;养分含量;水稻;稗草
水稻(Oryza sativa)是世界主要粮食作物之一,稗草(Echinochloa crusgalli)一直是稻田最严重的恶性杂草。稗草光能利用效率高、吸水吸肥能力强、幼苗生长快、植株高、叶面积大,在与水稻的竞争中占有优势,稗草共生是导致水稻产量下降的重要因素。品种不同,水稻产量下降幅度也不同,其与稗草的竞争能力也存在差异,分蘖数多、高秆、叶面积较大、晚熟的水稻品种在与稗草的竞争中具有相对优势(Smith,1974;Van Devender et al.,1997;Gealy et al.,2005)。此外,少数具有化感特性的水稻品种能够通过分泌化感物质抑制稗草的生长(Willey et al.,1980;Chen et al.,2008)。利用作物自身的化感作用控制农田杂草是生态安全条件下农田杂草综合防治的有效途径。
竞争和化感是两个不同却又相互关联的植物种间作用机制,但关于水稻化感特性对水稻与稗草竞争的影响常被忽略。水稻化感品种对稗草的化感抑制作用至少涉及两个密不可分的机制,即:水稻首先通过与伴生杂草形成化学识别及信息传递,然后合成、释放相应的化感物质抑制稗草(Kong et al.,2006;Kong,2008)。无论是稗草释放的信号物质还是水稻化感品种释放的化感物质都能够影响土壤微生物和土壤酶的活动(Gu et al.,2008;Sun et al.,2014),而土壤微生物群落结构和土壤酶活性与土壤养分可利用性密切相关。那么,在水稻化感品种与稗草的相互作用中,化感品种能否通过表型可塑性响应稗草的竞争?水稻化感品种能否通过养分竞争影响稗草生长?相关研究的开展将有助于了解水稻化感品种与稗草的种间相互作用,有助于深入分析水稻化感品种的抑草机制。本文通过田间实验,研究水稻化感品种和非化感品种对伴生稗草的响应差异。主要拟解决以下两个问题:(1)稗草伴生条件下,水稻化感品种和非化感品种表型可塑性响应的差异;(2)水稻化感品种和非化感品种对稗草养分竞争能力的差异。
1 材料方法
1.1 实验材料
化感品种选择PI32177,来源于美国USDAARS种质资源库。非化感品种选择辽粳9,由辽宁省水稻研究所提供。稗草种子收集于中国科学院沈阳生态实验站,自然风干后在尼龙袋中保存。
实验地位于中国科学院沈阳生态实验站(41°31′N,123°24′E),土壤类型为淋溶土(斑纹简育湿润淋溶土),土壤基本理化性质为pH 6.54;有机质1.74%;全氮1.6 g·kg-1;速效氮178.14 mg·kg-1;全磷0.41 g·kg-1;速效磷70.04 mg·kg-1;全钾1.20 g·kg-1;速效钾53.80 mg·kg-1。
1.2 实验设计
实验按水稻单种、水稻与稗草按1∶1间隔混种、稗草单种3种方式种植,2个水稻品种,共5个处理,采用随机区组实验设计,每个处理重复4次。小区设在4个29 m×1.2 m的苗床上,每个小区种植17行,每行12穴,行穴距为34 cm×12 cm,小区间距0.5 m。水稻和稗草采用直播的种植方式,每穴播种水稻或稗草7~8粒,在水稻2叶期进行间苗,每穴保留5株苗。总施氮量折合N 180 kg·hm-1、总施磷量折合P2O598 kg·hm-1、总施钾量折合K2O 120 kg·hm-1施肥。基肥施磷酸二铵200 kg·hm-1,分别在分蘖期和抽穗期追肥,第一次施硫酸铵500 kg·hm-1,第二次施硫酸铵300 kg·hm-1、硫酸钾300 kg·hm-1。
1.3 指标测定
1.3.1 植株生长发育情况
于水稻和稗草成熟期,在田间每个小区随机选取5穴植株,测定株高、叶面积和生物量。田间测定自然株高;采用干重法测定叶面积;将根、茎叶、籽粒分开,置于105 ℃烘箱中杀青30 min,然后在80 ℃下烘干至恒重,称量生物量。
1.3.2 植物养分含量
将每个小区所取的植物样品烘干称重后,混匀过筛。采用H2SO4-H2O2消煮法溶解植株中的氮、磷、钾。全氮采用半微量凯氏定氮仪测定;全磷采用钼锑抗比色法测定;全钾采用火焰分光光度计测定(鲁如坤,2000)308-315。
1.3.3 土壤养分含量测定
于水稻和稗草成熟期,在每个小区随机选取5穴水稻,取以根基为圆心0~5 cm范围内0~15 cm土层土壤,混匀,自然风干,过2 mm筛子,备用。铵态氮和硝态氮含量采用KCl提取-比色法测定;有效磷含量采用NaHCO3提取-钼锑抗比色法测定;有效钾含量用NH4Ac提取-火焰光度法测定(鲁如坤,2000)146-195。
1.4 竞争关系分析
1.4.1 相对总产量
采用相对总产量(Relative yield total,RYT)衡量物种的竞争能力,计算公式如下(Harper,1997):

式中,Ymi和Ymj分别为混种小区水稻和稗草的单穴生物量,Yi和Yj分别为单种小区水稻和稗草的单穴生物量。RYT>1表明两物种对资源有不同的需求,表现为避免竞争或共生关系;RYT=1表明两物种对相同的资源有共同的需求;RYT<1表明两物种相互竞争。
1.4.2 相对竞争能力
相对竞争能力(Relative competitive ability Ams)表示一种植物对另一种植物资源竞争能力的大小。计算公式如下(Willey et al.,1980):

式中,Ymi和Ymj分别为混种时水稻和稗草的生物量,Yi和Yj分别为单种时水稻和稗草的生物量。Ams>0表明水稻的竞争能力强于稗草;Ams=0表明水稻的竞争能力与稗草持平;Ams<0表明水稻的竞争能力弱于稗草。
1.4.3 营养竞争比例
采用水稻对稗草的养分竞争比例(Competition ratio of rice to barnyardgrass CRrb)来衡量水稻的养分竞争能力,计算公式如下(Morris et al.,1993):
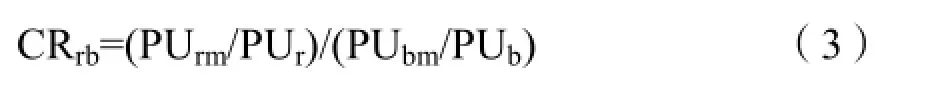
式中,PUrm和PUbm分别表示混种处理水稻和稗草养分吸收量,PUr和PUb分别表示单种处理水稻和稗草养分吸收量。CRrb>1表明水稻对养分的竞争能力强于稗草;CRrb=1表明水稻对养分的竞争能力与稗草持平;CRrb<1表明水稻对养分的竞争能力弱于稗草。
1.5 数据分析
同一水稻品种单种处理和稗草混种处理生物量、植株养分含量和土壤养分含量采用student’s t test比较。不同水稻品种混种对稗草的生物量、植株养分含量和土壤养分含量的影响差异采用单因素随机区组方差分析(one-way ANOVA)分析,不同处理间的差异采用Tukey’s检验比较。数据统计分析采用SPSS 16.0,图表绘制采用Excel 2007。
2 结果分析
2.1 稗草混种对水稻生长发育的影响
2个水稻品种生物量对稗草混种的响应不同(图1)。混种处理化感品种PI312777根生物量显著高于单种处理(t=4.60,P=0.019),但总生物量和茎叶生物量与单种处理差异不显著。混种处理非化感品种辽粳9根生物量和茎叶生物量分别较单种处理低62.4%和34.4%,二者之间差异达到显著水平(根:t=10.73,P=0.002;茎叶:t=3.35,P=0.015)。稗草混种导致水稻籽粒生物量降低(图2)。稗草混种处理,水稻品种PI312777和辽粳9籽粒生物量分别较单种下降23.8%和40.8%,其中,辽粳9与单种处理之间的差异达到显著水平(t=4.45,P=0.004)。

图1 稗草混种对水稻生物量的影响Fig. 1 Effects of barnyardgrass on biomass of mixed-cultured rice
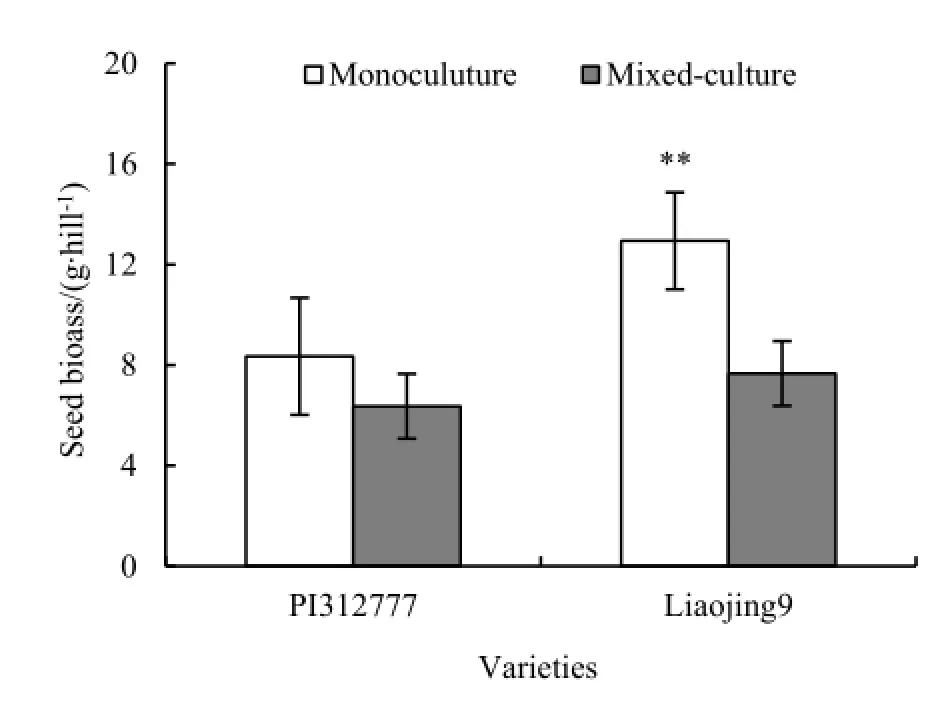
图2 稗草混种对水稻籽粒生物量的影响Fig. 2 Effects of barnyardgrass on seed biomass of mixed-cultured rice
稗草混种对化感品种PI312777株高影响不大,单种处理和混种处理差异不显著。在稗草混种条件下,非化感品种辽粳9的株高下降,与单种处理呈显著差异(t=2.71,P=0.035)(图3)。稗草混种对化感品种和非化感品种叶面积的影响也不同(图4)。PI312777单种处理叶面积显著低于稗草混种处理(t=2.66,P=0.037),而辽粳9单种处理叶面积显著高于稗草混种处理(t=3.72,P=0.010)。

图3 稗草混种对水稻株高的影响Fig. 3 Effects of barnyardgrass on height of mixed-cultured rice
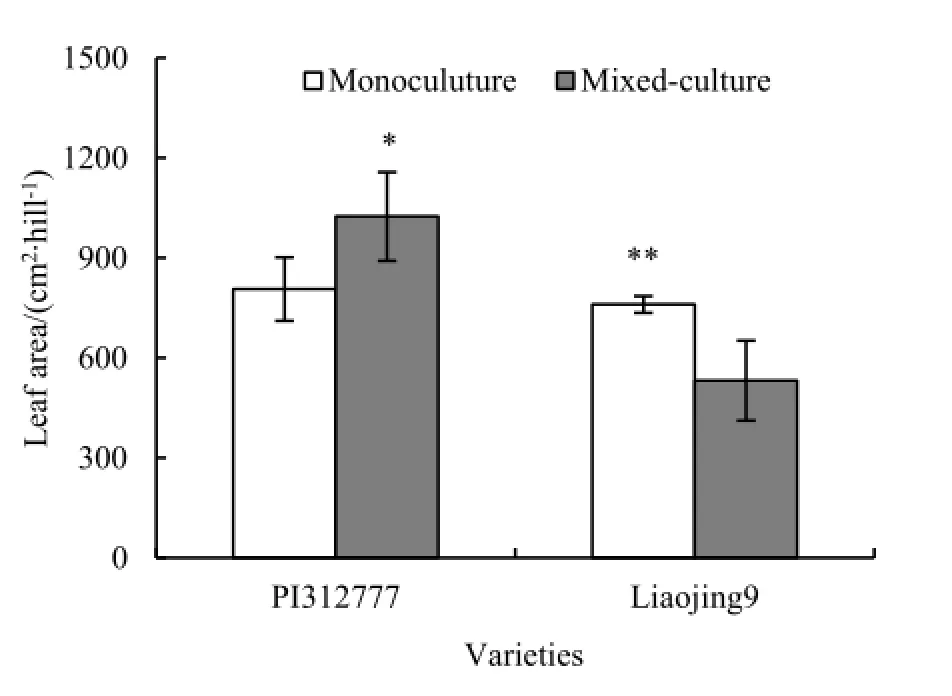
图4 稗草混种对水稻叶面积的影响Fig. 4 Effects of barnyardgrass on leaf area of mixed-cultured rice
2.2 水稻对混种稗草生长的影响
水稻和稗草混种处理稗草生物量高于单种处理,但化感品种PI312777和非化感品种辽粳9对伴生稗草生物量的影响不同(图5根生物量:F=17.17,P=0.003;茎叶生物量:F=14.6,P=0.005;图6籽粒生物量:F=28.9,P<0.001)。辽粳9混种处理稗草各生物量指标显著高于稗草单种处理(根生物量:t=17.17,P=0.003;茎叶生物量:t=4.24,P=0.004;籽粒生物量:t=9.98,P<0.001),但PI312777混种处理稗草各生物量指标与单种处理差异未达到显著水平。
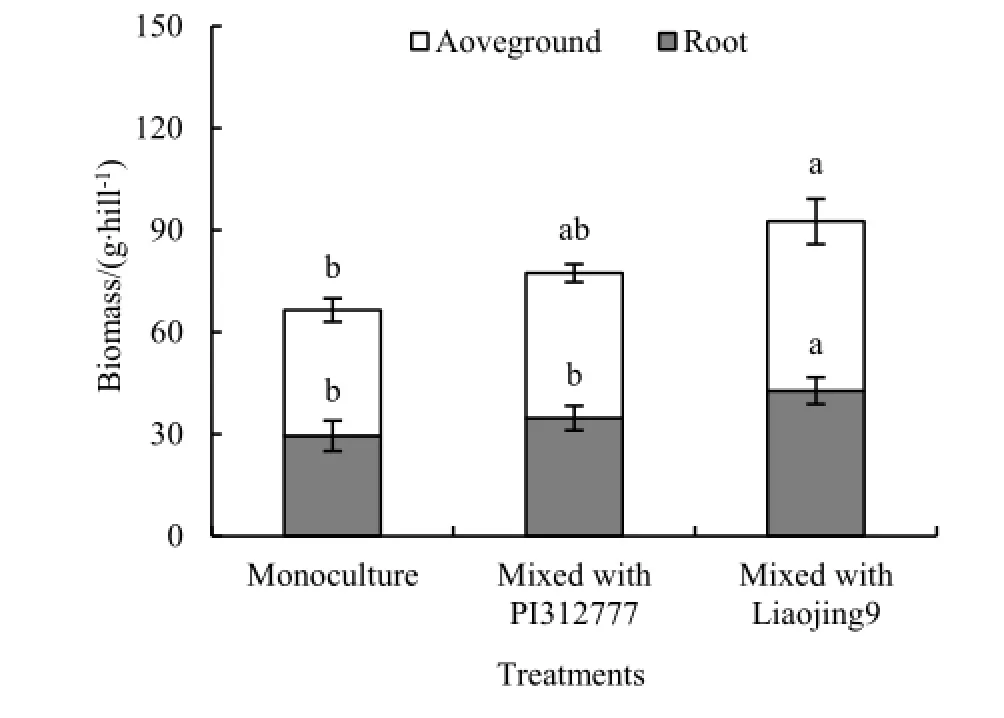
图5 水稻对混种稗草生物量的影响Fig. 5 Effects of rice on biomass of mixed-cultured barnyardgrass

图6 水稻对混种稗草籽粒生物量的影响Fig. 6 Effects of rice on seed biomass of mixed-cultured barnyardgrass
化感品种PI312777和非化感品种辽粳9对混种稗草株高均无显著影响(图7)。然而,PI312777和辽粳9对稗草叶面积的影响显著不同(图8 F=18.1,P=0.003),辽粳9混种处理稗草叶面积是单种的1.52倍(t=7.67,P=0.004),是其与PI31777混种的1.45倍(t=7.00,P=0.006)。
2.3 稗草混种对水稻土壤养分含量和植株养分含量的影响
混种处理化感品种PI312777和非化感品种辽粳9根区土壤硝态氮、有效磷和有效钾的含量与单种处理差异均不显著(表1)。但稗草混种处理2个水稻品种根区铵态氮含量的变化趋势不同,PI312777根区铵态氮含量显著高于单种处理(t=4.92,P=0.016),而辽粳9根区铵态氮含量显著低于单种处理(t=3.74,P=0.033)。
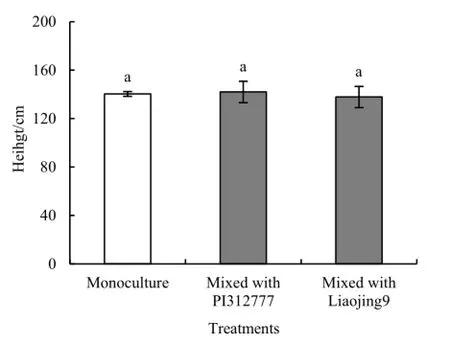
图7 水稻对混种稗草株高的影响Fig. 7 Effects of rice on height of mixed-cultured barnyardgrass barnyardgrass
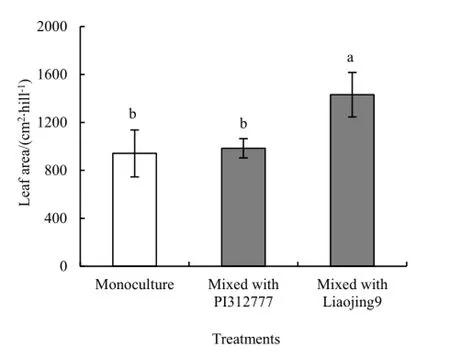
图8 水稻对混种稗草叶面积的影响Fig. 8 Effects of rice on leaf area of mixed-cultured
稗草混种对化感品种PI312777茎叶、根系和籽粒氮含量和钾含量影响不显著(表2);但显著提高了PI312777茎叶和籽粒的磷含量(茎叶:t=3.57,P=0.038;籽粒:t=3.21,P=0.048)。稗草伴生处理非化感品种辽粳9植株养分含量的变化与化感品种PI312777不同。稗草混种降低了辽粳9体内氮含量,其中茎叶和根系氮含量与单种处理呈显著差异(根系:t=4.07,P=0.027;茎叶:t=16.78,P<0.001)。稗草混种对辽粳9植株磷含量影响不明显,但显著提高了辽粳9茎叶的钾含量(t=17.97,P<0.001)。
2.4 水稻对混种稗草土壤养分含量和植株养分含量的影响
不同处理稗草根区土壤养分含量的变化趋势均表现为单种处理>辽粳9混种处理>PI312777混种处理(表3),其中铵态氮(F=6.78,P=0.029)、硝态氮(F=5.97,P=0.037)、速效钾含量(F=15.3,P=0.044)在各处理之间差异达到显著水平。然而,辽粳9混种处理稗草根区土壤养分与单种处理差异不显著,而PI312777导致混种稗草根区土壤养分含量显著降低(铵态氮:t=5.19,P=0.024;硝态氮:t=4.87,P=0.032;速效钾含量:t=7.68,P=0.040)。
化感品种PI312777和非化感品种辽粳9对混种稗草植株氮含量的影响不同(表4)。PI312777混种处理中,稗草茎叶氮和根系氮含量与单种处理的差异均未达到显著水平;辽粳9混种处理中,稗草茎叶氮含量较单种处理提高89.06%,显著高于单种处理(t=9.65,P=0.001)及其与PI312777混种处理(t=11.03,P<0.001)。与水稻混种后,稗草根系磷含量、茎叶和根系钾含量呈下降趋势,其中混种处理稗草茎叶钾含量与单种处理的差异达到显著水平(PI312777混种:t=9.99,P=0.001;辽粳9混种:t=6.46,P=0.009)。

表1 稗草混种对水稻根区土壤养分含量的影响Table 1 Effects of barnyardgrass on nutrient content in root zone soil of mixed-cultured rice mg·kg-1
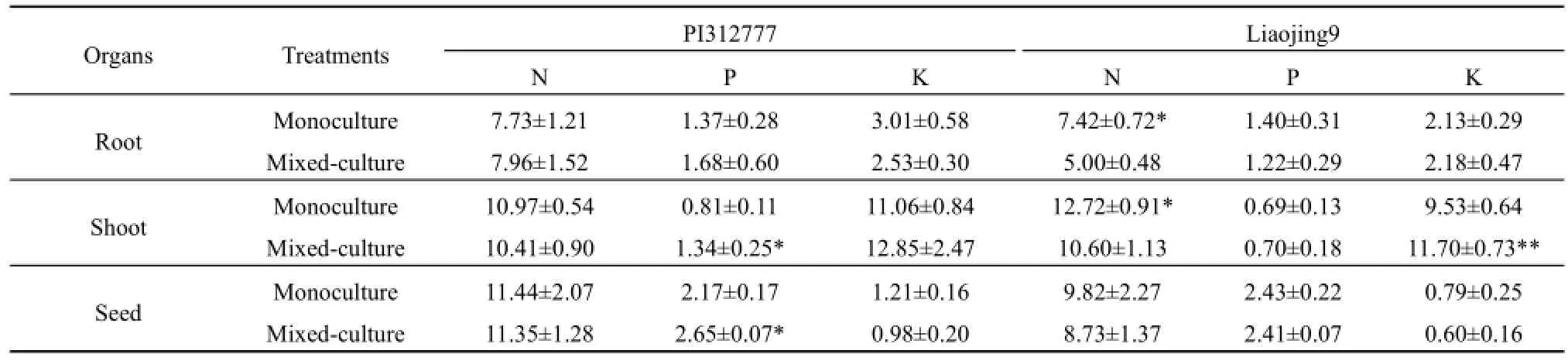
表2 稗草混种对水稻植株养分含量的影响Table 2 Effects of barnyardgrass on nutrient content of mixed-cultured rice g·kg-1

表3 水稻对混种稗草根区土壤养分含量的影响Table 3 Effects of rice on nutrient content in root zone soil of mixed-cultured barnyardgrass mg·kg-1
2.5 水稻和稗草竞争关系分析
2.5.1 相对竞争强度
水稻和稗草对资源需求相似,化感品种PI312777和非化感品种辽粳9与稗草的相对总产量均接近1,且二者之间差异不显著。然而,PI312777和辽粳9对稗草的相对竞争能力不同(表5)。PI312777对稗草的根系相对竞争能力高于0,地上部分相对竞争能力低于0。辽粳9根系和地上部分相对竞争能力均低于0,且PI312777对稗草的相对竞争能力高于辽粳9(根系:t=9.81,P=0.010;地上:t=4.55,P=0.020)。
2.5.2 养分竞争比例
化感品种PI312777和非化感品种辽粳9对稗草的养分竞争比例不同(表6)。PI312777根系和地上部分对稗草的养分竞争比例均大于1或接近1,但辽粳9对稗草的养分竞争比例均小于1,对二者养分竞争比例差异的t检验表明,PI312777根系和地上部分氮(根系:t=5.59,P=0.011;地上:t=6.28,P=0.008)、磷(根系:t=9.05,P=0.003;地上:t=3.63,P=0.036)、钾(根系:t=3.91,P=0.030;地上:t=7.93,P=0.004)对稗草的养分竞争比例均显著或极显著高于辽粳9。
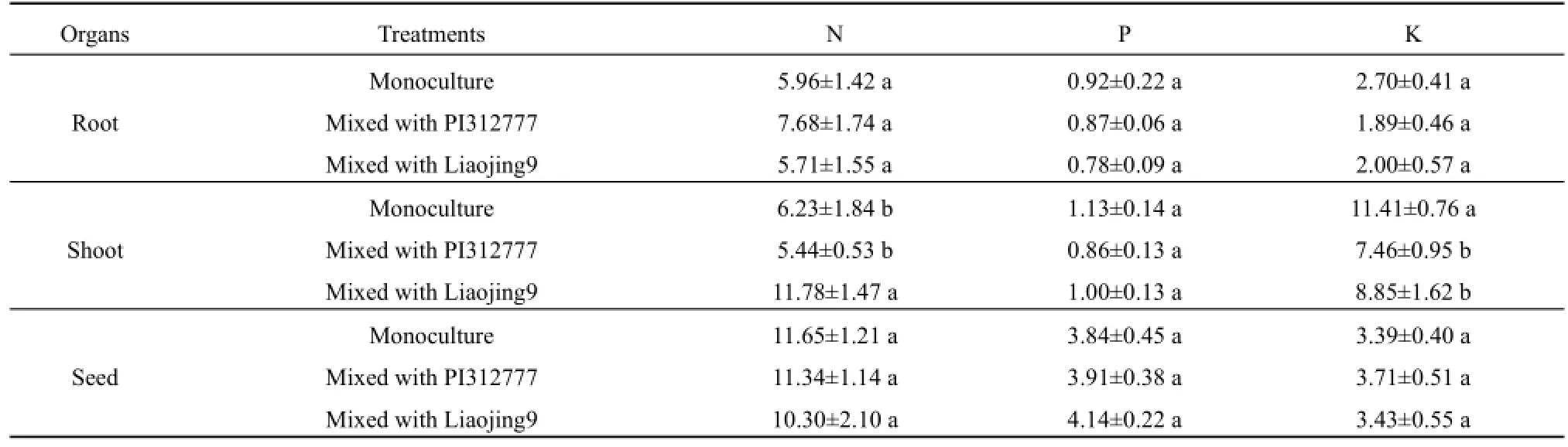
表4 水稻对混种稗草植株养分含量的影响Table 4 Effects of rice on nutrient content of mixed-cultured barnyardgrass g·kg-1

表5 水稻化感品种和非化感品种对稗草的相对总产量和相对竞争能力Table 5 Relative yield total and relative competitive ablitity of allelopathic rice and non-allelopathic rice to barnyardgrass

表6 水稻化感品种和非化感品种对稗草的养分竞争比率Table 6 Nutrition competition ratios of allelopathic rice and non-allelopathic rice to barnyardgrass
3 讨论
在水稻和稗草混种条件下,稗草生物量高于单种处理,但仅非化感品种混种处理与单种处理差异达到显著水平,表明与非化感水稻伴生有利于稗草的生长。稗草伴生对化感品种和非化感品种的影响强度也不同。稗草混种条件下,化感品种植株生物量特别是根生物量显著增加,而非化感品种根生物量、地上生物量和籽粒生物量均显著降低,表明稗草对化感水稻生长无显著抑制作用,但显著抑制非化感品种的生长。结合相对总产量和相对竞争强度分析可以看出,在水稻和稗草的伴生体系中,化感品种较非化感品种对稗草竞争能力更强。
地上部分竞争主要是对光的竞争。植物的光竞争能力与其株高、叶面积、光合特性等生物学特征密切相关(李博,2001)。在水稻和稗草伴生系统中,稗草为C4植物,水稻为C3植物,稗草株高较高、叶面积较大、光合利用能力强,较水稻具有光竞争优势,伴生水稻将有利于减弱稗草种内对光的竞争,促进稗草的生长发育(De Vida et al.,2006;Gealy et al.,2010)。然而,水稻化感品种和非化感品种对稗草竞争的响应不同。水稻和稗草混种条件下,化感品种叶面积增加将有助于其在与稗草的竞争中获得更多的光能,减少稗草株高和叶面积对光的竞争优势对自身生长发育的不利影响,表明水稻化感品种能够通过表型重塑更好地适应稗草伴生,提高自身对稗草的竞争能力。非化感品种与稗草混种,株高和叶面积均显著降低,稗草叶面积显著增加,表明伴生稗草能够抑制非化感品种的生长,从而提高光竞争优势,促进其自身的生长发育。
根系的生长状况影响了植物的养分吸收能力,根系生物量较高的水稻品种对稗草竞争能力较强(Perera et al.,1992)。化感品种与稗草混种时,根系生物量显著增加,根冠比增加,根系对稗草的相对竞争能力高于0,表明化感品种能够响应稗草竞争,促进自身根系的生长,以获得地下竞争优势。非化感品种与稗草混种条件下,非化感品种根系生物量显著降低,稗草根系生物量显著增加,表明伴生稗草地下竞争优势明显,抑制了非化感品种根系的生长。可见,水稻化感品种和非化感品种根系特征对稗草伴生的响应也不同,化感品种较非化感品种对稗草的地下竞争优势更为显著。在水稻和稗草伴生系统中,植物根区土壤养分状态与单种相比有明显不同。水稻和稗草混种处理,水稻化感品种根区养分含量增加,稗草根区养分含量降低,而非化感品种对伴生稗草根区养分含量没有影响,稗草伴生导致非化感品种根区土壤氮含量显著降低。尽管单种和混种条件下植物养分含量变化不明显,但对水稻对稗草的养分竞争比例(CRrb)的研究表明,水稻化感品种对稗草的养分竞争比例高于非化感品种。综合伴生条件下植物根区养分含量、植株养分含量和养分竞争比例(CRrb)可以看出,与水稻非化感品种相比,水稻化感品种能够影响水稻和稗草伴生系统植物根区土壤养分可利用性,提高自身对稗草的养分竞争能力。
环境因子也是影响水稻和稗草种间相互作用的重要因素。尽管有研究表明水稻化感品种对稗草生长发育有明显的抑制作用(Olofsdotter,2001;Gealy et al.,2003),但其对稗草生长发育的影响受到环境条件的制约,而施肥(Hill et al.,1994)和淹水(Perera et al.,1992)是影响水稻抑草特性的两个重要生态因子。尽管施肥有利于水稻产量的增加,其对稗草生长的促进作用却更强,施肥能够提高稗草对水稻的竞争力(Kazemeini et al.,2005)。施肥条件下,水稻化感品种对杂草的抑制能力下降,随着施肥量的增加化感品种对稗草的抑制率降低,甚至为负值(Kong et al.,2008)。本研究中肥料的施用可能是化感品种没有对稗草表现出显著抑制作用的重要因素。淹水可以显著抑制稗草的生长,形成有利于水稻的正反馈,随着淹水时间的延长和淹水深度的增加,水稻对稗草的抑制作用增强(Pinto et al.,2008)。本试验是在苗床上进行的,整个试验期苗床保持湿润,但几乎未出现长期淹水过程。淹水时间较短、淹水深度较浅可能也降低了本研究中水稻对稗草的竞争力。可见,水稻和稗草的种间相互作用受到环境条件的影响,而不同环境条件下水稻化感品种对稗草竞争抑制的影响仍有待进一步研究。
4 结论
稗草混种条件下,水稻化感品种根系生物量提高、叶面积增大,植株磷含量、根区土壤铵态氮含量增加;非化感品种生物量、株高、叶面积显著下降,根区土壤铵态氮含量和植株氮含量亦下降。与水稻化感品种混种,稗草株高、生物量、叶面积与单种对照差异不显著,根区土壤养分含量降低;与非化感水稻混种,稗草株高、生物量、叶面积显著升高,根区土壤养分含量与单种差异不显著。化感品种对稗草的相对竞争能力和养分竞争比例高于非化感品种。可见,水稻化感品种和非化感品种对稗草的伴生响应不同。稗草伴生条件下,水稻化感品种形态特征的响应及其对稗草根区土壤养分的影响,提高了其自身对伴生稗草的竞争能力。
致谢:感谢中国科学院沈阳生态实验站在实验过程中给予的帮助。
CHEN X H, HU F, KONG C H. 2008. Varietal improvement in rice allelopathy [J]. Allelopathy Journal, 2(2): 379-384.
DE VIDA F B P, LACA E, MACKILL D, et al. 2006. Relating rice traits to weed competitiveness and yield: a path analysis [J]. Weed Science, 54(6): 1122-1131.
GEALY D R, FISCHER A J. 2010.13C Discrimination: A stable isotope method to quantify root interations between C3Rice (Oryza sativa) and C4Barnyardgrass (Echinochloa crus-galli) in Flooded Fields [J]. Weed Science, 58: 359-368.
GEALY D R, ESTORNINOS L E, GBUR E E, et al. 2005. Interference interactions of two rice cultivars and their F3 crosss with barnyardgrass (Echinochloa crus-galli) in a replacement series study [J]. Weed Science, 53: 323-330.
GEALY D R, WAILES E J, ESTORNINOS L E, et al. 2003. Rice cultivar differences in suppression of barnyardgrass (Echinochloa crus-galli) and economics of reduced propanil rates [J]. Weed Science, 51(4): 601-609.
GU Y, WANG P, KONG C H. 2008. Effects of rice allelochemicals on the microbial community of flooded paddy soil [J]. Allelopathy Journal, 22(2): 299-309.
HARPER J. 1997. Population Biology of Plants [M]. London: Academic Press.
HILL J E, SMITH R J, BAYER D E. 1994. Rice weed control current technology and emerging issues in temperate rice [J]. Australian Journal of Experimental Agriculture, 34(7): 1021-1029.
KAZEMEINI S A, GHADIRI H. 2005. Interaction effect of barnyardgrass (Echinochloa crus-galli) densities and different levels of nitrogen fertilizer application on growth and yield of rice in Kushkak, Fars Province [J]. Iranian Journal of Plant Pathology, 41(1): 95-106.
KONG C H, HU F, WANG P, et al. 2008. Effect of allelopathic rice varieties combined with cultural management options on paddy field weeds [J]. Pest Management Science, 64(3): 276-282.
KONG C H, LI H B, HU F, et al. 2006. Allelochemicals released by rice roots and residues in soil [J]. Plant and Soil, 288(1): 47-56.
KONG C H. 2008. Rice allelopathy [J]. Allelopathy Journal, 22(2): 261-273.
MORRIS R A, GARRITY D P. 1993. Resource capture and utilization in intercropping: non-nitrogen nutrients [J]. Field Crops Research, 34(3-4): 319-334.
OLOFSDOTTER M. 2001. Rice-A step toward use of allelopathy [J]. Agronomy Journal, 93(1): 3-8.
PERERA K K, AYRES P G, GUNASENA H P M. 1992. Root-growth and the relative importance of root and shoot competition in interactions between rice (Oryza sativa) and Echinochloa crus-galli [J]. Weed Research, 32(1): 67-76.
PINTO J J O, GALON L, DAL MAGRO T D, et al. 2008. Control of Echinochloa spp. as a function of management methods in flooded rice [J]. Planta Daninha, 26: 767-777.
SMITH R J. 1974. Competition of Barnyardgrass with rice cultivars [J]. Weed Science, 22(5): 423-426.
SUN B, WANG P, KONG C H. 2014. Plant-soil feedback in the interference of allelopathic rice with barnyardgrass [J]. Plant and soil, 377(1): 309-321.
VAN DEVENDER K W, COSTELLO T A, SMITH R J. 1997. Model of rice (Oryza sativa) yield reduction as a function of weed interference [J]. Weed Science, 45(2): 218-224.
WILLEY R W, RAO M R. 1980. A competitive ratio for quantifying competition between intercrops [J]. Experimental Agriculture, 16(2): 117-125.
李博. 2001. 植物竞争——作物与杂草相互作用的实验研究[M]. 北京:高等教育出版社: 1-14.
鲁如坤. 2000. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社.
Differences between Allelopathic and Non-allelopathic Rice in Responses to Competiting Barnyardgrass
SUN Bei1, SU Benqing1, GUO Wei1, WANG Guojiao1, ZHOU Bin2
1. Agronomy Department, Shenyang Agriculture University, Shenyang 110866, China; 2. Liaoning Institute of Meteorological Science, Shenyang 110166, China
It is an effective way to develop integrated and ecological security weed management to control weedy rice using crop allelopathic varieties. However, there are few studies on the effects of allelopathy on competition in rice-barnyardgrass coexistence. We studied the resource competition ability of allelopathic rice and non-allelopathic rice compared to barnyardgrass using biomass, morphological plasticity, nutrient contents of plant and root zone soil in monoculture and mixed-culture in the field to test whether there were differences between allelopathic and non-allelopathic rice in response to competing barnyardgrass. When mixed with barnyardgrass, there was the increase of root biomass and leaf area of allelopathic rice, and decrease of biomass and leaf area of non-allelopathic rice. Mixed-culture with allelopathic rice did not lead to significant differences in morphological characteristic and nutrient content of barnyardgrass. However, there were significant increase of height, biomass, and leaf area of barnyardgrass in mixed-culture with non-allelopathic rice. Soil ammonium nitrogen in root zone and nitrogen content of allelopathic rice increased in presence of barnyardgrass, while non-allelopathic rice had lower nitrogen content of plant in mixed-culture with barnyardgrass than in monoculture. Allelopathic rice had more relative competitive ability and nutrient competition ratio of rice to barnyardgrass compared to non-allelopathic rice. All these results indicate morphological plasticity and effects on soil nutrient content in root zone of allelopathic rice improve its ability to compete with neighboring barnyardgrass compared to non-allelopathic rice.
allelopathy; morphological plasticity; soil nutrient content; rice; barnyardgrass
10.16258/j.cnki.1674-5906.2017.03.002
S314; X173
A
1674-5906(2017)03-0371-08
孙备, 苏本卿, 郭伟, 王国骄, 周斌. 2017. 水稻化感品种和非化感品种对稗草伴生的响应差异[J]. 生态环境学报, 26(3): 371-378.
SUN Bei, SU Benqing, GUO Wei, WANG Guojiao, ZHOU Bin. 2017. Differences between allelopathic and non-allelopathic rice in responses to competiting barnyardgrass [J]. Ecology and Environmental Sciences, 26(3): 371-378.
国家自然科学基金项目(31600350);辽宁省科学技术计划博士启动基金项目(201501061);辽宁省教育厅科学技术研究计划项目(L2015486)
孙备(1980年生),女,讲师,博士,主要从事农业生态学研究。E-mail: ecology_syau@163.com
2016-12-28
