闽东秋茄(Kandelia candel)人工林与天然林异龄叶性状比较
2017-05-30潘云龙朱清波陈欣凡李彬欧阳玉莹吴承祯洪滔
潘云龙 朱清波 陈欣凡 李彬 欧阳玉莹 吴承祯 洪滔


摘 要 比较闽东不同生境秋茄[Kandelia candel (Linn.) Druce]人工林与天然林异龄叶性状间的相互关系,结果显示:(1)2种林分的秋茄林叶MLA、SLA和LDMC,当年生均大于往年生,分别为MLA(38.4 cm2>36.0 cm2)、SLA(78.9 m2/kg>67.5 m2/kg)、LDMC(258.7 mg/g>193.8 mg/g);(2)当年生和往年生叶MLA人工林均大于天然林,分别为当年生(40.7 cm2>36.1 cm2)、往年生(37.5 cm2>34.4 cm2)。当年生叶SLA人工林小于天然林,为(78.4 m2/kg<79.4 m2/kg),往年生叶SLA人工林大于天然林,为(71.0 m2/kg>64.0 m2/kg)。当年生与往年生叶LDMC人工林均大于天然林,分别为当年生(344.4 mg/g>173.0 mg/g)、往年生(225.6 mg/g>162.0 mg/g)。研究表明,异龄叶对光资源获取和储存物质以构建叶器官这2种投资策略的权衡,是植物在各自生境中适应性进化的结果。秋茄能对生境因子胁迫调整生活策略应对,具有较强的环境适应性,适宜作为海岸防护林树种人工栽植。
关键词 秋茄;异龄叶;叶性状
中图分类号 S718.3 文献标识码 A
Abstract In the paper, the experiments were conducted to compare the foliar traits of uneven-aged leaves of Kandelia candel plantations and natural forests in the eastern regions of Fujian Province. The results showed that: (1)The leaf MLA, SLA and LDMC of the two kinds of K. candel forests were all higher in current-year than those in former-year leaves, respectively(38.4 cm2>36.0 cm2), (78.9 m2/kg>67.5 m2/kg), (258.7 mg/g>193.8 mg/g).(2)The MLA in both current-and former-year leaves of the plantation were significantly higher than those of the natural forest, respectively(40.7 cm2>36.1 cm2), (37.5 cm2>34.4 cm2). The leaf SLA of the plantation was higher in current-year leaves of the natural forest(78.4 m2/kg<79.4 m2/kg), while former-year leaves showed the reverse trend (71.0 m2/kg>64.0 m2/kg). The leaf LDMC was greater in the plantation than the natural forest in current-year and former-year leaves, respectively(344.4 mg/g>173.0 mg/g), (225.6 mg/g>162.0 mg/g). The study showed it was the result of adaptive evolution for uneven-aged leaves to hold investment trade-off between light resources acquisition and material storage to build leaf organs in various habitats. The K. candel could adjust live strategies to cope with stressed factors and develop strong environment adaptability, and therefore K. candel was suitable to be planted as the coastal shelter forest trees.
Key words Kandelia candel; uneven-aged leaves; leaf traits
doi 10.3969/j.issn.1000-2561.2017.04.007
植物葉片是维持陆地生态系统机能的最基本要素,是大气-植物系统能量交换的基本单元[1],对环境变化具有重要的指示意义[2]。植物叶片适应环境变化所形成的生存对策[3-6]称为植物叶性状(leaf traits),叶性状作为植物性状的重要定量指标,能够反映植物因环境的变化而形成的内在生理及外在形态方面的适应对策及植物资源的利用能力,被认为是环境状况的敏感指示者[7],具有重要的生态学及植物生理学研究价值[8-10]。
随着对自然环境变化研究的深入,叶性状成为研究植物对环境的适应机理的热点[11]。前人研究认为,植物往年生叶片因生活史较长,结构成熟固化,叶片中很大一部分物质用于增加叶肉细胞密度或构建保卫结构[12-14],因此,面对环境因子变化时,可能响应不敏感[15];此外,随叶龄增加,叶片衰老过程中结构改变、叶片内部CO2导度下降、扩散受阻,叶内主要养分元素含量及叶片光合能力衰退,由此引起的资源再分配导致异龄叶对环境响应表现出较大的差异[16-18]。而当年生叶处于叶片生长初期,需投入更多资源于强化光合器官、获得最大的光资源竞争优势[19-20],以增大光截获能力去补偿自身建成消耗来应对环境变化[21],也有学者认为,展叶过程中叶片的光合作用强度与叶寿命成反比[22],叶龄低的个体,一般要比叶龄高的光合能力强,对于水分胁迫的调节能力较往年生强[23],因此,当环境因子发生扰动时,当年生叶对环境的响应可能较生长后期的叶片更为敏感[24]。由于当年生叶与往年生叶在获取资源能力及维持生态系统稳定性方面具有不同的生态功能,由此推断,海岸湿地植物异龄叶间的叶性状因生长策略、生活史不同可能产生差异。平均叶面积(mean leaf area,MLA)、比叶面积(specific leaf area,SLA)和叶干物质含量(leaf dry matter content,LDMC)是植物叶片的三个关键叶性状,综合反应植物利用资源的能力以及适应环境的生存对策。其中,MLA反映植物获取光资源的能力;SLA反映了叶片捕获光照资源的能力和形成干物质的结果;LDMC为叶片的干重与鲜重之比,表征了叶片在自然状态下的含水率状况。前人对MLA、SLA和LDMC等主要叶片特性之间的关系进行大量研究[25],项目组也曾研究叶片在不同环境下的适应策略[26]及海岸山地优势树种叶性状海拔资源位特征[27-28]、薛立等[29]在逆境情况下预测叶性状变化特征、王希群等[30]对叶面积指数进行研究、黄海侠等[31]分析常绿植物新老叶特征等,但研究海岸湿地树种异龄叶片性状差异的研究仍未见报道[32]。
研究地位处福建省罗源湾滨海湿地,地方为发展经济,近年在湾区兴建钢铁厂等重工业,这些外部因素促使环境因子明显改变,秋茄叶可能对这种环境变化产生响应,因此从不同林分类型秋茄异龄叶片受环境影响产生差异角度探寻海岸湿地植物叶片对环境变化的响应规律,对推动我国东南沿海红树林恢复、保护提供科学依据。
1 材料与方法
1.1 材料
研究区域位于福建省福州市罗源湾滨海区(26°23′~26°39′N,119°07′~119°54′E),全区东西长约25 km,南北宽约15 km,总面积约230 km2。罗源湾口窄腹大,海湾曲折,呈倒葫芦状,在东北角缺口通东海,属中亚热带海洋性季风气候,年平均气温19 ℃,年平均降水量1 652 mm,年平均日照时数1 691.1 h。研究区曾大量分布秋茄红树林,后被互花米草入侵替代,当地林业部门治理互花米草后,在清理迹地多次人工栽植多种红树林树种,唯密植(株距0.5 m)的秋茄幼苗成功存活。现复植秋茄林成林面积33 hm2,林相整齐,平均树高2 m,林龄3 a。天然秋茄林均高2.5 m,林龄5 a,株距0.9 m,分布均匀,长势旺盛。
1.2 方法
按多点混合法采取植物样品,在人工林与天然林各设5块10 m×10 m样地,每样地选择3株标准木进行叶片采集。采集叶片时,依据枝条上芽鳞痕形成的节环来区分不同年龄枝段,当年生枝段上已完全展开的叶片定义为当年生叶片,其他较老枝条上的叶片统称为往年生叶片。使用枝剪从每株个体的4个不同方向各采剪10片完好无损的叶片。采集叶片后打湿处理,放入自封袋密封并放置水桶内带回。在12 h内,从每个自封袋中取40片完好无损的叶片清洗擦干,稱其鲜重。用叶面积仪CI-203(美国产)测定样品叶面积(S),每叶重复3次。将叶片装入信封置于干燥箱中以80℃烘干至恒重,再称量其干重。
通过以下公式计算出每株叶片的MLA(cm2)、SLA(m2/kg)和LDMC(mg/g):
为避免从单一植物类型讨论植物叶片性状变异,从天然林与人工林两个角度探讨阔叶树种异龄叶片的变异性。在相同条件下,对同一被测量进行连续多次测量,所得结果的一致性配对数据t检验来判断叶片性状在异龄叶片之间的差异。计算异龄叶性状变异系数,以体现叶性状变异程度。
1.3 数据与分析
采用Excel 2016软件对试验所得数据进行均值、标准差、变异系数的计算,运用SPSS 19.0中的单因素方差分析法(One-way ANOVA)和多重比较(least-significant difference,LSD)法对比同一林分不同叶龄间及不同林分相同叶龄间各性状的差异。
2 结果与分析
2.1 异龄叶性状间的差异性
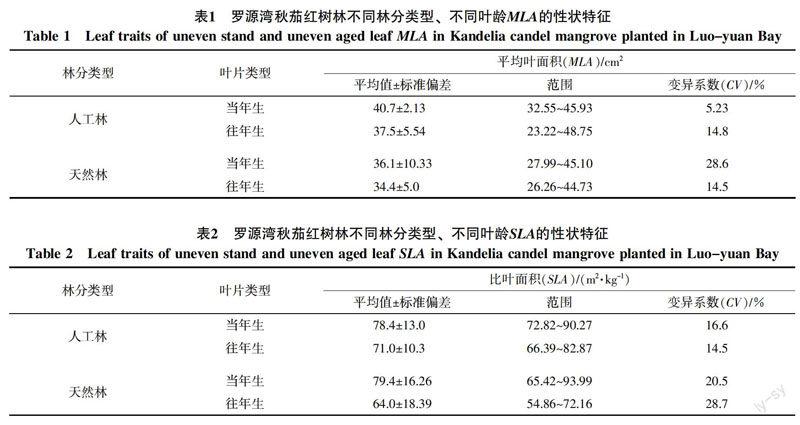
从异龄叶性状比较分析,罗源湾秋茄红树植物异龄叶MLA、SLA和LDMC表现出一定规律性(表1~3)。总体上,人工林与天然林叶面积MLA、SLA与LDMC当年生均大于往年生,分别为(38.4 cm2>36.0 cm2)、(78.9 m2/kg>67.5 m2/kg)、(258.7 mg/g>193.8 mg/g)。①对同种林分不同叶龄的秋茄林而言,人工林与天然林当年生MLA大于往年生,分别为(40.7 cm2>37.5 cm2)、(36.1 cm2>34.4 cm2)。SLA当年生也大于往年生,分别为(78.4 m2/kg>71.0 m2/kg)、(79.4 m2/kg>64.0 m2/kg);LDMC同样大于往年生,分别为(344.4 mg/g>225.6 mg/g)、(173.0 mg/g>162.0 mg/g)。②从不同林分相同叶龄的秋茄林而言,人工林当年生与往年生叶MLA均大于天然林当年生与往年生叶片,分别为(40.7 cm2>36.1 cm2)、(37.5 cm2>34.4 cm2)。人工林当年生叶SLA小于天然林当年生叶,为(78.4 m2/kg<79.4 m2/kg),往年生相反,为(71.0 m2/kg>64.0 m2/kg)。人工林当年生与往年生LDMC均大于天然林,分别为(344.4 mg/g>173.0 mg/g)、(225.6 mg/g>162.0 mg/g)。
2.2 异龄叶性状的变异性
变异系数是衡量植物变异程度的一个统计量,变异系数大说明性状可塑性较强。对秋茄林叶性状变异系数研究发现:①在同种林分不同叶龄的秋茄林变异系数上,人工林当年生MLA小于往年生,为(5.23%<14.77%),天然林MLA却相反,为(28.6%>14.5%);SLA变异系数表现出与MLA相反的规律,为人工林当年生变异系数大于往年生,天然林当年生变异系数小于往年生;其值为(16.6%>14.5%)、(20.5%<28.7%);人工林LDMC变异系数小于往年生,分别为(13.7%>9.7%)、天然林变异系数相等(18.7%<18.1%)。②在不同林分相同叶龄的秋茄林变异系数上,人工林当年生MLA变异系数明显小于天然林当年生,为(5.23%<28.6%)、人工林往年生变异系数大于天然林往年生,为(14.8%>14.5%);人工林当年生与往年生SLA变异系数明显小于天然林当年生与往年生,为(16.6%<20.5%)、(14.5%<28.7%)。人工林当年生与往年生LDMC变异系数小于天然林当年生与往年生,为(13.7%<18.7%)、(9.7%<18.1%)。
3 讨论
常绿植物叶片生活史较长,面临复杂多变的环境状况,为适应环境条件,致使叶面积在全球范围内变化达到6个数量级[33],因此,叶片性状(MLA、SLA和LDMC)反映了植物适应性进化过程及生活策略差异[34]。本研究中,秋茄MLA分布于23.22~48.75 cm2,人工林的MLA均值当年生与往年生分别为40.7、37.5 cm2,天然林的MLA均值当年生与往年生分别为36.1、34.4 cm2,均表现出当年生MLA大于往年生MLA的规律。前人认为,植物叶片面积受养分[35]、温度[36]等环境因素影响;但本研究结论表明,叶龄也明显影响叶面积大小。本研究中,秋茄叶MLA当年生大于往年生的规律与黄海侠等[31]研究天童山常绿阔叶植物MLA结论相反,这可能因秋茄为海岸湿地树种,其生境较陆生植物恶劣,林分密度远高于陆生植物(本研究区秋茄人工林株距为50 cm2、天然林株距为90 cm2),以抵御潮汐水流的冲击。这导致植物个体之间空间竞争激烈,植物必须尽可能扩大占据较优生态位的新叶叶面积,获取更多光照等资源以满足自身需求导致。光照条件是影响叶面积大小的重要因素,更大的叶面积被认为是应对低光环境的生态学策略[37]。在湾区内新建的钢铁厂等重工业排放,造成空气中反射太阳光的颗粒物大量增加,可能导致新叶在一定程度上面临光强较往年降低,为弥补自身需求,获取最大的光资源和碳生产,当年生叶片在生长初期就将叶片面积展开至最大[38-39]。秋茄SLA人工林与天然林当年生大于往年生,分别为(78.4 m2/kg>71.0 m2/kg)、(79.4 m2/kg>64.0 m2/kg),说明SLA大的叶片,其叶片干物质含量较小,同时较大的叶片面积有利于捕获更多的光能[40]。SLA还体现植物构建叶面积时所投资的单位生物量, 与叶片建成消耗负相关[41],秋茄往年叶较当年叶承受更长时间的环境胁迫,在防御保护构造上的额外投资较多[34],导致往年叶的建成消耗大于当年叶。
秋茄人工林与天然林当年叶与往年叶的各性状指标具有明显差别。天然林林龄较长,从其树高(2.5 m)、株距(0.9 m)可以看出,叶片空间结构及生长策略相对稳定。而人工林林龄只有2 a,为适应环境胁迫,林分密度较大(株距0.5 m),造成林分内个体竞争激烈,叶性状指标较天然林更高。也说明秋茄容易受生境因子胁迫,调整生活策略应对,有较强环境适应性,适宜作为海岸防护林树种人工栽植。
总之,人工林和天然林异龄叶性状指标均具有明显差别,这反映异龄叶对光资源获取能力、储存物质构建叶器官这两种投资策略的权衡,是植物在各自生境中适应性进化的结果。
参考文献
[1] 毛 伟, 李玉霖, 张铜会, 等. 不同尺度生态学中植物叶性状研究概述[J]. 中国沙漠, 2012, 32(1): 33-41.
[2] 张 林, 罗天祥, 邓坤枚, 等. 云南松比叶面积和叶干物质含量随冠层高度的垂直变化规律[J]. 北京林业大学学报, 2008, 30(1): 40-44.
[3] Ackerly D. Self-Shading, Carbon gain and leaf dynamics: a test of alternative optimality models[J]. Oecologia, 1999, 119(3): 300-310.
[4] Coley P D. Effects of plant-growth rate and leaf lifetime on the amount and type of anti-herbivore defense[J]. oecologia, 1988, 74(4): 531-536.
[5] Vendramini F, Díaz S, Gurvich D E, et al. Leaf traits as indicators of resource-use strategy in floras with succulent species[J]. New Phytologist, 2002, 154(1): 147-157.
[6] Westoby M. A leaf-height-seed(LHS0plant ecology strategy scheme[J]. Plant and soil, 1998, 199(20): 213-227.
[7] 許松葵, 薛 立. 6种阔叶树种幼林的叶性状特征[J].西北林学院学报, 2012, 27(6): 20-25.
[8] Chapin F S, Bloom A J, Field C B, et al, Plant-responses to multiple environmental-factors[J]. Bioscience, 1987, 37(1): 49-57.
[9] Wright I J, Groom P K, Lamont B B, et al. Leaf trait relationships in Australian plant species[J]. Functional Plant Biology, 2004, 31(5): 551-558.
[10] 张 林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展[J]. 植物生态学报, 2004, 28(6): 844-852.
[11] 李永华, 罗天祥, 卢 琦. 青海省沙珠玉治沙站17种主要植物叶性因子的比较[J]. 生态学报, 2005, 25(5): 994-999.
[12] Zhang L, Luo T. Advances in ecological studies on leaf lifespan and associated leaf traits[J]. Acta Phytoecologica Sinica, 2004, 28(6): 844-852.
[13] Westoby M. The worldwide leaf economics spectrum[J]. Nature, 2004, 428(6 985): 821-827.
[14] Brian Anacker, Nishanta Rajakaruna, David Ackerly, et al. Ecological strategies in California chaparral: interacting effects of soils, climate, and fire on specific leaf area[J]. Plant Ecology & Diversity, 2011, 4(2-3): 179-188.
[15] Oguchi R, Hikosaka K, Hirose T. Does the photosynthetic light-acclimation need change in leaf anatomy?[J]. Plant Cell & Environment, 2003, 26(4): 505-512.
[16] Field C. Allocating leaf nitrogen for the maximization of carbon gain: Leaf age as a control on the allocation program[J]. Oecologia, 1983, 56(2): 341-347.
[17] Bleecker A B. The evolutionary basis of leaf senescence: Method to the madness?[J]. Current Opinion in Plant Biology, 1998, 1(1): 73-78.
[18] Ono K, Nishi Y, Watanabe A, et al. Possible Mechanisms of Adaptive Leaf Senescence[J]. Plant Biology, 2001, 3(3): 234-243.
[19] Field C, Mooney H A. Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub[J]. Oecologia, 1983, 56(2): 348-355.
[20] 夏莹莹, 毛子军, 马立祥, 等. 水分条件对红松和西伯利亚红松针叶脯氨酸与叶绿素含量的影响[J]. 植物研究, 2008, 28(3): 330-335.
[21] Schaffer W M, Fox G A. Plant strategies and the dynamics and structure of plant communities[J]. The Quarterly Review of Biology, 1989, 64(2): 409-411.
[22] Warren C R. Why does photosynthesis decrease with needle age in Pinus pinaster?[J]. Trees, 2006, 20(2): 157-164.
[23] Sultan S E. Phenotypic plasticity for plant development, function and life history[J]. Trends in Plant Science, 2000, 5(12): 537-542.
[24] Westoby M. The worldwide leaf economics spectrum[J]. Nature, 2004, 428(6985): 821-827.
[25] ülo Niinemets. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs[J]. Ecology, 2001, 82(2): 453-469.
[26] 王英姿, 洪 伟, 吴承祯, 等. 灵石山米槠林优势种群不同叶龄叶属性的研究[J]. 福建林学院学报, 2009, 29(3): 203-209.
[27] 洪 滔, 吴承祯, 陈 灿, 等. 福建海岸山地5种优势树种叶形态特征对海拔资源位的响应[J]. 福建林学院学报, 2013, 33(1): 7-11.
[28] 洪 滔, 吴承祯, 陈 灿, 等.闽东南海岸山地几种树种海拔资源位的叶绿素特征[J]. 福建农林大学学报, 2013, 42(1): 35-39.
[29] 薛 立, 曹 鶴. 逆境下植物叶性状变化的研究进展[J]. 生态环境学报, 2010, 19(8): 2 004-2 009.
[30] 王希群, 马履一, 贾忠奎, 等. 叶面积指数的研究和应用进展[J]. 生态学杂志, 2005, 24(5): 537-541.
[31] 黄海侠, 杨晓东, 孙宝伟, 等. 浙江天童常绿植物当年生与往年生叶片性状的变异与关联[J]. 植物生态学报, 2013, 37(10): 912-921.
[32] 李善家, 苏培玺, 张海娜, 等.荒漠植物叶片水分和功能性状特征及其相互关系[J]. 植物生理学报, 2013, 49(2): 153-160.
[33] Milla R, Reich P B. The scaling of leaf area and mass: the cost of light interception increases with leaf size[J]. Proceedings Biological Sciences, 2011, 274(1 622): 2 109-2 115.
[34] Weiner J. Allocation, plasticity and allometry in plants[J]. Perspectives in Plant Ecology Evolution & Systematics, 2004, 6(4): 207-215.
[35] Roth-Nebelsick A, Kerp H. Evolution and function of leaf venation architecture: A review [Review][J]. Annals of Botany, 2001, 87(5): 553-566.
[36] Yang D M, Zhang J J, Zhou D, et al. Leaf and twig functional traits of woody plants and their relationships with environmental change: A review[J]. Chinese Journal of Ecology, 2012, 31(3): 702-713.
[37] Mcdonald P G, Fonseca C R, Westoby M. Leaf-Size Divergence along Rainfall and Soil-Nutrient Gradients: Is the Method of Size Reduction Common among Clades?[J]. Functional Ecology, 2003, 17(1): 50-57.
[38] Williams K, Percival F, Merino J, Mooney H A. Estimation of tissue construction cost from heat of combustion and organic nitrogen content. Plant[J]. Cell & Environment, 1987, 10, 725-734.
[39] Wright I J, Wearoby M.Cross-species relationships between seedling relative growth rate, nitrogen productivity and root vs.leaf function in 28 Australian woody species[J]. Funct Ecol, 2000, 14(1): 97-107.
[40] Feng Y L, Fu G L, Zheng Y L. Specific leaf area relates to the differences in leaf construction cost, nitrogen allocation and use efficiency between invasive and noninvasive congeners of two genera[J]. Planta, 2008, 228(3): 383-390.
[41] And B F C, Hicks D J. The Ecology of Leaf Life Spans[J]. Annual Review of Ecology & Systematics, 1982, 13(1): 229-259.
