树鼩中APP基因特征描述及可变剪接体的分型鉴定
2017-05-18罕园园孙晓梅匡德宣陆彩霞陈玲霞仝品芬王文广李娜代解杰
罕园园,孙晓梅,匡德宣,陆彩霞,陈玲霞,仝品芬,王文广,李娜,代解杰
(中国医学科学院北京协和医学院医学生物学研究所树鼩种质资源中心,云南省重大传染病疫苗研发重点实验室,中国医学科学院医学生物学研究所实验树鼩标准化与应用研究省创新团队,昆明 650118)
研究报告
树鼩中APP基因特征描述及可变剪接体的分型鉴定
罕园园,孙晓梅,匡德宣,陆彩霞,陈玲霞,仝品芬,王文广,李娜,代解杰*
(中国医学科学院北京协和医学院医学生物学研究所树鼩种质资源中心,云南省重大传染病疫苗研发重点实验室,中国医学科学院医学生物学研究所实验树鼩标准化与应用研究省创新团队,昆明 650118)
目的 区分并鉴定树鼩中APP基因mRNA的多种可变剪接体,对APP基因特征进行描述,并确定其在各组织中的表达分布。方法 参考已知人的和树鼩基因组预测的APP基因序列,设计树鼩APP基因mRNA剪切体外显子的共同特异性引物。分别从树鼩不同组织中提取总RNA,反转录为cDNA,利用高保真酶扩增目的剪切体DNA。根据PCR扩增产物电泳条带的有无和大小初步判断剪接体的型别,最后将PCR产物胶回收进行测序鉴定,对获得的基因进行特征描述,并结合定量PCR的结果确定了各剪接体在不同组织中分布情况。结果 结果表明树鼩APP剪接体的全长为3514 bp,有一个109 bp的5’-UTR,1092bp的3’-UTP。APP基因在调查的9个物种中高度同源保守,显示树鼩与灵长类存在一个较近的亲缘关系。通过三维建模获得了树鼩和人的APP基因共同拥有的4个结构域。同时确认这4种通过外显子跳跃产生的APP可变剪接体在不同组织中的分布和表达。4种检测到的剪接体APP770,APP695,APP751,APP677均表达于肺、肾和肠,表达量最高的是肺、肌肉和睾丸。结论 对树鼩APP基因可变剪接体的表达研究,有助于推动树鼩作为阿尔茨海默病模型深入研究疾病的发生机制和药物研发。
树鼩;APP基因;可变剪接体;阿尔茨海默病模型
淀粉样前体蛋白(amyloid precursor protein, APP)是中枢神经系统广泛表达的跨膜蛋白。它是阿尔茨海默病(Alzheimer’s disease, AD)病理改变老年斑的主要成分β淀粉蛋白(amyloid beta protein, Aβ)的前体蛋白。Aβ聚集形成的寡聚体具有神经毒性,被认为是AD的主要病理机制,这些过量产生的Aβ主要来源于APP的异常裂解,因此,APP一直被认为是是阿尔兹海默病AD形成和发展的关键因素[1]。在人体内,已经发现10种APP 基因的转录物,它们主要由三个不同的外显子(外显子2、7、8、15)经可变剪接所产生。APP 的表达非常广泛,可存在于几乎所有的神经元和非神经元组织。但是,各种转录物的表达方式显示一定的细胞和组织特异性。如含有Kunitz型蛋白酶抑制剂(Kunitz protease inhibitor, KPI)序列的APP751 和APP770 主要表达于非神经元组织,而不含KPI 序列的APP695则主要表达于神经元。另外,在脑发育、成熟过程及部分AD 病人,剪接形式也可发生变化[2]。
APP基因的序列、结构对Aβ的聚集有重要的作用,基于人基因组中发现的家族性APP基因突变而建立的转基因小鼠由于能够过表达APP基因,从而造成Aβ的聚集沉淀,形成类似人AD中最主要的病理特征—老年斑(senile plaques, SP),是现在研究AD的重要动物模型。Aβ形成年龄依赖性的SP并在脑相关区域聚集沉淀,这种现象在许多的哺乳动物中都有发现,包括熊、牛、狗、猫和所有的非人灵长类[3, 4]。在这些动物中Aβ的序列与人类完全一致,但目前为止,尚未在大、小鼠中发现类似的SP存在,这很可能是因为大、小鼠Aβ在序列与人有3个氨基酸序列的差异导致Aβ无法聚集形成SP。
树鼩属于介于灵长目与食虫目之间的独立阶元—攀鼩目树鼩科,与啮齿类相比,树鼩更接近非人灵长类和人类。现在世界上许多国家已经开展了树鼩实验动物方面的研究,并取得了丰硕的成果[5]。树鼩是公认的近视模型动物[6],又因其大脑发达,多用于神经系统的研究及神经系统疾病模型的复制,如脑血管病[7]、抑郁症[8]、老年痴呆[9]等,2012年,Yamashita 等[10]的报道支持了猕猴和树鼩均有这种人类老年痴呆症病理样的老年斑。从基因序列上来说,Pawlik等[4]早在1999年便对脑中的APP基因序列进行了测定,获得了APP一个可变剪接体的序列信息,结果发现和人的同源性大于大小鼠,并且Aβ序列也和人类的完全一致。综上,树鼩有可能是创建人类老年痴呆症模型的理想动物。
通过鸟枪测序法获得的树鼩基因组序列中,对树鼩的APP基因及可变剪接体进行了预测,依据测序结果与人相应序列的比对,预测得到树鼩可能存在6种剪接体[11],不排除这些潜在的剪接体有些可能实际上并不存在以及有其他剪接体存在的可能。鉴于APP基因在AD中的重要作用及树鼩在阿尔兹海默症模型的应用,弄清树鼩APP基因的所有剪接体的类型及在不同组织的表达差异、表达模式,对今后阿尔兹海默症树鼩动物模型创制和疾病发生机理研究具有重要意义。
1 材料与方法
1.1 实验材料及分析软件
滇西亚种雄性树鼩(Tupaiabelangeri),由中国医学科学院医学生物学研究所树鼩种质资源中心【SCXK(滇)K2013-0001】提供。TRIzol试剂、逆转录试剂盒、Primer-Star DNA聚合酶、DNA Marker DL 2000购自TaKaRa Biotechnology公司(大连,中国);胶回收试剂盒购自Tiangen公司(北京,中国)。利用生物信息学软件DNAman进行比对分析,利用生物信息学软件Mega7绘制系统进化树。利用Swiss Model工作站进行三维建模,并用Pymol显示。
1.2 分子生物学方法
1.2.1 引物设计
通过在PubMed上查询已知人的(基因ID:351)和预测的树鼩APP基因序列(基因ID:102489280,KF479228.2),根据此设计出共同的特异性引物。利用电泳区分出不同长度的剪接体,胶回收后测序确定其分型。
1.2.2 总RNA的提取
树鼩组织RNA的提取和定量。精确称量100 mg树鼩各组织,加入1 mL预冷的TRIzol提取液,使用超声粉碎的方法将组织充分裂解,其余步骤按TRIzol试剂说明书进行。所提取的RNA 溶解于DEPC处理水中。采用紫外分光光度法定量。
1.2.3 RNA反转录
为cDNA利用RT-PCR方法扩增APP cDNA,逆转录反应按试剂盒中的说明书进行。
1.2.4 PCR扩增
以Oligo(dT)为引物,逆转录合成cDNA,之后以此为模板,进行PCR扩增。在反应体系中先将模板于95℃变性5 min,循环参数:94℃30 s,55℃45 s,72℃ 1 min,35 个循环,72℃延伸10 min。APP上游:GTGCCCACTGACGGCAAT,APP下游:GCTCCTCCAGGGATGTATTTATT。
1.2.5 PCR的产物回收
扩增产物的胶回收用1.0% 琼脂糖凝胶电泳,紫外灯下检查PCR 产物。之后将PCR 条带用手术刀小心割取,按照胶回收试剂盒说明书分步回收PCR 产物。
1.2.6 PCR测序
产物测序将电泳后回收的PCR产物送上海生工公司测序,检测PCR产物的正确性。
1.2.7 总APP剪接体定量PCR
反应混合液为2× qPCR预混液10 μL,上下游引物各0.4 μL,ddH2O 7.2 μL,模板2 μL。引物见表1。PCR循环条件为保持3 min 95℃(95℃、7 s, 57℃、10 s,72℃、15 s)45个循环。APP定量上游:GAAGCCAACCAACCAGTGAC,APP定量下游:CTAAGCAGCGGTAGGGAATG,GDPH定量上游:CCATCACCATCTTCCAGGAGCGAG,GDPH定量下游:CAAAGGTGGAGGAGTGGGTGTCG。
2 结果
2.1 APP770核酸序列和推导的氨基酸序列
树鼩的APP基因由18个外显子构成。测序结果表明树鼩APP剪接体的全长为3514 bp,有一个109 bp的5’-UTR,1092 bp的3’-UTP。三个潜在的AATAAA聚腺苷酸化信号位于多聚Aβ尾巴上游处。在推测的树鼩APP基因氨基酸产物中,在N端携带有一个19氨基酸残基的信号肽(图1)。

图1 树鼩APP基因的核酸序列和推导的氨基酸序列Fig.1 Nucleotide and deduced amino acid sequence of APP gene in the Tupaia chinensis
2.2 APP基因的进化分析
为评价APP蛋白的进化保守性,与其他物种(人、食蟹猴、大鼠、小鼠、猪、狗、黑猩猩、兔)的氨基酸序列的比对分析。显示,APP基因在调查的9个物种中高度同源保守,有一个17个氨基酸的信号肽序列,同源性很高。树鼩与人的APP氨基酸序列有97.3%的一致性,仅有20个氨基酸序列不同,并且大部分差异氨基酸位于蛋白末端序列(图2)。树鼩与人(Homosapiens)、食蟹猴(Macacafascicularis)、大鼠(Rattusnorvegicus)、小鼠(Musmusculus)、猪(Susscrofa)、狗(Canislupus)、黑猩猩(Pantroglodytes)、兔(Oryctolaguscuniculus)的同源相似性分别为97.3%、97.3%、97.4%、97.1%、97.9%、97.4%、97.4%、97.1%。
在基于核酸序列和氨基酸序列构建的NJ进化树中,观察到一个与基于已知序列和基因组序列获得的进化树相同的聚类模式,树鼩与灵长类显示出一个较近的亲缘关系(图3),获得的进化结果稳定可靠。

图2 树鼩与其他亲缘较近的8种脊椎动物APP氨基酸序列的比对结果Fig.2 Alignment of the APP amino acid sequences in 9 vertebrate species

图3 分别基于氨基酸序列(A)、核酸序列(B)的进化树Fig.3 Phylogenetic trees of the IL7 gene based on amino acid sequences (A) and deduced nucleotide sequences (B)
2.3 树鼩APP蛋白与人APP蛋白三维结构比对
通过三维建模,树鼩和人的APP基因分别有4个结构域:半胱氨酸丰富的球状区E1;KPI区;α螺旋丰富的E2区,包含了一部分的Aβ,延伸至跨膜区;跨膜区和C末端信号转导区。12个差异氨基酸位于无规则卷曲中,8个差异氨基酸位于E1区和KPI区中,而E2区和跨膜信号区高度同源保守,没有差异氨基酸,可能和维持功能相关(图4)。
2.4 APP基因mRNA各种剪接体的序列比对
为确认树鼩中所有的APP基因的可变剪接体,我们对从不同组织中获得的总RNA中的大量cDNA克隆进行测序,除了以上提到的完整的典型的APP770剪接体,我们还确认了其他3种不同的APP基因可变剪接体(图5,基因ID号:KY399768~KY399771)。通过序列间的比对确认了这4种通过外显子跳跃产生的APP可变剪接体,包括APP770,APP695,APP751,APP677四种。
2.5 APP mRNA和各可变剪接体的表达模式
定量PCR的方法确认了APP所有剪接体在10个树鼩不同组织中的表达情况,4种剪接体均表达于肺、肾和肠,APP695和APP677在心脏、肝、脾、肌肉、肾上腺中共表达,APP751和APP695在大脑、小脑、海马和睾丸中共表达,睾丸中还表达APP677,脊髓中单独表达APP695,脂肪中单独表达APP770,APP770主要在肌肉中分布(图6)。

图4 树鼩(A)和人(B)的APP蛋白三维建模Fig.4 Structure of APP in the tree shrew (A) and humans (B)

图5 树鼩APP基因mRNA及其可变剪接体的示意图Fig.5 Schematic structure of APP mRNA and its transcripts in Chinese tree shrew
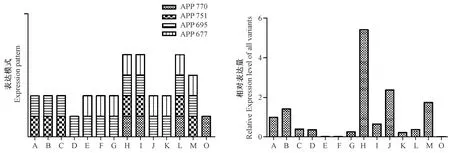
注:A:大脑;B:小脑;C:海马;D:脊髓;E:心脏;F:肝;G:脾;H:肺;I:肾;J:肌肉;K:肾上腺;L:肠;M:睾丸;N:脂肪。图6 树鼩APP基因在不同组织的分布(左)及相对表达量(右)Note. A. Brain;B. Cerebellum;C. Hippocampus;D. Spinal cord;E. Heart;F. Liver;G. Spleen;H. Lung;I. Kidney;J. Muscle;K. Adrenal gland;L. Intestine;M. Testis;N. Adipose tissue.Fig.6 Distribution patterns of APP(left)and its transcripts expression levels in different tissues(right)
3 讨论
APP具有促进细胞增生和神经营养因子的作用,在细胞膜上是一种跨膜糖蛋白,通过与细胞膜外基质的相互作用介导神经细胞之间的粘附,增加神经突触之间的联系及突触可塑性。APP可能通过作用于细胞表面的特异受体来刺激轴突的发生和生长,APP通过蛋白酶剪切后产生的Aβ,长久以来一直被认为是导致AD产生的最根本原因。因此,影响APP剪切的重要因素(包括APP基因的结构、基因的序列和突变、可变剪接体的存在形式和分布)一直是研究的热点[12, 13],目前,人[14]、大鼠[15]、小鼠[16]等物种中的APP基因及所有的可变剪接体都已经有报道,而被认为是理想痴呆模型动物的树鼩的APP基因仅有其中的一种剪接体APP695有报道,以及鸟枪法测序预测获得的六种可变剪接体,这些信息还远远不够。
因此,本研究旨在区分并鉴定树鼩APP mRNA的所有可变剪接体,在对这一与AD疾病发生和进展密切相关的关键分子的基因进行特征描述的基础上,再结合可变剪接体的分布情况进一步调查APP基因在不同组织中的表达量。本研究首次获得了APP最长的可变剪接体APP770的分子序列,并通过blast获得的同源性最高的9种物种中进行序列比对分析,结果表明,树鼩特异的氨基酸有5个,分别是第12位的V、 193位的N、 254位的A、 301位的Q、 354位的W。有趣的是,N末端的序列同源性较低,而C末端的长达143氨基酸的序列以及A基的序列与人的完全一致。
在基于核酸序列和氨基酸序列构建的NJ进化树中,发现一个与基于已知序列和基因组序列获得的进化树相同的聚类模式,树鼩与猪和狗更为接近,在进化上处于同一分支,而人、黑猩猩和猴最为接近,在进化上处于同一分支,大小鼠和兔最为接近,在进化上处于同一分支,树鼩与灵长类显示出一个较近的亲缘关系(图3),获得的进化结果稳定可靠。
通过三维建模,树鼩和人的APP基因三维结构非常类似,分别有4个结构域:半胱氨酸丰富的球状区E1;KPI区;α螺旋丰富的E2区,包含了一部分的Aβ,延伸至跨膜区;跨膜区和C末端信号转导区。共20个的差异氨基酸中,12个差异氨基酸位于无规则卷曲中,8个差异氨基酸位于E1区和和KPI区中,而E2区和跨膜信号区高度同源保守,没有差异氨基酸,可能和维持功能相关,这个区域中有与AD发病最为相关的Aβ分子编码区,可以看到,此次测序的结果与之前的一样,树鼩的Aβ序列与人的完全一致,而大小鼠存在3个氨基酸的差异,这可能与老年树鼩脑中可以形成SP有关。
为确认树鼩中所有的APP基因的可变剪接体,对从不同组织中获得的总RNA中的大量cDNA克隆进行测序,除了以上提到的完整的典型的APP770剪接体,还确认了其他3种不同的APP基因可变剪接体(图4,基因ID号:KY399768~KY399771)。同时确认了这4种通过外显子跳跃产生的APP可变剪接体在不同组织中的分布(图5),包括APP770、APP695、APP751、APP677四种。在之前公布的树鼩基因组序列中,预测获得了6种可变剪接体,排除了由于技术限制未能测到的35个氨基酸的N末端和一个544位点的FFF序列检测为SLSL而多一个氨基酸之外,预测得到的其实为:所有外显子(APP770),缺外显子8(APP751),7和8(APP695),8和15(APP733),7、8和15(APP677),15(APP752)六种类似人的可变剪接体,经过实验检测,仅验证了四种的存在,没有APP733和APP752的存在。
APP剪接体的分布有组织特异性,4种剪接体均表达的是肺、肾和肠,APP695和APP677在心脏、肝、脾、肌肉、肾上腺中共表达,APP751和APP695在大脑、小脑、海马和睾丸中共表达,睾丸中还表达APP677,脊髓中单独表达APP695,脂肪中单独表达APP770。选择性剪切得到的可变剪接体通常具有组织特异性,并在特定组织和发育阶段起着关键作用,本研究结果和之前的结果类似,APP695主要在脑中分布,但肌肉中未检测到APP770的存在可能是受到了技术的限制。其表现的共表达方式也很可能与功能相关。同时,通过定量PCR确定表达量最高的是肺、肌肉和睾丸,大脑、小脑和脊髓等神经组织中表达量中等。
总之,本研究首次确认了树鼩APP基因的可变剪接体的分型和表达模式,提示树鼩有可能是作为神经系统疾病的理想动物。
[1] Selkoe DJ and Hardy J.The amyloid hypothesis of Alzheimer’s disease at 25 years[J]. EMBO MolMed, 2016,8(6): 595-608.
[2] Kirkitadze MD, Bitan G, Teplow DB, et al. Paradigm shifts in Alzheimer’s disease and other neurodegenerative disorders: the emerging role of oligomeric assemblies[J]. J Neurosci Res, 2002,69(5): 567-577.
[3] Head E.A canine model of human aging and Alzheimer’s disease[J]. Biochim Biophys Acta, 2013.1832(9): 1384-1389.
[4] Pawlik M, Fuchs E, Walker LC,et al. Primate-like amyloid-beta sequence but no cerebral amyloidosis in aged tree shrews[J]. Neurobiol Aging, 1999,20(1): 47-51.
[5] 徐林, 张云, 梁斌, 等. 实验动物树鼩和人类疾病的树鼩模型研究概述[J]. 动物学研究, 2013,34(02): 59-69.
[6] 周广龙, 朱勤, 李振宇, 等. 树鼩在眼科学的基础研究进展[J]. 中国实验动物学报, 2015, 23(06): 652-655.
[7] 李树清, 李凡, 何亮,等. 缺血后适应促进树鼩血栓性脑缺血时紧密连接occludin/ZO-1蛋白表达及抑制脑水肿的机制[J]. 中国病理生理杂志, 2016,32(3) :477-484.
[8] Schmelting B, Corbach-Sohle S, Kohlhause S, et al. Agomelatine in the tree shrew model of depression: effects on stress-induced nocturnal hyperthermia and hormonal status[J]. Eur Neuropsychopharmacol, 2014,24(3): 437-447.
[9] Lin N, Xiong LL, Zhang RP, et al. Injection of Abeta1-40 into hippocampus induced cognitive lesion associated with neuronal apoptosis and multiple gene expressions in the tree shrew[J]. Apoptosis, 2016,21(5): 621-640.
[10] Yamashita A, Fuchs E, Taira M, et al. Somatostatin-immunoreactive senile plaque-like structures in the frontal cortex and nucleus accumbens of aged tree shrews and Japanese macaques[J]. J Med Primatol, 2012,41(3): 147-157.
[11] Fan Y, Yu D, Yao YG. Tree shrew database (TreeshrewDB): a genomic knowledge base for the Chinese tree shrew[J]. Sci Rep, 2014,4: 7145.
[12] Nicolas G, Charbonnier C, Campion D. From common to rare variants: the genetic component of Alzheimer disease[J]. Hum Hered, 2016, 81(3): 129-141.
[13] Tcw J, Goate AM. Genetics of beta-amyloid precursor protein in Alzheimer’s disease[J]. Cold Spring Harb Perspect Med, 2016, a024539.
[14] Cuccaro ML, Carney RM, Zhang Y, et al. SORL1 mutations in early- and late-onset Alzheimer disease[J]. Neurol Genet, 2016,2(6): e116.
[15] Sandbrink R, Masters CL, Beyreuther K. Beta A4-amyloid protein precursor mRNA isoforms without exon 15 are ubiquitously expressed in rat tissues including brain, but not in neurons[J]. J Biol Chem, 1994,269(2): 1510-1517.
[16] Izumi R, Yamada T, Yoshikai S,et al. Positive and negative regulatory elements for the expression of the Alzheimer’s disease amyloid precursor-encoding gene in mouse[J]. Gene, 1992,112(2): 189-195.
Characterization of tree shrew APP gene and identification of its alternatively spliced variants
HAN Yuan-yuan, SUN Xiao-mei, KUANG De-xuan, LU Cai-xia, CHEN Ling-xiaTONG Pin-fen, WANG Wen-guang, LI Na, DAI Jie-jie*
(Center of Tree Shrew Germplasm Resources, Institute of Medical Biology, Chinese Academy of Medical Sciences and Peking Union Medical College. Yunnan Key Laboratory of Vaccine Research and Development on Severe Infectious Diseases.Yunnan Innovation Team of Standardization and Application Research on Tree Shrew.Kunming 650118, China )
Objective The aim of this study was to differentiate and identify all the variants of APP mRNA in tree shrew, describe the characteristics of APP genes, and determine its distribution in different tissues. Method Based on the known human APP sequence and predicted tree shrew APP gene, we designed a pair of common specific primer. We extracted total RNA from different tissues respectively, using RT-PCR to get the targeted cDNA. We differentiated the variants by electrophoresis. Finally we recollected the RT-PCR products for sequencing. Combined with the qPCR results, we confirmed the quantitative distribution of the variants in different tissues. Results Our sequencing results showed that the length of the tree shrew APP spliceosome was 3514 bp with a 109 bp 5′-UTR and a 1092 bp3′-UTP. APP genes in the investigated 9 species were highly homologous and conservative, tree shrews and primates show a very close genetic relationship. The tree shrew APP genes shared four domains by 3D modeling. We also confirmed the distribution pattern of 4 spliceosomes derived from exon jumping. All existed APP variants, including APP770, APP695, APP751, and APP677, were all expressed in the lung, kidney and intestine. The highest expression levels were in the lung, muscle, and testicles. Conclusions This study helps to promote the studies of tree shrews as an Alzheimer’s disease model and the mechanism of its pathogenesis and drug development.
TupaiaBelangeri; APP mRNA variants; APP distribution; Alzheimer’s disease model
DAI Jie-jie, E-mail: djj@imbcams.com.cn
国家科技支撑计划项目 (2014BAI01B00);云南省联合支持国家计划项目(2015GA009);云南省科技人才和平台计划项目。
罕园园(1983年-),女,助理研究员。E-mail: hyy@imbcams.com.cn
代解杰,男,项目负责人。E-mail: djj@imbcams.com.cn
Q95-33
A
1005-4847(2017) 02-0132-07
10.3969/j.issn.1005-4847.2017.02.004
2017-01-04
