中间球海胆幼体及稚海胆生长性状的遗传参数估计
2017-05-17韩奋杰张伟杰秦宇博陈顺周秘陈小慧常亚青
韩奋杰,张伟杰,秦宇博,陈顺,周秘,陈小慧,常亚青
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023;2.辽宁省海洋环境预报与防灾减灾中心,辽宁沈阳110001)
中间球海胆幼体及稚海胆生长性状的遗传参数估计
韩奋杰1,张伟杰1,秦宇博2,陈顺1,周秘1,陈小慧1,常亚青1
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023;2.辽宁省海洋环境预报与防灾减灾中心,辽宁沈阳110001)
为估计中间球海胆Strongylocentrotus intermedius幼体及稚海胆生长性状的遗传参数,采用巢式不平衡设计方法建立了44个中间球海胆全同胞家系,并测量了各家系海胆四腕幼体、六腕幼体和八腕幼体等各期浮游幼体的体长以及受精后30 d的稚海胆壳径,采用动物模型和约束性最大似然法(REML)估计了中间球海胆幼体和稚海胆生长性状的遗传参数。结果表明:中间球海胆四腕幼体和八腕幼体体长的遗传力分别为0.705±0.373和0.538±0.444,为高度遗传力,六腕幼体体长和稚海胆壳径的遗传力均接近于0,为低度遗传力;各阶段幼体体长和稚海胆壳径的共同环境效应为0.048~0.352;各阶段幼体体长间遗传相关为正相关(0.300~0.518),各阶段幼体体长与稚海胆壳径间的遗传相关为负相关(-0.152~-0.435)。本研究结果可为制定中间球海胆的早期选择和间接选择选育方案提供参考依据。
中间球海胆;幼体;稚海胆;生长性状;遗传参数
中间球海胆Strongylocentrotus intermedius又称虾夷马粪海胆,隶属于棘皮动物门、海胆纲、正形目、球海胆科[1-2],原产于日本北方和俄罗斯远东等地沿海,其性腺色泽好﹑味甜,在国际市场上很受欢迎,是海胆类中经济价值最高的种类之一。大连海洋大学于1989年从日本将中间球海胆引入中国,随后掌握了其人工育苗技术[3-4],并在筏式和工厂化养殖技术以及环境因子等方面进行了研究[5-8]。目前,中间球海胆已成为中国主要的海胆养殖种类之一,国内每年的苗种生产量已达亿枚以上。为了提高中间球海胆的产量和品质,避免近亲交配等原因带来的种质退化,利用选择育种等方法对其生长和品质性状进行遗传改良十分必要。
经济性状的遗传力和遗传相关等遗传参数估计是水产动物选择育种的一项基础工作,遗传力和遗传相关在育种值估计、选择指数确定、选择反应预测、间接选择、早期选择、选择方法比较及育种规划决策等方面起着十分重要的作用[9-10]。关于水产动物经济性状的遗传参数估计有较多的报道。Aulstad等[11]通过同胞相关法估计了虹鳟Salmo gairdneri 150 d体质量的遗传力;Hetzel等[12]估计了日本对虾Penaeus japonicus生长性状的遗传力; Kong等[13]利用45个全同胞家系和动物模型方法估计了12月龄长牡蛎Crassostrea gigas壳长的遗传力;孟思远等[14]估计了刺参Apostichopus japonicus体长、体质量、肉刺数目和肉刺长度等4个性状的遗传力。
对中间球海胆的选择育种及其家系选育研究已有报道,但对中间球海胆遗传参数的研究多集中于幼海胆和成海胆。如刘小林等[15]最早对中间球海胆生长性状的遗传力进行了估计,认为海胆3~5月龄体质量的狭义遗传力为0.339~0.523,壳径的狭义遗传力为0.316~0.487。Liu等[16]进一步估计了中间球海胆8~12月龄壳高、壳径和体质量的遗传力和遗传相关。Chang等[17]估计了中间球海胆6~18月龄生长性状的遗传力,以及收获季节海胆生长性状和生殖腺性状的遗传力和性状间的遗传相关,认为中间球海胆生长性状具有中高度遗传力,多数生殖腺性状和生长性状间存在显著的遗传相关。然而,关于中间球海胆幼体和稚海胆生长性状的遗传参数目前尚未见报道。由于早期生长性状的遗传参数可为开展早期选择提供依据,因此,中间球海胆幼体及稚海胆生长性状的遗传参数亟待研究。
本研究中利用巢式不平衡设计建立了中间球海胆的全同胞家系,用动物模型和约束性最大似然法(REML)估计了中间球海胆浮游幼体和稚海胆两个阶段生长性状的遗传力和性状在不同时期的遗传相关,以期为中间球海胆选择育种提供必要的基础参数,并为早期选择育种方案的制定提供参考依据。
1 材料与方法
1.1 材料
本试验中家系构建所用亲本为大连海洋大学农业部北方海水增养殖重点实验室连续5代选育群体,其中雄性亲本26枚,雌性亲本44枚。家系构建前,对亲本进行促熟,水温为16~18℃,投喂海带和煮熟的贻贝肉,每2 d全量换水一次,连续充气。亲胆产卵前2 h将其运到育苗车间准备产卵。
1.2 方法
1.2.1 家系构建与培育 采用催产剂(0.5 mol/L的KCl溶液)进行催产,分别收集每个亲胆的精液或卵液,严防混合,按照雄雌比例为1∶(1~2)的巢式设计和避免近亲交配的原则构建家系,共建立26个半同胞家系,包含44个全同胞家系。
受精完成后,将各家系受精卵分别置于独立的10 L孵化桶内,经2次洗卵去除多余的精子后进行孵化。孵化水温为18.0~19.0℃,孵化密度为50 ind./mL,光照度低于300 lx。受精后30 h左右至棱柱幼体,此时利用260目筛绢做成的漏网捞取水面幼体至500 L培育池内,培育密度调整为0.8 ind./mL,水温与光照条件不变,微弱充气使幼体在水中分布均匀。投喂饵料为牟氏角毛藻Chaetoceros muelleri,每天4次,投饵前镜检幼体摄食消化情况,并以此确定投喂量,每天换水1次,每次为全量的1/2,每隔4 d倒池1次。在此条件下培育约20 d,幼体发育经二腕幼体、四腕幼体、六腕幼体和八腕幼体期,八腕幼体后期往培育槽内投放附有底栖硅藻的波纹板,使幼体附着变态为稚海胆,变态后提升光照度为3000 lx,适量增加充气量,停止投饵,其他培育条件不变。
1.2.2 生长性状测量 分别于受精后5 d(四腕幼体)、10 d(六腕幼体)和15 d(八腕幼体)时,对26个半同胞家系(含44个全同胞家系)内中间球海胆的幼体进行随机取样,每个家系取30只幼体至烧杯中,滴入1 mL福尔马林溶液将幼体固定,待幼体沉至杯底后,逐个吸取幼体至显微镜下,用目微尺测量幼体体长。在受精后30 d,浮游幼体变态为稚海胆后,对24个半同胞家系(含40个全同胞家系)进行取样,从每个家系取30只稚海胆,用目微尺在显微镜下测量其壳径。
1.2.3 遗传参数估计 采用多性状个体动物模型估计四腕幼体、六腕幼体和八腕幼体体长以及稚海胆壳径的遗传参数,育种分析模型为

其中:yij表示第j个全同胞下第i只海胆各性状的表型值;u表示各性状的平均值;ai表示各性状的加性遗传效应;cj表示第j个全同胞的共同环境效应;eij表示性状的随机残差。利用ASReml软件估计各性状的方差组分,并根据以下公式计算遗传参数:
遗传力(h2)计算公式为

共同环境效应(c2)计算公式为
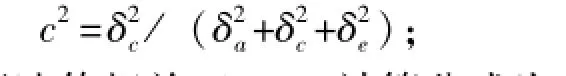
两性状间遗传相关(rgxy)计算公式为
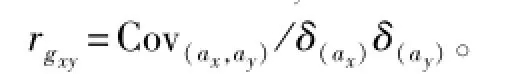
2 结果与分析
2.1 中间球海胆幼体及稚海胆生长性状的表型统计量
从表1可知:中间球海胆第5~15天浮游幼体体长为519.81~813.61 μm,第30天稚海胆壳径平均值为684.29 μm;浮游幼体体长的变异系数为10.93%~14.18%,且随着幼体体长的增长而逐渐增大,至稚海胆时,变异系数增加至31.90%。
2.2 中间球海胆幼体及稚海胆生长性状的方差组分估计
从表2可知:各方差组分中,遗传方差组分较不稳定,四腕幼体和八腕幼体的遗传方差组分较大,而六腕幼体和稚海胆的遗传方差组分却极小;共同环境方差组分和随机残差方差组分均随着幼体的生长而逐渐增大。
2.3 中间球海胆幼体及稚海胆生长性状的遗传力估计
从表3可知:四腕幼体和八腕幼体体长的遗传力较高,分别为0.705±0.373和0.538±0.444,而六腕幼体体长和稚海胆壳径的遗传力均接近于0;各阶段幼体的共同环境效应为0.048~0.352,以六腕幼体时期最高,四腕幼体时期最低。
2.4 中间球海胆幼体及稚海胆生长性状间的遗传相关
从表4可知:中间球海胆在第5天、10天和15天时期内的遗传相关为正相关,为0.300~0.518,而第30天的稚海胆壳径与各期浮游幼体的体长的遗传相关为负相关,为-0.152~-0.435。

表1 中间球海胆幼体及稚海胆的表型统计量Tab.1 Phenotype statistics of larvae and juveniles of sea urchinStrongylocentrotus intermedius

表2 中间球海胆幼体及稚海胆生长性状的方差组分估计Tab.2 Variance component estimates for growth traits of larvae and juveniles of sea urchinStrongylocentrotus intermedius

表3 中间球海胆幼体及稚海胆生长性状的遗传力和共同环境效应估计Tab.3 Heritabilities and common environmental effect for growth traits of larvae and juveniles of sea urchinStrongylocentrotus intermedius
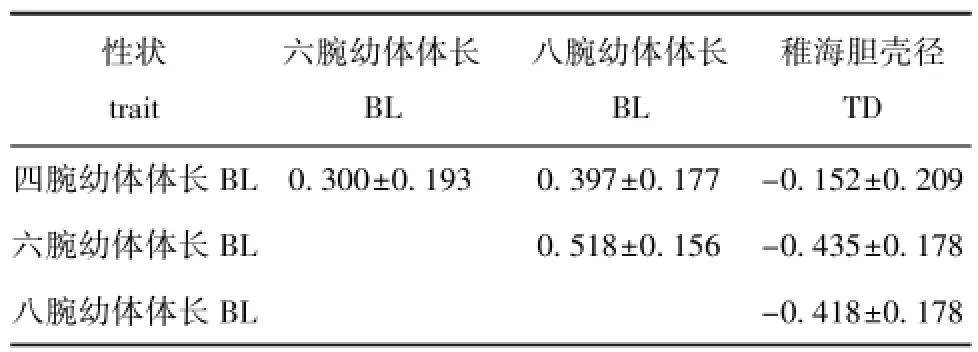
表4 中间球海胆幼体及稚海胆生长性状的遗传相关Tab.4 Genetic correlations for growth traits of larvae and juveniles of sea urchinStrongylocentrotus intermedius
3 讨论
3.1 中间球海胆幼体及稚海胆的表型变异
变异是选择育种的基础,评估一个物种的某个性状是否具有选择潜力,往往根据该性状的表型变异作初步判断。本研究中,中间球海胆5~15 d浮游幼体(分别为四腕幼体、六腕幼体和八腕幼体)体长的表型变异系数为10.93%~14.18%,高于王庆志等[19]、陶后全等[20]报道的长牡蛎、马氏珠母贝幼体壳长和壳高的变异系数(分别为4.45%~12.41%、4.4%~10.22%),而与栾生等[21]、李云峰等[22]报道的刺参浮游幼体体长的表型变异系数相近(分别为9.58%~10.66%、14.6%)。随着个体生长,幼体及稚海胆生长性状的变异系数逐渐增大,这与王庆志等[19]、陶后全等[20]、李云峰等[22]、郭华阳等[23]的研究结果相似,这一规律在水产动物中较为普遍。其原因一方面可能是随着个体的生长,与个体生长相关的基因差异逐渐显现,使个体表型差异增大;另一方面可能是由于水产动物在幼体培育过程中不同家系所处环境难以保持完全一致,环境变异的产生增加了个体生长性状的表型变异。因环境变异组分不能遗传给后代,需将其从表型变异中剖分出来获得遗传变异,再必须进行遗传力估计才能达到准确判断性状选育潜力的目的。
3.2 中间球海胆幼体及稚海胆生长性状的遗传力
中间球海胆不同阶段生长性状的遗传力估计值变化较大,四腕幼体和八腕幼体体长的遗传力均为高度遗传力(分别为0.705和0.538),而六腕幼体体长和稚海胆期壳径的遗传力均接近于0,为低度遗传力。共同环境效应较高可能是六腕幼体遗传力低的主要原因。共同环境效应估计值与遗传力估计值具有一定的负相关性,六腕幼体体长和稚海胆期壳径的共同环境效应均较高,而四腕幼体和八腕幼体体长的共同环境效应则较低,这表明共同环境效应对遗传力产生了一定的影响。李冬春等[24]对菲律宾蛤仔壳长的遗传力估计得到了同样的结果,即在90 d和330 d时,壳长遗传力估计值为0.26~0.36,而300 d时壳长的遗传力估计值仅为0.02,共同环境效应估计值与遗传力估计值同样具有负相关性。共同环境效应一般包括母体效应,以及家系分养时由于家系间环境不同和家系内环境相同造成的环境效应[9]。本研究中,四腕幼体时期的共同环境效应仅为0.048,说明家系间环境差异和母体效应极低,由于母体效应具有随后代生长而降低的一般规律,因此,六腕幼体至稚海胆时期共同环境效应的增加,应为家系间培育环境的不一致造成的。中间球海胆的浮游幼体需经过多次变态,不同家系的变态率差异会导致变态后的培育密度在家系间存在一定的差异,这一培育环境的差异是难以避免的,由其造成的较高共同环境效应降低了六腕幼体时期的遗传力。由于REML法能剖分加性效应和共同环境效应,因此,使用REML法会比用传统的同胞组内相关法在遗传力的估计上更为准确[25]。例如,栾生等[21]利用全同胞组内相关法估计刺参中耳幼体体长的遗传力为0.75,而李云峰等[22]利用REML方法估计仅为0.29,共同环境效应为0.26。本研究中估计的中间球海胆四腕幼体和八腕幼体体长的遗传力(分别为0.538、0.705)高于刘小林等[15]、Liu等[16]、Chang等[17]估计的中间球海胆生长性状的遗传力,表明这两个时期幼体体长具有极大的选育潜力,在此时进行选择应会收到较好的效果,而在六腕幼体和稚海胆时期,体长的遗传力极低,应避免进行选择。
3.3 中间球海胆幼体及稚海胆生长性状的遗传相关
对中间球海胆生长性状的遗传相关研究主要集中于同一时期的不同性状间[16-17],本研究中对中间球海胆不同时期生长性状间的遗传相关进行了估计。遗传相关是间接选择理论中重要的遗传参数,同一性状在不同时期是否具有遗传相关是判断早期选择可行性的重要依据[9]。中间球海胆四腕幼体、六腕幼体和八腕幼体体长的遗传相关为正相关。Hilbish等[26]研究表明,硬壳蛤胚壳Ⅰ期和第2天的壳长间具有较高的遗传正相关。梁冰冰等[27]研究表明,文蛤附着变态期及稚贝期的壳长具有正向的遗传相关。本研究结果与以上研究结果类似,说明浮游幼体期间各期幼体体长相关的基因可能较相似,或基因间存在一定的连锁关系。第30天的稚海胆壳径与各期浮游幼体的体长间的遗传相关为负相关,这说明控制稚海胆壳径的基因可能与之前控制幼体体长的基因不同,且具有反向作用。这一结果可能与海胆的生物学特征有一定关系,海胆在附着变态时会将占体长大部分比例的腕脱去,原来的前庭复合体、肠胃和外壳构成稚海胆的身体[1],此时的壳径与原幼体的体长可能由相反作用的基因控制,而导致了遗传相关为负值。遗传相关估计的结果表明,中间球海胆幼体期间进行早期选择具有一定的可行性,即通过对早期浮游幼体进行正向选择,可实现对晚期幼体的正向选择和稚海胆的负向选择。
[1] 常亚青,丁君,宋坚,等.海参、海胆生物学研究与养殖[M].北京:海洋出版社,2004.
[2] 亓守冰,张伟杰,常亚青,等.中间球海胆(♀)与光棘球海胆(♂)种间杂交及自繁后代生长与表型特征比较[J].大连海洋大学学报,2015,31(5):461-466.
[3] 王子臣,常亚青.虾夷马粪海胆人工育苗的研究[J].中国水产科学,1997,4(1):60-67.
[4] 王子臣,常亚青.经济类海胆增养殖研究进展及前景[J].海洋科学,1997,21(6):20-22.
[5] 常亚青,王子臣.虾夷马粪海胆筏式人工养殖研究[J].大连水产学院学报,1997,12(2):7-14.
[6] 常亚青,王子臣,孙丕海,等.虾夷马粪海胆的海区渡夏、室内中间培育及工厂化养成[J].中国水产科学,1999,6(2):66-69.
[7] 常亚青,王子臣.低盐度海水和饵料对虾夷马粪海胆的影响[J].海洋科学,1997,21(3):1-2.
[8] 常亚青,王子臣,王国江.温度和藻类饵料对虾夷马粪海胆摄食及生长的影响[J].水产学报,1999,23(1):69-76.
[9] 盛志廉,陈瑶生.数量遗传学[M].北京:科学出版社,1999.
[10] Gjedrem T.Genetic improvement of cold-water fish species[J]. Aquaculture Research,2000,31(1):25-33.
[11] Aulstad D,Gjedrem T,Skjervold H.Genetic and environmental sources of variation in length and weight of rainbow trout(Salmo gairdneri)[J].Journal of the Fisheries Research Board of Canada,1972,29(3):237-241.
[12] Hetzel D J S,Crocos P J,Davis G P,et al.Response to selection and heritability for growth in the kuruma prawn,Penaeus japonicus [J].Aquaculture,2000,181(3-4):215-223.
[13] Kong Ning,Li Qi,Yu Hong,et al.Heritability estimates for growth-related traits in the Pacific oyster(Crassostrea gigas)using a molecular pedigree[J].Aquaculture Research,2015,46 (2):499-508.
[14] 孟思远,常亚青,李文东,等.仿刺参幼参阶段4个生长性状遗传力的估计[J].大连海洋大学学报,2010,25(6):475-479.
[15] 刘小林,常亚青,相建海,等.虾夷马粪海胆早期生长发育的遗传力估计[J].中国水产科学,2003,10(3):206-211.
[16] Liu Xiaolin,Chang Yaqing,Xiang Jianhai,et al.Estimates of genetic parameters for growth traits of the sea urchin,Strongylocentrotus intermedius[J].Aquaculture,2005,243(1-4):27-32.
[17] Chang Yaqing,Zhang Weijie,Zhao Chong,et al.Estimates of heritabilities and genetic correlations for growth and gonad traits in the sea urchin Strongylocentrotus intermedius[J].Aquaculture Research,2012,43(2):271-280.
[18] 林元震.R与ASReml-R统计分析教程[M].北京:中国林业出版社,2014.
[19] 王庆志,李琪,刘士凯,等.长牡蛎幼体生长性状的遗传力及其相关性分析[J].中国水产科学,2009,16(5):736-743.
[20] 陶后全,吴灶和,白成,等.马氏珠母贝早期生长性状的遗传参数估计[J].广东海洋大学学报,2014,34(3):10-16.
[21] 栾生,孙慧玲,孔杰.刺参耳状幼体体长遗传力的估计[J].中国水产科学,2006,13(3):378-383.
[22] 李云峰,常亚青,田燚,等.仿刺参耳状幼体和稚参阶段的体长遗传力估计[J].大连水产学院学报,2009,24(1):30-33.
[23] 郭华阳,张殿昌,李恒德,等.合浦珠母贝幼体生长性状的遗传力及其相关性分析[J].湖北农业科学,2011,50(21): 4441-4444.
[24] 李冬春,闫喜武,霍忠明,等.蛤仔(Ruditapes philippinarum)养成期壳长遗传力与育种值估计[J].海洋与湖沼,2013,44 (6):1550-1556.
[25] 刘增廷,张沅,解春亭.方差组分估计方法的比较[J].畜牧兽医学报,1989,20(4):289-294.
[26] Hilbish T J,Winn E P,Rawson P D.Genetic variation and covariation during larval and juvenile growth in Mercenaria mercenaria [J].Marine Biology,1993,115(1):97-104.
[27] 梁冰冰,王鸿霞.文蛤早期发育阶段生长性状遗传参数的估计[J].海洋科学,2016,40(3):47-52.
Estimates of genetic parameters for growth traits in larval and juvenile sea urchin Strongylocentrotus intermedius
HAN Fen-jie1,ZHANG Wei-jie1,QIN Yubo2,CHEN Shun1, ZHOU Mi1,CHEN Xiao-hui1,CHANG Ya-qing1
(1.Key Laboratory of Mariculture&Stock Enhancement in North China's Sea,Ministry of Agriculture,Dalian Ocean University,Dalian 116023,China; 2.Liaoning Province Marine Environmental Forecasting Center,Shenyang 110001,China)
The body length of four-armed larva,six-armed larva,eight-armed larva and shell diameter of juvenile sea urchin Strongylocentrotus intermedius(30 days post fertilization)were measured in forty-four full-sib families by an unbalanced nest design and genetic parameters for growth traits were estimated using animal model technology and restricted maximum likelihood method(REML).The results showed that heritability of body length was 0.705±0.373 in four-armed larvae and 0.538±0.444 in eight-armed larvae,higher than heritability(nearly zero)in six-armed larvae and diameter in the juveniles.Common environmental effects at each larval stage and juvenile stage were ranged from 0.048 to 0.352,and genetic correlations of body length of different stage larvae were positive and ranged from 0.300 to 0.518.While all genetic correlations of larval body length were negative at each stage and shell diameter of juveniles ranging from-0.152 to-0.435.The findings will provide guidance to the indirect selection and pre-selection in selective breeding of sea urchin.
Strongylocentrotus intermedius;larva;juvenile sea urchin;growth trait;genetic parameter
10.16535/j.cnki.dlhyxb.2017.02.004
2095-1388(2017)02-0145-05
S968.9
:A
2016-06-20
农业部农业科研杰出人才及创新团队项目;辽宁省农业攻关及成果产业化项目(2015203003)
韩奋杰(1995—),男,硕士研究生。E-mail:fenjiehan@163.com
常亚青(1967—),男,博士,教授。E-mail:yqchang@dlou.edu.cn
