高原鼠兔干扰对青海湖流域高山嵩草草甸植物多样性及地上生物量的影响
2017-05-23金少红刘彤庞晓攀于成郭正刚
金少红,刘彤,庞晓攀,于成,郭正刚
(兰州大学草地农业科技学院,草地农业生态系统国家重点实验室,甘肃 兰州 730020)
高原鼠兔干扰对青海湖流域高山嵩草草甸植物多样性及地上生物量的影响
金少红,刘彤,庞晓攀,于成,郭正刚*
(兰州大学草地农业科技学院,草地农业生态系统国家重点实验室,甘肃 兰州 730020)
采用堵洞法设置高原鼠兔干扰区和非干扰区,然后在干扰区和非干扰区内利用样方法研究了高原鼠兔干扰对青海湖流域高山嵩草草甸植物多样性和地上生物量的影响。结果表明,植物物种多样性和不同植物功能群多样性对高原鼠兔干扰的响应并不一致,其中干扰增加了杂类草功能群丰富度指数与物种丰富度指数,降低了物种均匀度指数、莎草科功能群丰富度指数和禾草科功能群均匀度指数(P<0.05)。干扰区内物种均匀度指数、物种丰富度指数和杂类草功能群丰富度指数随干扰强度增加而呈增加趋势,但禾草科功能群均匀度指数却呈显著降低趋势。虽然高原鼠兔干扰显著降低了植物群落总生物量,莎草科、禾草科和豆科功能群生物量(P<0.05),但显著增加了杂类草功能群生物量(P<0.05)。干扰区随干扰程度增加,莎草科功能群和豆科功能群生物量逐渐降低,杂类草功能群生物量逐渐增加,而禾草科功能群生物量则先增加后降低。鉴于高山嵩草草甸植物群落物种多样性和功能多样性对高原鼠兔干扰的响应不一致,植物物种多样性和功能群多样性之间不能互相替代。
高原鼠兔;群落物种组分;物种多样性;功能群多样性;地上生物量
青海湖是我国面积最大的内陆高原咸水湖,它以巨大的水体与流域内的天然草地和森林共同形成了西部荒漠风沙东侵的生态屏障[1],因此青海湖流域的植被系统通过防风固沙、涵养水源、保持水土,保育生物多样性和调节气候等多种功能而维系着青藏高原东北部生态安全的保障[2-3]。青海湖流域草地面积占流域总面积比例超过72%[1],不仅是青海省畜牧业的主要载体[1],而且是生态屏障的核心组分[4]。
高山嵩草草甸是青海湖流域广泛分布的一种高寒草甸类型[5],草层低矮、植株密聚,虽然是优良放牧地[6],但也是草地小型啮齿类动物高原鼠兔适宜分布区[7]。高原鼠兔对高山嵩草草甸的作用往往随着其干扰程度的变化而发生明显的变化[8-10],适当高原鼠兔干扰能够提高高寒草甸的禾本科植物比例[9],增强植物群落的种间联结性[11],改变主要植物种群的分布格局[10]和植物群落组分以及不同组分的地位[12]。不同植物对高原鼠兔干扰的响应存在分异,说明高原鼠兔干扰改变了草地植物群落的物种多样性,然而一个草地植物群落内的不同植物间可能具有冗余性和功能相似性[13-14],其对环境敏感性的响应趋同[15],形成植物功能群,植物功能群对特定环境因素有相似的反应[16-18],其作为一个相对统一的整体对草地生态系统干扰做出反应[19],因此植物功能群被用于简化一定尺度内众多物种对外加干扰响应的研究[20],从而助于了解植物群落整合性、稳定和演替等生态过程[17]。然而理解高原鼠兔对高山嵩草草甸生态系统干扰过程时,能否用植物功能群代替植物多样性,尚需要科学的试验提供依据。
植物多样性是草甸生态系统变化的重要内容和标志[16,21],也是草地生态系统生产力维持的基础[22],因此要明晰高原鼠兔干扰对青海湖流域高山嵩草草甸影响的生态过程,首先需要查清植物多样性和地上生物量对高原鼠兔干扰的响应。因此本研究通过分析青海湖流域高原鼠兔干扰对植物物种多样性、功能群多样性以及群落内不同功能群地上生物量影响,阐明植物物种多样性和功能群多样性在响应高原鼠兔干扰的异同,探析能否用植物功能群多样性替代植物物种多样性分析高原鼠兔干扰对高山嵩草草甸的影响。
1 材料与方法
1.1 研究地区概况
青海湖流域位于青藏高原,四面环山,四周高中间低,形成一个封闭式内陆盆地。气候属内陆高原半干旱气候,高寒阴湿。年均温-3.0~3.0 ℃,绝对最高温26 ℃, 绝对最低温-35.8 ℃,年日照时数为2800~3100 h,年均降水量300~400 mm,其90%左右集中于5-9月[1],年蒸发量1440 mm左右。青海湖流域的草地类型以温性草原、高寒草原和高寒草甸为主,其中高寒草甸面积最大。高山嵩草草甸是高寒草甸的主要草地亚类[6],分布较广,以高山嵩草(Kobresiapygmaea)为优势种,常见伴生植物有矮生嵩草(K.humilis),乳白香青(Anaphalislactea),高原早熟禾(Poaalpigena),青藏苔草(Carexmoorcroftii)等。
1.2 研究方法
1.2.1 试验设计 2015年7月在青海湖流域黑马河(地理位置99°36′13′′ E,36°44′55′′ N,海拔3691 m)的冬季放牧地(牧草生长季不放牧),选择地势相对一致的高山嵩草草甸,夏季围栏没有放牧。高原鼠兔是群居性动物,喜欢栖息于开阔生境,以降低捕食风险。高原鼠兔干扰区与非干扰区有时是渐进的,而有时是镶嵌分布的。本研究高原鼠兔干扰区选择以现场观察是否有高原鼠兔洞口和高原鼠兔出没为准,若同时发现有高原鼠兔洞口和高原鼠兔出没,则选定为高原鼠兔干扰区。为避免草地类型和地貌地形对研究结果的影响,根据干扰区的草地类型和地形地貌在干扰区周围选择非干扰区,非干扰区选择以几乎没有高原鼠兔洞口和高原鼠兔出没为准,该方法已成功应用于土拨鼠[23]、獾和狐狸[24]的研究。最后选择的高原鼠兔干扰区和非干扰区距离大约3 km,草地植物群落的优势种一样,被判定为草地类型一样,然而干扰区内优势植物的优势度要比非干扰区内优势植物的优势度低。选定好高原鼠兔干扰区和非干扰区后,分别随机在干扰区和非干扰区内设置10个25 m×25 m(625 m2) 的样地,样地间距离大于25 m,然后用顶端有红漆的木桩四角标记,并采用塑料绳标记样地四周。针对干扰区内的样地,利用连续3 d堵洞法测定各样地的有效洞口数[2],以3 d测得的平均值作为该区高原鼠兔干扰水平,其分别为 2,5,7,9,11,13,15,26,29,32,在每个样地内,在两条对角线的交汇处先设置一个样方,然后沿任意一条对角线,从交汇处向外延伸10 m,再分别设置2个样方,样方面积为1 m×1 m。
1.2.2 样方调查 样地内高原鼠兔有效洞口数调查完成的次日,开始样方调查。记录每个样方中出现的植物种类,然后测定物种的高度和盖度。若样方中某一物种的株(丛)数超过30,则随机选择30株利用钢卷尺测定其株高,若样方内某一物种的株(丛)数不足30株则全部测定;物种盖度采用针刺法测定,然后分种收获每个物种的地上生物量,分种装至信封袋,带回实验室烘干称重[10]。根据青藏高原已有植物功能群的划分实例[22],本研究依据不同植物的生活型,将其分别划分为莎草科(Cyperaceae)、禾草科(Graminea)、豆科(Leguminosae)和杂类草(Weed) 4个功能群。
1.2.3 指标计算 物种重要值的计算采用公式:
IV=(HR+CR+FR)/3
(1)
式中:IV为重要值;HR为相对高度;CR为相对盖度;FR为相对生物量[25]。
物种多样性测度采用多样性指数、均匀度指数和丰富度指数,其中多样性指数采用Shannon-Wiener指数(H),其计算公式:
H=-∑PilnPi
(2)
式中:H为多样性指数;Pi是种的相对重要值。
丰富度指数(S)用Patrick指数,
S=样方内出现的物种总数
(3)
均匀度指数(J)采用Pielou,其计算公式:
J=H/ln(S)
(4)
植物功能群多样性的测度也采用多样性指数、均匀度指数和丰富度指数,其中多样性指数采用Shannon-Wiener指数(H′),其计算公式:
H′=-∑Pi′lnPi′
(5)
式中:H′为功能群多样性指数;Pi′是指该功能群内所有种的相对重要值。
功能群丰富度指数(S′)用Patrick指数,
S′=样方内该功能群中所包含的物种总数
(6)
功能群均匀度指数(J′)采用Pielou,其计算公式:
J′=H′/ln(S′)[4]
(7)
1.3 数据分析
先采用独立样本t检验,检验高原鼠兔干扰对指标的影响差异是否显著,若差异不显著, 则不进行进一步分析;若差异显著(P<0.05),再采用R-3.3.2-windows进行相关性检验。
2 结果与分析
2.1 高原鼠兔干扰对高山嵩草草甸植物群落组分及其重要值的影响
高原鼠兔干扰对高山嵩草草甸植物群落组分具有一定的影响,表现为干扰区样方较非干扰区样方的物种数增多(表1),增加的物种主要是节节草和委陵菜。高原鼠兔干扰没有改变高山嵩草在植物群落内的地位和作用,表现为干扰区高山嵩草的重要值虽然有所下降,但其仍然位列所有物种之首,仍然为优势种。高原鼠兔干扰明显改变了主要伴生种在群落内的地位和作用,如乳白香青、达乌里秦艽、麻花艽等杂草的重要值在群落内的排序上升,说明其在群落内的地位有所增加;矮生嵩草、青藏苔草、车前、青海黄芪、扁蓿豆等植物重要值的排序降低,说明其在群落内的地位和作用有所下降,但垂穗披碱草、荷叶凤毛菊等植物的重要值在群落内的排序变化不大,说明其在群落内的地位和作用变化不明显。

表1 高原鼠兔干扰对物种组成及其重要值的影响Table 1 The compositions and important value of plant species under different effective burrow entrance densities
注:干扰区和非干扰区样方内的物种数不一样,此表列出的是干扰区和非干扰区内有19个样方均出现的植物种,若某植物种出现样方数低于19个,则没有列出。表中数值为某物种各个样方重要值的平均值。
Note: The number of species disturbed area and undisturbed area are not the same. The table is a list of the disturbed area and undisturbed area which has 19 species plots appeared, if a species of each number less than 19, are not listed. The data in the table is the average value of a species important value with each subplot.
2.2 高原鼠兔干扰对高山嵩草草甸物种多样性和总生物量的影响
高原鼠兔干扰对物种多样性指数没有明显影响,但其影响了物种丰富度指数、均匀度指数和植物群落总生物量(图1)。高原鼠兔干扰显著降低了植物群落总生物量(F=3.317,df=16,sig(双侧)=0.000)和均匀度指数(F=3.154,df=16,sig(双侧)=0.035),但增加了丰富度指数(F=0.543,df=16,sig(双侧)=0.001)。
高原鼠兔不同干扰程度对植物群落总生物量没有显著影响(图2),但明显影响了物种均匀度指数和物种丰富度指数(图3)。干扰区内,随高原鼠兔有效洞口密度增加,物种均匀度指数和物种丰富度指数整体均表现为增加趋势(图3)。

图1 鼠兔干扰对物种多样性和生物量的影响Fig.1 The species diversity and biomass under different effective burrow entrance densities
2.3 高原鼠兔干扰对高山嵩草草甸不同植物功能群
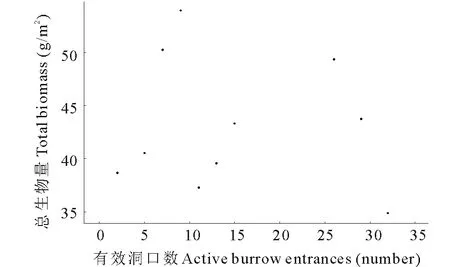
图2 干扰区不同高原鼠兔干扰水平下植物生物量的变化Fig.2 The plant biomass in disturbed area under different disturbance levels of plateau pika
多样性及其生物量的影响
高原鼠兔干扰对莎草科功能群多样性指数和均匀度指数(图4)、禾草科功能群多样性指数和丰富度指数(图5)、豆科功能群多样性指数、丰富度指数和均匀度指数(图6)、杂类草功能群多样性指数和丰富度指数没有明显影响(图7),但其影响了莎草科功能群丰富度指数和生物量(图4)、禾草科功能群均匀度指数和生物量(图5)、豆科功能群生物量(图6)、杂类草功能群丰富度指数和生物量(图7)。高原鼠兔干扰显著降低了莎草科功能群的丰富度指数(F=0.000,df=16,sig(双侧)=0.035)和生物量(F=4.875, df=12.468,sig(双侧)=0.000)(图4),禾草科功能群的均匀度指数(F=2.866, df=16,sig(双侧)=0.039)和生物量(F=5.611,df=9.5,sig(双侧)=0.000)(图5),以及豆科功能群的生物量(F=18.034,df=10,sig(双侧)=0. 007)(图6),但增加了杂类草功能群的丰富度指数(F=1.048,df=16,sig(双侧)=0.023)和生物量(F=21.024,df=8,sig(双侧)=0.013)(图7)。

图3 高原鼠兔干扰区不同干扰水平下物种丰富度指数和均匀度指数的变化Fig.3 The species richness index and the species evenness index in disturbed area under different disturbance levels of plateau pika
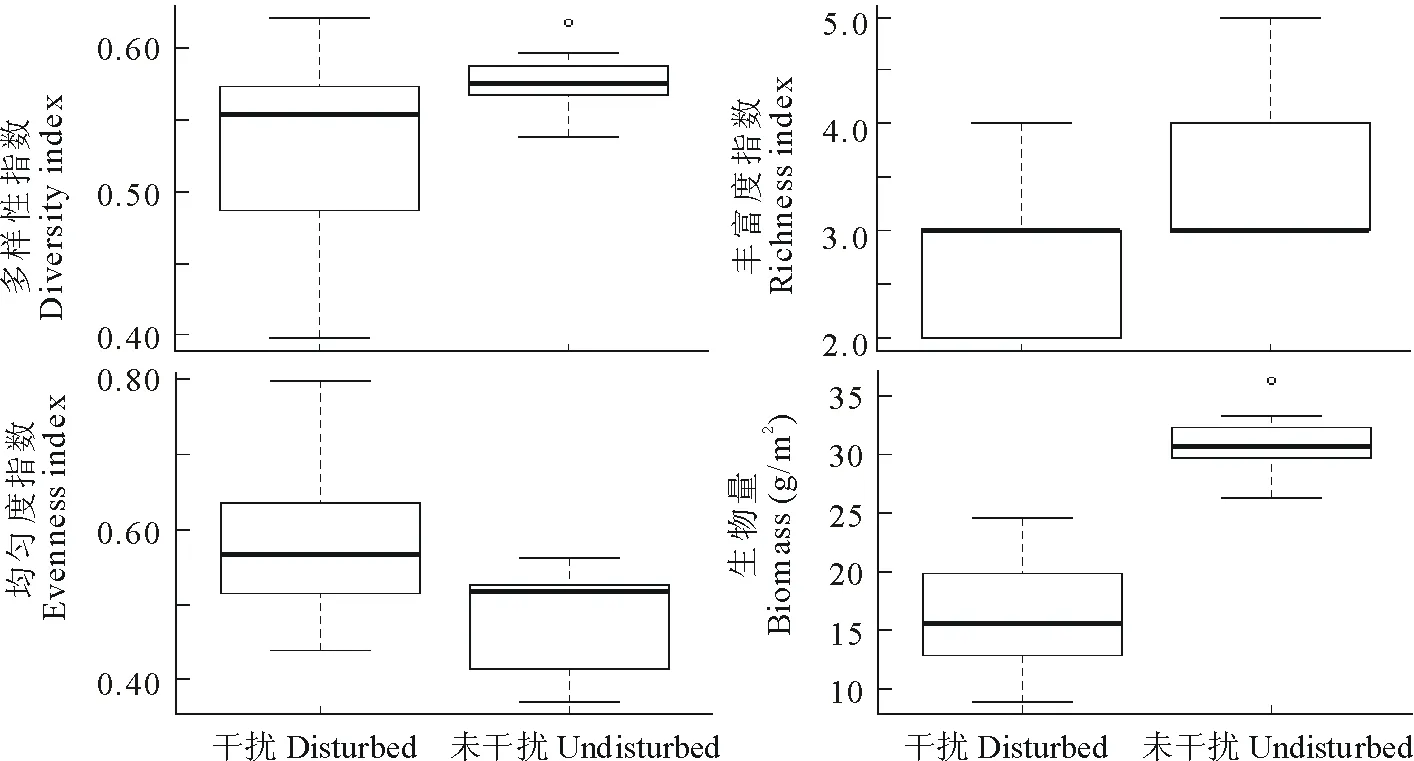
图4 鼠兔干扰对莎草科多样性及生物量的影响Fig.4 The diversity and biomass of Cyperaceae under different effective burrow entrance densities

图5 鼠兔干扰对禾草科多样性及生物量的影响Fig.5 The diversity and biomass of Graminea under different effective burrow entrance densities

图6 鼠兔干扰对豆科多样性及生物量的影响Fig.6 The diversity and biomass of Leguminous under different effective burrow entrance densities

图7 鼠兔干扰对杂类草多样性及生物量的影响Fig.7 The diversity and biomass of weed under different effective burrow entrance densities

图8 干扰区不同高原鼠兔干扰水平下莎草科丰富度指数的变化Fig.8 The richness index of Cyperaceae in disturbed area under different disturbance levels of plateau pika
虽然高原鼠兔干扰影响了莎草科功能群丰富度指数和生物量、禾草科功能群均匀度指数和生物量、豆科功能群生物量、杂类草功能群丰富度指数和生物量,但不同高原鼠兔干扰程度对莎草科功能群丰富度指数没有显著影响(图8),而不同高原鼠兔干扰程度明显影响了禾草科均匀度指数(图9)和生物量(图10)、豆科功能群生物量、杂类草功能群丰富度指数(图9)和生物量(图10)。

图9 高原鼠兔干扰区不同干扰水平下杂类草丰富度指数和禾草科均匀度指数Fig.9 The richness index of weed and the evenness index of Gramineae in disturbed area under different disturbance levels of plateau pika
高原鼠兔干扰区内,随高原鼠兔有效洞口密度增加,禾草科均匀度指数整体表现为降低趋势(图9),而杂类草功能群的丰富度指数整体表现为增加趋势。不同功能群植物生物量对高原鼠兔干扰的响应不一致。随高原鼠兔干扰强度增加,莎草科功能群生物量、豆科功能群生物量整体表现为降低趋势(图10),禾草科功能群生物量则先增加后降低,在中度干扰程度时生物量较高(图10),但杂类草功能群生物量随高原鼠兔干扰强度增加呈现增加态势(图10)。

图10 干扰区不同高原鼠兔干扰水平下各植物功能群生物量的变化Fig.10 The biomass of different plant functional groups in disturbed area under different disturbance levels of plateau pika
3 讨论
高原鼠兔是高寒草甸生态系统中小型动物的优势种之一[7,26-27],其不仅通过采食直接影响植物生长发育[28],而且通过挖掘、排泄粪尿以及过剩食物碎屑堆积等作用[29],增加小生境的异质性[30],间接地影响植物生长发育[31]。高原鼠兔活动可能会促进某些植物生长,也可能抑制某些植物生长,从而改变植物群落的物种组分。研究表明,高原鼠兔干扰不仅改变了高寒草甸植物群落内的物种数,而且改变了原有物种在植物群落内的地位和作用。一方面高原鼠兔干扰增加了青海湖流域高山嵩草草甸的植物物种数,但增加的物种主要是杂类草,这与高原鼠兔干扰对甘南自治州玛曲县高寒草甸植物群落组分的影响趋同[12],这说明高原鼠兔干扰虽然能够丰富高寒草甸内的植物种类,但从草原放牧利用角度,增加的植物多为放牧季家畜不直接采食的植物;另一方面,高原鼠兔干扰后优势种高山嵩草重要值虽有所下降,但仍位列所有物种之首,表明群落组分的变化属于群落内部波动,群落性质没有发生变化,因此高原鼠兔干扰主要改变了主要伴生种重要值的排序,从而改变了这些物种在群落内的地位和作用[12,32],譬如高原鼠兔干扰提前了乳白香青、达乌里秦艽等旱生和中旱生植物重要值的排序,说明高原鼠兔干扰加强了旱生和中旱生植物在群落内的地位,降低了矮生嵩草等植物重要值的排序,说明中生植物在群落内的地位下降,主要是高原鼠兔活动会形成大量裸斑,镶嵌于高寒草甸内,降低了土壤含水量[33],生境旱化所致[11]。研究表明,高原鼠兔干扰增加了物种丰富度指数,这与栉鼠干扰对阿根廷Puna荒漠草原植物丰富度的影响趋同[34],主要是高原鼠兔干扰一定程度上改变了草地植物的生境[35],虽然仅西藏嵩草(K.tibetica)衰退至退出草地植物群落,但不同物种因其在群落内的地位发生变化,不同植物种适应生境存在差异[11],因此植物群落内不同物种的分布较原来群落分布发生一定的变化,同时高原鼠兔干扰后部分先锋种随机入侵高原鼠兔干扰区内的微环境[36],因此物种的丰富度指数有所增加,但物种均匀度指数有所降低,而物种多样性指数的变化与丰富度指数和均匀度指数的变化不一致,主要是物种多样性指数既反映群落内种的绝对密度,又反映种的相对密度[9],丰富度指数和均匀度指数变化过程的不一致性,导致多样性指数变化不大。
虽然植物群落内物种较多,但有些植物物种在草甸生态系统内的某一方面具有相似的功能,这些具有相似功能的植物间具有冗余作用,一个物种的消失或增加,有可能减弱或增强植物的某一功能,但不会使植物群落的某些功能丧失。高原鼠兔干扰显著增加了杂类草功能群的丰富度指数,降低了莎草科功能群的丰富度指数和禾草科功能群的均匀度指数,主要是高原鼠兔活动增加了环境异质性[12,37],鹅绒委陵菜等适应异质性资源较强的可塑性物种侵入干扰区[38],但莎草科功能群内的西藏嵩草因环境变化而退出群落,而禾草科功能群落内扁穗冰草等物种适应性增加,而高原早熟禾适应性降低,从而使禾草科功能群内物种均匀度指数降低。高原鼠兔干扰显著降低植物群落总生物量,但不同有效洞口密度下的群落总生物量没有显著变化,这与高原鼠兔干扰对甘南自治州玛曲县高寒草甸植物生物量的影响趋同[11]。然而高原鼠兔干扰显著影响了青海湖流域草地植物不同功能群生物量,杂类草功能群生物量增加,莎草科、禾草和豆科功能群的生物量均降低,主要是高原鼠兔干扰后高山嵩草草甸裸斑面积增大,生境旱化[11],湿生的高山嵩草、中生的高原早熟禾等植物生物量减少,而一些阔叶植物如达乌里秦艽和莓叶委陵菜等杂类草的生物量增加,从而导致莎草科和禾草科生物量下降,但杂草生物量增加量比禾草和莎草生物量降低量小,从而导致群落总生物量降低。在干扰区,中度干扰程度时禾草科功能群生物量较高,这说明适度高原鼠兔扰动有利于增加优良牧草的产量,提高草甸放牧质量,这与高原鼠兔干扰对青海湖流域内沙柳河河岸高寒草甸禾草科生物量的影响趋同[39]。
本研究结果表明,高山嵩草草甸植物群落物种多样性和功能多样性对高原鼠兔干扰的响应不一致,因此在研究高原鼠兔干扰高寒草甸时,植物物种多样性和功能多样性之间不能互相替代。植物物种多样性是功能群多样性的保障,而功能群多样性是物种多样性的体现和载体[40],植物物种多样性和功能群多样性间虽然存在联系,但其揭示高寒草甸的核心目标存异,植物物种多样性强调的是某一物种存在的价值,其核心是生物多样性保育,而功能群多样性强调的是物种存在与否对高寒草地生态系统功能的影响,因此评价某一物种的减少对生态系统的影响时,还需要考虑植物群落中其他具有类似功能的物种[13,15],因此高原鼠兔干扰下植物功能群内会丧失(增加)某些物种,但由于其他冗余种存在产生的补充(保险)效果,使得这些物种的丧失(增加)不足以在短期内导致某个生态功能的缺失(出现)[14],即群落中具有相似功能的物种的存在,在一定程度上降低了干扰后由于物种缺失(增加)造成的高山嵩草草甸生态系统功能缺失(出现)的可能性,因此不能仅以物种变化特征简单地推断出功能群的变化。不同功能群内的植物物种对高原鼠兔干扰的敏感性不同[4],它们在群落中的作用也各有不同,多数生态系统的功能和作用在很大程度上由群落优势种决定,功能群的研究在一定程度上淡化了植物个体对环境适应性的差异,无法呈现群落不同植物物种之间相互作用引起的群落特征的变化,所以,也不能以功能群的变化来推断群落对高原鼠兔干扰的响应。因此,研究高原鼠兔干扰高寒草甸生态系统时,若主要考虑生物多样性保育,建议采用物种多样性描述,而若主要考虑草甸生态系统功能时,建议采用功能群描述。
References:
[1] Feng Z W, Feng Z Z. Major ecological and environmental problems and counter measures in the Lake watershed, Qinghai. Ecology and Environment, 2004, 13(4): 467-469. 冯宗炜, 冯兆忠. 青海湖流域主要生态环境问题及防治对策. 生态环境, 2004, 13(4): 467-469.
[2] Chun M. The evaluation of natural grassland on ecological value in Nangqian County. Qinghai Prataculture, 2010, 19(3): 31-36. 春梅. 囊谦县高寒草甸类草地生态价值评价. 青海草业, 2010, 19(3): 31-36.
[3] Zhang X S. Eco-economic functions of the grassland and its patterns. Science and Technology Review, 2000, 9(8): 3-7. 张新时. 草地的生态经济功能及其范式. 科技导报, 2000, 9(8): 3-7.
[4] Shi H X, Hou X Y, Shi S L,etal. Relationships between plant diversity, soil property and productivity in an alpine meadow. Acta Prataculturae Sinica, 2015, 24(10): 40-47. 石红霄, 侯向阳, 师尚礼, 等. 高山嵩草草甸初级地上生物量、多样性与土壤因子的关系. 草业学报, 2005, 24(10): 40-47.
[5] Zhang Y, Dong S K, Gao Q Z,etal. Responses of alpine vegetation and soils to the disturbance of plateau pika (Ochotonacurzoniae) at burrow level on the Qinghai-Tibetan Plateau of China. Ecological Engineering, 2016, 88: 232-236.
[6] Li X Q. The research on the ecological function position of Qinghai Qilian Mountain Area. Qinghai Prataculture, 2009, 18(4): 19-23. 李旭谦. 青海湖流域草地类型及其分布. 青海草业, 2009, 18(4): 19-23.
[7] Zhou X R, Guo Z G, Guo X H. The role of plateau pika and plateau zokor in alpine meadow. Pratacultural Science, 2010, 27(5): 38-44. 周雪荣, 郭正刚, 郭兴华. 高原鼠兔和髙原鼢鼠在髙寒草甸中的作用. 草业科学, 2010, 27(5): 38-44.
[8] Sun F D, Chen W Y, Liu L,etal. Effects of plateau pika activities on seasonal plant biomass and soil properties in the alpine meadow ecosystems of the Tibetan Plateau. Grassland Science, 2015, 61(4): 195-203.
[9] Guo Z G, Li X F, Liu X Y,etal. Response of alpine meadow communities to burrow density changes of plateau pika (Ochotonacurzoniae) in the Qinghai-Tibet Plateau. Acta Ecologica Sinica, 2012, 32(1): 44-49.
[10] Guo Z G, Zhou X R, Hou Y. Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on soil physicochemical property of the bare land and vegetation land in the Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 2012, 32: 104-110.
[11] Pang X P, Wang Q, Jia T T,etal. Effect of burrow entrance densities of plateau pika (Ochotonacurzoniae) on interspecific association inKobresiapygmaeameadow. Acta Prataculturae Sinica, 2015, 24(5): 224-230. 庞晓攀, 王倩, 贾婷婷, 等. 高原鼠兔有效洞口数密度对高山嵩草草甸植物种间联结性的影响. 草业学报, 2015, 24(5): 224-230.
[12] Jia T T, Mao L, Guo Z G. Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on plant niche of alpine meadow communities in the Qinghai-Tibet Plateau. Acta Ecologica Sinica, 2014, 34(4): 869-877. 贾婷婷, 毛亮, 郭正刚. 高原鼠兔有高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响. 生态学报, 2014, 34(4): 869-877.
[13] Guo Q L, Lu X L, Ding S Y. The classification of plant functional types based on the dominant herbaceous species in the riparian zone ecosystems in the Yiluo River. Acta Ecologica Sinica, 2012, 32(14): 4434-4442. 郭屹立, 卢训令, 丁圣彦. 伊洛河河岸带生态系统草本植物功能群划分. 生态学报, 2012, 32(14): 4434-4442.
[14] Hu N, Fan Y L, Ding S Y,etal. Progress in researches on plant functional groups of terrestrial ecosystems. Acta Ecologica Sinica, 2008, 8(7): 3302-3311. 胡楠, 范玉龙, 丁圣彦, 等. 陆地生态系统植物功能群研究进展. 生态学报, 2008, 28(7): 3302-3311.
[15] Zang R G, Zhang Z D. Plant functional groups and their dynamics in tropical forests: a review. Acta Ecologica Sinica, 2010, 30(12): 3289-3296. 臧润国, 张志东. 热带森林植物功能群及其动态研究进展. 生态学报, 2010, 30(12): 3289-3296.
[16] Wang C T, Long R J, Ding L M. The effects of differences in functional group diversity and composition on plant community productivity in four types of alpine meadow communities. Biodiversity Science, 2004, 12(4): 403-407. 王长庭, 龙瑞军, 丁路明. 高寒草甸不同草地类型功能群多样性及组成对植物群落地上生物量的影响. 生物多样性, 2004 12(4): 403-407.
[17] Yang X H, Bao J Y, Han G D,etal. Plant functional groups and their applications in ecology research. Journal of Dalian Nationalities University, 2009, 11(5): 397-409. 杨晓慧, 鲍雅静, 韩国栋, 等. 植物功能群及其在生态学研究中的应用. 大连民族学院学报, 2009, 11(5): 397-409.
[18] Fan Y L, Hu N, Ding S Y,etal. The classification of plant functional types based on the dominant herbaceous species in forest ecosystem at Funiu Mountain. Acta Ecologica Sinica, 2008, 28(7): 3092-3101. 范玉龙, 胡楠, 丁圣彦, 等. 伏牛山自然保护区森林生态系统草本植物功能群的分类. 生态学报, 2008, 28(7): 3092-3101.
[19] Zheng W, Zhu J Z, Pan C D. Effect of grazing disturbance on plant functional group and community structure of meadow community in Kanas Scenic Area. Chinese Journal of Grassland, 2010, 32(1): 92-98. 郑伟, 朱进忠, 潘存德. 放牧干扰对喀纳斯草地植物功能群及群落结构的影响. 中国草地学报, 2010, 32(1): 92-98.
[20] Vitousek P M, Hooper D U. Biological diversity and terrestrial ecosystem biochemistry[M]//Biodiversity and Ecosystem Function. Berlin Heidelberg: Springer-Verlag, 1994: 3-14.
[21] Lu X L, Liang G F, Tang Q,etal. Plant species of the non-agricultural habitats in the lower reaches of the Yellow River plain agro-landscape. Acta Ecologica Sinica, 2014, 34(4): 789-797. 卢训令, 梁国付, 汤茜, 等. 黄河下游平原农业景观中非农生境植物多样性. 生态学报, 2014, 34(4): 789-797.
[22] Han G D, Jiao S Y, Bi Li G T,etal. Effects of plant species diversity and productivity under different stocking rates in theStipabrevifloraGriseb desert steppe. Acta Ecologica Sinica, 2007, 27(1): 182-188. 韩国栋, 焦树英, 毕力格图, 等. 短花针茅草原不同载畜率对植物多样性和草地地上生物量的影响. 生态学报, 2007, 27(1): 182-188.
[23] Chen W Y, Zhao M, Li G Y,etal. The influence of different types of fertilizer application level on the Gannan desertification of alpine meadow of plant characteristics and the productive forces. Journal of Natural Resources, 2012, 27(2): 254-267. 陈文业, 赵明, 李广宇, 等. 不同类型施肥水平对甘南沙化高寒草甸植物群落特征及地上生物量的影响. 自然资源学报, 2012, 27(2): 254-267.
[24] Barth C J, Liebig M A, Hendrickson J R,etal. Soil change induced by prairie dogs across three ecological sites. Soil Science Society of America Journal, 2014, 78(6): 2054-2060.
[25] Kurek P, Kapusta P, Holeksa J. Burrowing by badgers (Melesmeles) and foxes (Vulpesvulpes) changes soil conditions and vegetation in a European temperate forest. Ecological Research, 2014, 29(1): 1-11.
[26] Sun F D, Long R J, Lu C X. Effects of plateau pikas (Ochotonacurzoniae) burrow densities on plant community composition and population diversity in alpine meadow. Journal of Arid Land Resources and Environment, 2010, 24(7): 181-186. 孙飞达, 龙瑞军, 路承香. 高原鼠兔不同洞穴密度对高寒草地植物群落组成及多样性的影响. 干旱区资源与环境, 2010, 24(7): 181-186.
[27] Su J H, Nan Z B, Ji W H. Effects of livestock grazing on rodents in grassland ecosystems. Acta Prataculturae Sinica, 2016, 25(11): 136-148. 苏军虎, 南志标, 纪维红. 家畜放牧对草地啮齿动物影响的研究进展. 草业学报, 2016, 25(11): 136-148.
[28] Sun F D, Long R J, Guo Z G,etal. Effects of rodents activists on plant community and soil environment in alpine meadow. Pratacultural Science, 2011, 28(1): 146-151. 孙飞达, 龙瑞军, 郭正刚, 等. 鼠类活动对高寒草甸植物群落及土壤环境的影响. 草业科学, 2011, 28(1): 146-151.
[29] Pech R P, Arthur A D, Zhang Y M,etal. Population dynamics and responses to management of plateau pikas(Ochotonacurzoniae). Journal of Applied Ecology, 2007, 44(3): 615-624.
[30] Wesche K, Nadrowski K, Retzer V. Habitat engineering under dry conditions: The impact of pikas (Ochotonapallasi) on vegetation and site conditions in southern Mongolian steppes. Journal of Vegetation Science, 2007, 18(5): 665-674.
[31] Li W J, Zhang Y M. Impacts of plateau pikas on soil organic matter and moisture content in alpine meadow. Acta Theriologica Sinica, 2006, 26(4): 331-337. 李文靖, 张堰铭. 高原鼠兔对高寒草甸土壤有机质及湿度的作用. 兽类学报, 2006, 26(4): 331-337.
[32] Xi W L, An S Z, Jin F L,etal. Change of community structure of altocherbiprata under different utilization ways. Pratacultural Science, 2010, 27(5): 15-18. 锡文林, 安沙舟, 靳发兰, 等. 不同利用方式下山地高草草甸群落结构的变化. 草业科学, 2010, 27(5): 15-18.
[33] Zhang Y M, Liu J K. Effects of plateau zokors (Myospalaxfontanierii) on plant community and soil in an alpine meadow. Journal of Mammalogy, 2003, 84(2): 644-651.
[34] Natalia L, Paola S, Carlos E B. Effect of herbivory and disturbances by Tuco-Tucos (CtenomysMendocinus) on a plant community in the Southern Puna Desert. Arctic, Antarctic and Alpine Research, 2007, 39(1): 110-116.
[35] Smith A T, Foggin J M. The plateau pika (Ochotonacurzoniae) is a keystone species for biodiversity on the Tibetan Plateau. Animal Conservation, 1999, 2(4): 235-240.
[36] Wu R X, Chai Q, Zhang J Q,etal. Impacts of burrows and mounds formed by plateau rodents on plant species diversity on the Qinghai-Tibetan Plateau. Rangeland Journal, 2015, 37(1): 117-123.
[37] Wang Q J, Wang W Y, Jing Z C,etal. Effects of vegetation cover change of alpineKobersiameadow on plant community structure and diversity in source region of the Yangtze and Yellow River. Resources Science, 2006, 28(2): 118-124. 王启基, 王文颖, 景增春, 等. 江河源区高山嵩草草甸覆被变化对植物群落特征及多样性的影响. 资源科学, 2006, 28(2): 118-124.
[38] Shen Z X, Chen Z Z, Wang Y H. Clonal growth of stoloniferous herbPotentillaanserinaon degraded and non-degraded alpine meadow soil. Chinese Journal of Applied Ecology, 2003, 14(8): 1332-1336. 沈振西, 陈佐忠, 王彦辉. 高寒退化与未退化草甸土壤下匍匐茎鹅绒萎陵菜的克隆生长特征的比较. 应用生态学报, 2003, 14(8): 1332-1336.
[39] Zhao G Q, Li G Y, Ma W H,etal. Impacts ofOchotonapallasidisturbance on alpine grassland community characteristics. Chinese Journal of Applied Ecology, 2013, 24(8): 2122-2128. 赵国琴, 李广泳, 马文虎, 等. 高原鼠兔扰动对高寒草地植物群落特征的影响. 应用生态学报, 2013, 24(8): 2122-2128.
[40] Sun G Y, Zhang R, Zhou L. Trends and advances in researches on plant functional diversity and functional groups. Acta Ecologica Sinica, 2003, 23(7): 1430-1435. 孙国钧, 张荣, 周立. 植物功能多样性与功能群研究进展. 生态学报, 2003, 23(7): 1430-1435.
Effects of plateau pika (Ochotonacrzoniae) disturbances on plant species diversity and aboveground plant biomass in aKobresiapygmaeameadow in the Qinghai Lake Region
JIN Shao-Hong, LIU Tong, PANG Xiao-Pan, YU Cheng, GUO Zheng-Gang*
CollegeofPastoralAgricultureScienceandTechnology,LanzhouUniversity,StateKeyLaboratoryofGrasslandAgro-ecosystem,Lanzhou730020,China
A field survey was carried out to determine the effects of plateau pika disturbances on plant species diversity and productivity in aKobresiapygmaeameadow in the Qinghai Lake Region. A random stratified paired sampling design was used to select a disturbed area containing burrow entrances and plateau pika simultaneously, and an undisturbed area without burrow entrances or plateau pika. The field experiment was conducted in early May, 2015. The dominant plant was the same in the disturbed and undisturbed areas, but its degree of dominance was lower in the disturbed area than in the undisturbed area. Ten plots (25 m×25 m) were randomly placed with at least 25 m distance between plots. Each plot was fenced to exclude other wild herbivores. In each plot, vegetation was sampled in three subplots on the diagonal (each 1 m×1 m, with at least 10 m distance between subplots). The results showed that plateau pika disturbances had different effects on species diversity and plant functional groups. Plateau pika disturbances increased the richness index of weeds and plant species richness, and decreased the plant evenness index, the richness index of Cyperaceae, and the evenness index of Gramineae (P<0.05). In disturbed areas, plant species richness and evenness and the richness index of weeds gradually increased with increasing plateau pika disturbance levels, while the evenness index of Gramineae decreased. Plateau pika disturbances significantly reduced the total community biomass (P<0.05), but the trends in the changes of biomass differed among functional groups. Plateau pika disturbances significantly decreased the biomass of Cyperaceae, Gramineae, and Leguminosae, but increased the biomass of weeds (P<0.05). With increasing plateau pika disturbance levels, the biomass of Cyperaceae and Leguminosae decreased, the biomass of weeds increased, and the biomass of Gramineae first increased and then decreased. These results suggested that analyses of plant diversity cannot replace analyses of plant functional diversity when trying to understand the responses of alpine meadows to plateau pika disturbances.
plateau pika; species composition; species diversity; functional group diversity; aboveground plant biomass
10.11686/cyxb2016391
http://cyxb.lzu.edu.cn
2016-10-24;改回日期:2016-12-23
国家重点研发计划(2016YFC0502005),国家行业(农业)公益项目(201203041)和国家自然科学基金(31172258)资助。
金少红(1994-),女,甘肃兰州人,在读硕士。E-mail:jinshh12@lzu.edu.cn*通信作者Corresponding author. E-mail: guozhg@lzu.edu.cn
金少红, 刘彤, 庞晓攀, 于成, 郭正刚. 高原鼠兔干扰对青海湖流域高山嵩草草甸植物多样性及地上生物量的影响. 草业学报, 2017, 26(5): 29-39.
JIN Shao-Hong, LIU Tong, PANG Xiao-Pan, YU Cheng, GUO Zheng-Gang. Effects of plateau pika (Ochotonacrzoniae) disturbances on plant species diversity and aboveground plant biomass in aKobresiapygmaeameadow in the Qinghai Lake Region. Acta Prataculturae Sinica, 2017, 26(5): 29-39.
