浴仙湖沉积物记录的云南极端干旱事件与生态响应评价
2017-05-16陈光杰黄林培陈小林刘园园卢慧斌陶建霜康文刚
胡 葵 陈光杰 黄林培 陈小林 刘园园 卢慧斌 陶建霜 康文刚
(云南师范大学旅游与地理科学学院, 高原地理过程与环境变化云南省重点实验室, 昆明 650500)
浴仙湖沉积物记录的云南极端干旱事件与生态响应评价
胡 葵 陈光杰 黄林培 陈小林 刘园园 卢慧斌 陶建霜 康文刚
(云南师范大学旅游与地理科学学院, 高原地理过程与环境变化云南省重点实验室, 昆明 650500)
为评价湖库生态环境对极端气候的响应特征, 以经历水位明显降低的云南浴仙湖为研究对象, 通过沉积物多指标分析并结合多变量统计方法, 识别了2009—2013年极端干旱事件驱动湖泊生态环境变化的模式。其中沉积物粒度显示近百年来湖泊水文条件总体稳定, 但在2011年左右粗颗粒物质和中值粒径增大, 同期沉积物元素、同位素记录表明水体营养水平没有明显变化。硅藻群落近百年来以湖泊底栖物种占绝对优势, 约2011年开始优势种属Fragilaria被Nitzschia、Navicula等河流水体中常见的物种所替代, 且硅藻多样性指标总体增加, 而指示水位变化的PCA主轴一解释了硅藻群落结构的主要变化(约55.8%)。总之, 极端干旱事件的发生与持续明显改变了湖泊生物群落结构与多样性特征, 已成为影响该气候敏感区水资源安全与生态环境功能的重要挑战之一。
浴仙湖; 极端干旱; 湖泊沉积物; 硅藻; 生物多样性; 粒度
在全球持续变暖的背景下, 区域极端干旱事件的发生频率与幅度总体增强, 导致大量湖库水位下降、水资源总量降低, 从而威胁区域社会经济发展与生态环境功能[1]。云南地区气候类型多样、地理单元众多、地理环境复杂, 近60年来该地区呈现平均气温总体上升、年平均降水量总体减少的趋势[2]。尤其是2009年秋季到2013年春季, 云南地区出现的极端干旱事件持续时间长、影响范围广。据不完全统计, 截至2010年3月下旬, 云南省受灾人数为2405万, 农作物受旱面积超过3.15×104km2, 农业经济损失高达170×108元[3]。2009年8月至2010年3月, 西南地区80%的植被覆盖受到极端干旱抑制作用的影响[4], 并导致了陆地生态系统植物初级生产量的明显降低[5]。目前国内关于极端干旱的研究主要集中于干旱程度的时空分布[6]与陆地生态系统的影响评价[7]。如张彬等[8]研究表明, 极端干旱事件会造成草原生态系统群落的优势种更替和多样性改变; 同时干旱气候通过植物光合作用可以降低陆地生态系统总初级生产力[9]。
目前围绕云南湖泊的调查研究集中于湖泊富营养化的过程与机制[10—12], 鲜有对湖泊生态系统响应极端干旱气候的模式研究, 主要原因包括缺乏长期、连续的湖泊监测数据。湖泊沉积物记录了湖泊生态与环境变化的长期历史[13], 已被广泛应用于评价湖泊环境变化的时段、幅度和生态响应模式[14]。硅藻是湖泊初级生产者的重要组成, 因其细胞壁由硅质构成可在湖泊沉积物中较好地保存, 加之其属种丰富、生命周期较短(1—7d)且对气候与环境变化十分敏感, 已成为湖泊生态环境评价中重要的生物指标之一[15]。已有沉积物调查表明, 气温上升可以导致湖泊热力分层增强从而有利于小型硅藻的生长[16,17]。对中高纬度地区湖泊沉积物等记录的研究显示, 长期的气候变暖驱动了硅藻等生物群落结构与多样性的区域分布[18]。国内对硅藻响应气候变化的研究主要集中于内陆地区的高盐度湖泊千年时间尺度的变化模式[19]。在长江中下游地区, 气候变暖和水动力变化可以导致硅藻群落的长期变化[20]。在云南地区, 近几十年来的气温变暖导致了泸沽湖硅藻群落的演替[21], 同时现代调查表明, 洱海硅藻群落的构建受到了季节性气候与环境变化的控制[22]。因此, 可以应用湖泊沉积物记录来对云南地区的极端干旱气候事件及其生态环境效应开展系统评价。
本文选择位于云南省东南部且受到极端干旱气候明显影响的浴仙湖为研究对象, 通过提取湖泊沉积物并开展多指标综合分析, 结合气象监测等数据和主成分分析等方法, 系统识别极端干旱事件的沉积物信号、湖泊生物群落结构和多样性的响应模式, 从而为该地区应对气候变化与流域管理提供一定的基础数据和科学依据。
1 材料与方法
1.1 研究区概况
浴仙湖位于云南省文山州且属红河流域泸江水系, 是滇东南岩溶山原地区的一个断陷湖盆(北纬23°44′, 东经104°04′), 流域内主要分布岩溶地层发达的中低山山原地貌, 岩溶地质构造与石灰岩发育。湖泊水位在海拔1506 m时, 湖盆东西长2750 m,南北宽2250 m, 水域面积约2.27 km2, 湖水平均水深4.5 m, 最大水深15 m[23]。研究区地处北回归线附近低纬高原地带, 湖区属亚热带大陆性季风气候。年平均气温16.5℃, 极端高温出现在5—6月, 极端低温出现在1—2月, 年均降水量约900—1000 mm, 径流年变化主要受气候因素的控制, 湖水补给主要依赖地表径流及降水[24]。2013年8月的现代湖泊调查显示湖泊水位显著下降、湖泊水面面积锐减至约150 m2。
2013年的水质分析表明, 浴仙湖水体总磷为约3 μg/L、叶绿素含量约4 μg/L (表 1), 参照富营养水平的划分标准该湖泊目前为贫营养水体湖泊[25]。近60年来气象监测数据分析显示, 浴仙湖流域年平均气温总体呈升高的趋势, 降水量则呈现较大的波动(图 1)。在1951—1995年间年平均气温(±1 SD)为(18.6±0.4)℃, 且保持相对稳定, 但是在近20年来出现增温的趋势, 平均温度(±1 SD)为(19.2±0.6)℃, 且在2009—2010年达到最高年平均气温值(20.2℃)。而年降水量从1955年开始出现总体降低的趋势, 1995—2012年间年降水量(±1 SD)为(838.3±139.3) mm, 而2009—2012年期间年降水量持续偏低(843.5 mm)。

表 1 浴仙湖的湖泊学基本特征Tab. 1 Key limnological features of Yuxian Lake
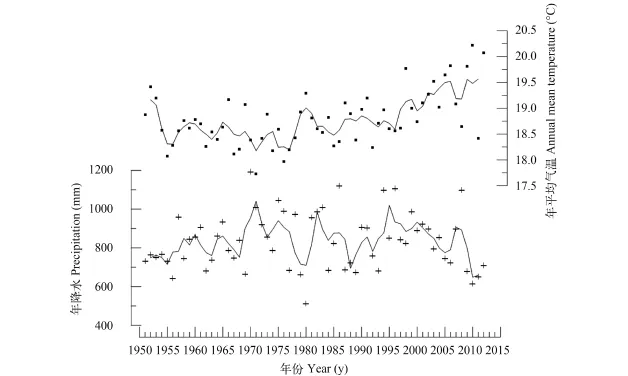
图 1 浴仙湖流域近60年来年平均气温、年降水及其3年滑动平均变化图Fig. 1 Annual mean temperature and precipitation over the past 60 years with 3-year running smooth lines
1.2 湖泊基本调查
2013年8月对浴仙湖开展野外调查工作, 采集了湖泊水样、同时现场采用YSI野外多参数水质仪获得部分水质数据。采集的水样置于4℃保温箱中至实验室进行水化学分析, 本文湖泊水质数据包括水深(H)、pH、叶绿素a (Chl.a)、NO3-N、NO2-N、NH3-N等理化指标(表 1)。
1.3 沉积物采样与实验室处理
沉积物采样 至2013年, 干旱导致湖泊面积骤减, 湖泊水体表面积不足150 m2(图 2)。为获取连续未受扰动的湖泊沉积物, 利用瑞典HTH重力采样器采集了长度为36 cm的柱状沉积岩芯一根(YX-1-GC-2; 23°44′N, 104°4′E, 水深: 0.2 m; 图 2)。现场对钻孔表层0—5 cm沉积物按0.5 cm间隔、深度5 cm以下按1 cm间隔分布进行分样, 样品避光保存于4℃并带回实验室进行冻干处理。实验室测试指标包括含水量测定、沉积物粒度、210Pb测年、碳氮元素、稳定同位素、硅藻群落等。以生物(硅藻)、物理化学指标为例识别近百年来浴仙湖生态环境的变化特征, 本文重点对浴仙湖钻孔上部30 cm样品(大约1900—2013 AD)进行系统分析。
实验室处理 沉积物样品采用210Pb和137Cs测年方法建立年代序列。具体分析步骤包括: 将冻干后的样品每次取少量压实装入测年样管中至约4 cm, 并将其表面处理水平, 向其注入约1 cm的环氧树脂AB结构胶; 封胶静置7d后, 放入美国Canberra公司的高纯锗探测器、数字化谱仪及多通道分析GAMMA仪器中测量放射性核素210Pb和137Cs比活度。最后应用恒定补给速率模型(CRS)建立钻孔的年代学序列[26]。
沉积物的粒度分析主要包括如下步骤: 称取干样0.3—0.5 g, 加入10 mL浓度10%的双氧水, 置于加热板上煮沸去除有机质, 至少量气泡为止, 再加入10 mL 10%的盐酸去除碳酸盐。充分反应后加入蒸馏水静置24h后去除上层废液。加入10 mL 0.05 mol/L的六偏磷酸钠分散剂后超声波振荡10min, 使用马尔文激光粒度仪(MS-2000)进行测量, 并使用粒径<2 μm、2—63 μm、>63 μm及中值粒径4个指标反映沉积物粒度分布特征。
沉积物全岩样品的碳、氮同位素分析分别采用国标标准VPDB和大气中的N2为参照[27,28]。主要步骤包括: 充分研磨约10 mg冻干全样样品, 并用锡箔杯包裹成球状; 之后将其送入反应炉中在960℃下燃烧, 产生的气体通过FLASH2000装置与Thermo MAT-253质谱仪进行同位素比值测定; 同时应用元素分析仪进行碳、氮元素含量测定。在本次分析中,沉积物同位素和元素含量的分析精度分别为±0.1‰和±1%。
沉积物硅藻样品参照国际通用方法进行处理[29]。具体步骤包括: 称取0.3—0.5 g湿样, 加10%盐酸溶液8 mL去除碳酸盐, 充分反应后加30%过氧化氢3 mL, 并将样品置于80℃水浴4h, 期间每过25min加3 mL过氧化氢, 充分去除硅藻细胞壁上的有机质。反应结束后以1200 r/min速度离心4min,抽出上层液体后加入蒸馏水混合, 重复清洗4次, 离心完成后加入几滴甲醛溶液密封低温保存。使用Naphrax®封片胶制作硅藻玻片, 硅藻鉴定分析用1000倍相差显微镜(Leica DM2500)油镜鉴定, 属种的鉴定以Krammer & Lange-Bertalot (1986—1999)[30]为主。依据硅藻保存情况, 每个样品的硅藻统计控制在400—420个, 属种丰度按百分比表示并进行群落结构分析。同时将壳体形态与生境相似的两个底栖物种F. construens和F. pinnata合并, 其中底栖硅藻种属按栖息地划分为静水水体和河流相两个硅藻类型[31]。
1.4 数据统计与分析
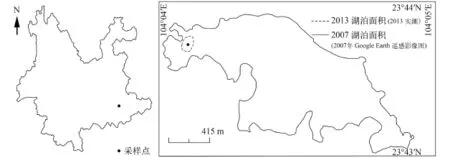
图 2 云南浴仙湖采样点位置示意图Fig. 2 Location and coring site of Yuxian Lake, Yunnan
硅藻地层图中选取百分比含量大于5%的硅藻物种, 并使用限制性聚类分析(CONISS)确认硅藻群落变化中具有统计学意义的显著时段。为探讨硅藻群落结构变化的主要方向, 对平方根转换后的群落数据进行主成分(PCA)分析。该方法是一种基于群落结构或组成数据而不需要考虑环境梯度的线性模型排序法[32,33]。硅藻群落的α多样性采用物种丰富度、Shannon指数和Pielou均匀度指数测度, β多样性采用Jaccard相异性指数测度, 硅藻物种丰富度指数进行稀有度校准后获得[34]。本文所有的统计分析均在R(3.2.2)软件中完成。
2 结果
2.1 钻孔年代序列
基于湖泊沉积物210Pb、137Cs比活度和CRS模型[35], 建立了浴仙湖近百年以来的年代序列(图 3)。浴仙湖沉积物样品的210Pb比活度随深度基本呈指数衰变, 但表层10 cm以上比活度偏低, 可能是由于干旱气候的持续使得沉积物样品210Pb信号受到流域过程变化的影响[36]。在浴仙湖沉积物中137Cs峰值出现在约14.5 cm(1986±2 AD; 图 3a), 与1986年4月苏联加盟共和国乌克兰的切尔诺贝利核电站核泄漏事件导致峰值相对应[37]。该研究区内大屯海[38]及长江中下游地区的太湖[39]同样检测到1986年左右的137Cs峰值, 而沉积物表层样品中137Cs出现的较高水平可能与流域地表侵蚀过程的增强相关, 与表层沉积物沉积速率明显增加的模式一致(图 3b)。
2.2 湖泊环境变化的沉积物记录
近百年来沉积物颗粒大小主要以粒径<63 μm的组分为主(图 4), 在约1968—2008年间沉积物粒度分布相对稳定。平均中值粒径(±1 SD)为(4.00± 0.16) μm, 主要以粉砂组分(2—63 μm)为主(约71%)。粗颗粒物质(粒度粒径>63 μm)百分比含量总体偏低(如<5%), 同时近10年来表现为总体增加、变化幅度增大的特征。近百年来沉积物中值粒径指标出现多次波动, 尤其在大概1920、1998、2011等年份出现峰值。
碳氮同位素数据显示近百年来δ13C和δ15N分别在–13.5‰—–18.0‰、4.0‰—5.5‰内波动, 其平均值(±1SD)分别为(–15.6±1.0)‰和(4.9±0.3)‰。其中δ13C总体呈下降趋势, 但幅度不大。δ15N则在1970—2000年左右出现低值(4.5±0.2)‰。C/N值在10—15间波动但没有显著的增加或降低趋势, 明显波动主要出现在约1900—1915年、约1995—2005年和约2011年。沉积物总碳(TC)和总氮(TN)含量在10—23、1—2变化, 平均值(±1SD)分别为(17±3)、(1±0) mg/g, 两个指标都保持相对稳定(图 4)。

图 3 浴仙湖钻孔沉积物年代序列Fig. 3 Depth profiles showing the radioactivities, age model and sedimentation rate for the sediment core of Yuxian Lake
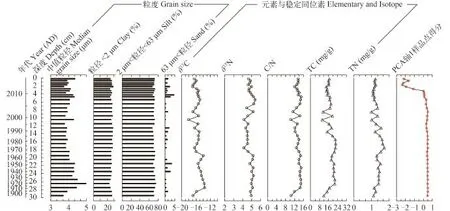
图 4 浴仙湖钻孔沉积物粒度、元素与稳定同位素随深度变化图Fig. 4 Stratigraphic profile showing the grain size, elemental and isotope data at Yuxian Lake
2.3 硅藻群落长期变化特征
浴仙湖钻孔中硅藻以底栖物种为主, 以Fragilaria construens/pinnata及Nitzschia palea为优势, 其次是Achnanthes minutissima、F. elliptica、F. pseudoconstruens、F. berolinensis、N. fruticosa和Surirella angusta (图 5)。浮游硅藻的百分比较低(<1.5%)且集中出现于约1963、1985年等, 浮游种主要有Aulacoseira ambigua和Cyclostephanos dubius。
限制性聚类分析并结合零模型检验的结果显示, 该钻孔硅藻群落结构可划分2个明显变化的阶段, 即在钻孔深度约3 cm (约2011年)前后出现重大转变。2011年以前, 硅藻组合以F. construens/pinnata和A. minutissima为绝对优势, 期间F. construens/ pinnata平均百分比为69.3%。2011年以后则以河流系统中常见的N. palea、N. fruticosa及S. angusta三个物种为优势, 且N. palea优势度高达40%。根据静水水体硅藻和河流相硅藻所占的百分比来看, 静水水体硅藻在约2011年开始大量减少, 而河流相硅藻则明显增加并成为优势物种。
硅藻数据PCA分析显示(图 6), 主轴1解了硅藻群落结构变化的58.9%, 主轴2解释了8.2%, 零模型检验表明仅有主轴1在统计学上是显著的。主成分分析双图中沿轴1负方向, 即2011—2013年以河流相硅藻N. palea、N. fruticosa及S. angusta为主(图 6)。而静水水体中常见的F. construens/pinnata和A. minutissima则分布于轴1反方向。
2.4 硅藻生物多样性指数
该钻孔硅藻物种丰富度、香农指数和均匀度指数呈现长期增加的趋势(图 7), 但物种丰富度自约2011年达到最大值后出现降低, 反映群落相异性的Jaccards指数在近百年来也呈现长期增加的趋势(图 7)。物种丰富度和香农指数在约1900年至2000年间保持相对稳定, 平均值(±1SD)分别为12±2、1.16±0.30; 2000年后开始增加, 期间平均值(±1SD)为19±5、1.60±0.48。近百年来硅藻群落的均匀度指数变化范围在0.3—0.8, 在1970s至2011年都有较大波动。约2011年以后, α多样性指数(主要为物种丰富度)出现降低趋势, 群落Jaccards相异性指数在0.24—0.67内变化, 约2011年出现了近百年来的最大值后总体下降。同时, PCA主轴1样品点得分与硅藻α多样性指数呈现显著相关的变化特征(如物种丰富度r=–0.79, P<0.001; 香农指数r=–0.78, P<0.001; 均匀度指数 r=–0.64, P<0.001)。
3 讨论
3.1 近百年来区域气候与湖泊环境变化的特征评价
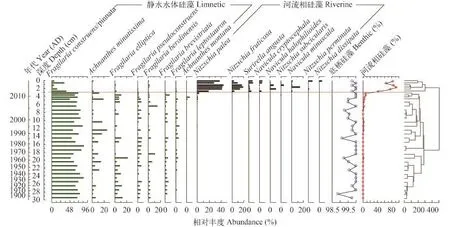
图 5 浴仙湖钻孔主要硅藻属种组合变化图与限制性聚类分析(CONISS)结果Fig. 5 Sedimentary profile showing the diatom assemblages for Yuxian Lake, with the red line indicating a significant species shift based on CONISS results
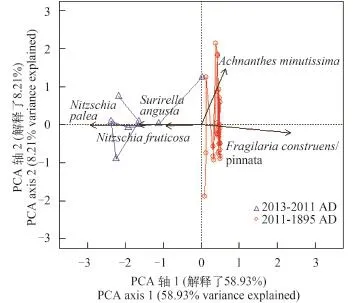
图 6 浴仙湖硅藻群落主成分分析双图Fig. 6 PCA biplot showing the first two axes for diatom assemblages in Yuxian Lake
沉积物粒度已有的研究结果表明, 沉积物粒径增大反映采样点离湖岸距离减小、湖泊水位下降,指示干旱气候; 反之, 沉积物粒径减小反映采样点离湖岸距离增大、湖泊水位上升[40,41]。如Wang等[42]对蒙古高原湖泊沉积环境记录的研究同样证明细粒沉积物对应较高的湖泊水位, 指示湿润的气候条件。与近60年的长期气温和降水平均值相比, 浴仙湖流域1970s—2000年期间年均气温偏低0.14℃, 降水增多约41.9 mm; 同期沉积物中值粒径整体偏小且较稳定, 与湖泊水位总体偏高相一致(即最大水深为约15 m)。而从约2011年(钻孔深度3 cm)开始,浴仙湖沉积物平均粒径呈现增大趋势(图 4)并与该区域极端干旱事件的出现相对应, 即从2009年年底开始流域气温总体升高、降水量大幅减少(图 1)。同时流域开发与农业用水的持续可能加速了湖泊水位的下降, 并缩短了流域地表颗粒物在湖泊水体中的搬运距离, 导致沉积物平均粒径的增大[40,41]。而距浴仙湖大约85 km的大屯海, 由于筑坝及干旱事件导致湖泊水动力减弱, 中值粒径则呈现减小的趋势[38], 可能反映了不同类型的湖泊粒度指标和水动力特征对极端干旱事件的差异性响应。
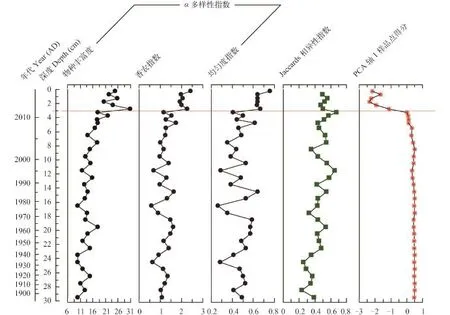
图 7 浴仙湖硅藻群落物种多样性指数与PCA样品点得分图Fig. 7 Stratigraphic profiles showing the diatom diversity indices and the PCA axis one site scores for Yuxian Lake
沉积物元素、稳定同位素记录及现代监测数据均表明, 浴仙湖近百年来水体营养水平总体偏低且长期变化的幅度较小。湖泊沉积物中元素组成(TC、TN、C/N)与碳、氮同位素(δ13C、δ15N)可以用来重建湖泊古生产力变化并探讨沉积物来源[43]。近百年来浴仙湖沉积物C/N平均值为13.4, 表明该湖主要以内源有机质(如藻类、水生植物)输入为主, 而δ13C、δ15N没有明显的增加或降低指示湖泊生产力没有显著变化。太湖不同湖区稳定同位素δ13C、δ15N的研究显示湖泊由贫营养至富营养的过程中δ13C和δ15N值增加[44]。已有研究表明, 湖泊典型藻类C/N约在5—10, 但是陆地维管束植物C/N值往往高于20[28,45], 而藻类生产力的增加可以使δ13C值降低[46]。卢慧斌等[47]研究表明, 近百年来洱海沉积物TN增大及浮游动物枝角类生物量的增加指示了湖泊富营养化的长期过程; 同时长江中下游太湖近百年来C/N的降低和TC、TN的增加示踪了湖区富营养化历史[39]。而浴仙湖沉积物TC和TN值没有明显升高的趋势, 这也与2013年水质监测数据(表 1)显示的该湖泊为贫营养水平一致。
3.2 近百年来湖泊生态系统变化特征
湖泊水深与栖息地变化是影响硅藻组成的主要环境因子。国内对硅藻与水深的研究大多集中于湖泊盐度的重建, 如胡晓兰等[48]对西藏多个湖泊表层沉积物硅藻的研究以Navicula、Fragilaria和Nitzschia为主, 但硅藻空间组合分布主要受水体盐度、碱度的控制。抚仙湖等云南地区的深水湖泊硅藻群落主要以浮游物种为主[49], 而浴仙湖的硅藻优势种以附生种(如F. construens/pinnata、A. minutissima)和河流相物种(如N. palea、N. fruticosa及S. angusta)为主, 且浮游种百分比很低(小于1.5%)。类似的, 位于云南西北部且营养水平偏低的云龙天池硅藻群落的底栖优势种同样以F. construens、F. pinnata和A. minutissima为主[50]。由于本研究提取的钻孔位于浴仙湖西北部近岸浅水区域, 因此硅藻群落以附生物种为主与北半球浅水湖泊硅藻群落的结构特征类似[51], 而少量的浮游物种的出现可能是通过水体或风力作用由敞水区带至沿岸带[52]。
同时, 浴仙湖硅藻群落的演替与优势物种的转换特征表现为对极端干旱胁迫的明显响应。1960年以来云南经历了四次明显的极端干旱事件,而2009年秋冬开始的极端干旱事件最为严重[3], 从而导致了湖泊环境与生态系统的明显变化。与浴仙湖流域气象资料、湖泊调查等结果相一致, 沉积物粒度分析显示了此次极端干旱气候导致的降水量减少、湖泊水位的显著降低(图 1、图 4)。地表水体蒸发量的增加与人为引水量剧增等叠加效应造成了湖泊面积锐减。同时浴仙湖生态系统结构出现显著变化, 如硅藻群落由约2011年以前由湖泊中常见的底栖物种F. construens/pinnata及A. minutissima迅速转变为以河流相物种N. palea和N. fruticosa为主(图 3), 同时该时间点也是硅藻群落相异性指数最大、PCA轴1样品点出现显著变化的阶段(图 6、图 7)。此次干旱事件起始于2009年秋季(10月左右), 干旱最为严重时间为2010—2012年[4]。基于210Pb建立的CRS年代模型在表层存在±1年的误差(图 3), 因此2009—2013年干旱事件及其生态环境影响在本文以约2011年为界。浮游硅藻百分比变化主要时间为1990s—2011年, 在湖泊水位和生态环境变化最大的2011年未出现显著变化, 因此该湖中浮游硅藻对干旱事件没有显著指示价值(图 5)。该区域内星云湖沉积物多指标记录显示, 全球变暖与极端气候的出现是影响云南中部地区湖泊硅藻群落变化的重要环境因子之一[53]。Laird等[54]利用沉积物硅藻重建了北美大草原过去2300年干旱强度和频率, 表明过去1200AD年北美大草原的干旱强度和频率主要受长期气候变化的驱动。1900—2011年期间, 浴仙湖钻孔硅藻组成主要以F. construens/pinnata为优势种; 类似的, 加拿大Worth湖的研究表明优势种F. pinnata主要分布在中等深度区(即约3—9 m)且适宜在低光条件下生长[51]。同时, 硅藻的生长与群落分布还受到湖泊栖息地与水动力条件的影响, 如附生型硅藻Achnanthes通常喜欢近岸生境[55], 而河流相硅藻(如Navicula和Nitzschia)能适应较强的水动力特征并在较低的光照条件下具有竞争优势[56]。因此在浴仙湖中, 极端干旱事件的持续与湖泊水位大幅下降、水动力增强, 都有利于Navicula、Nitzschia等硅藻种属在约2011年开始出现快速增加并成为优势物种(图 5)。
3.3 硅藻生物多样性响应极端干旱事件的变化模式
已有研究表明, 环境异质性与生态位分化是影响生物多样性模式的重要生态过程[57—59]。浴仙湖硅藻群落α多样性随时间呈现增加的特征, 可能反映了湖泊栖息地结构的变化与水体环境异质性的增加[51,60]。硅藻物种分布与水深的大量研究表明,湖泊底栖硅藻与浮游藻类相比具有较高的物种多样性[55,61]。湖滨浅水区拥有多样的生境(如沙质、泥质、水生植物等)和复杂的环境特征, 且光照条件多样、营养盐较高[55,62]。而云南西北部小型深水湖泊的硅藻群落多样性的长期变化受水位的波动影响较小[50], 反映了水文波动可能导致了不同水深的中小型湖泊中硅藻群落多样性呈现出差异的响应模式。
时空尺度上群落结构变化程度的β多样性指标[34], 可以直接指示主要的环境驱动因子[18,63]。浴仙湖沉积物记录表明, 硅藻群落β多样性数值在约2011年左右达到最大, 与硅藻群落中静水水体常见物种大量减少、河流相硅藻成为优势物种时间一致, 而2011年以来硅藻群落同质性增加、群落相异性指数降低(图 7)。该响应模式与中等干扰假说(Intermediate disturbance hypothesis)的预测模型相一致, 即中等程度的干扰频率能维持较高的物种多样性[64]。而Chase等[65]的研究同时表明, 生境条件的持续恶化会降低群落构建中随机过程的重要性、环境异质性降低, 从而加强了群落的均质化、降低群落β多样性指数。类似的, 随着滇池富营养化的持续与系统生产力的上升, 种间竞争压力的增强、水体透明度的降低、水生植被覆盖度的减少等过程直接导致了沉积物硅藻群落β多样性指数的持续降低[49]。因此, 云南湖泊硅藻群落在环境压力持续的背景下呈现了群落结构异质性的长期降低,反映了气候变化持续、人类活动增强等背景下该生物多样性热点地区面临着多样性保护的严峻挑战。
4 结论
区域与全球尺度上极端干旱事件发生的频率与强度增加, 导致了云南大量湖库的水位下降、水资源总量明显降低, 对区域社会经济发展与生态环境功能产生了威胁。本研究对浴仙湖沉积物多指标记录的研究表明, 近百年来浴仙湖环境的主要变化出现于2009—2013年极端干旱事件期间, 湖泊水位的快速降低导致了沉积物粒径的明显变化与硅藻群落的快速演替, 同时硅藻生物多样性呈现增加的趋势, 表明了极端干旱事件对湖泊生物群落构建的胁迫作用和对湖泊生态系统结构的驱动影响。因此, 2009—2013年极端干旱事件导致了浴仙湖生物群落结构的快速变化是过去100年里没有出现过的系统突变事件, 这表明积极采取气候适应对策是对这一气候敏感地区进行湖泊管理与生态修复的重要任务。
致谢:
感谢高原地理过程与环境变化云南省重点实验室赵帅营、段立曾、王教元、施海彬在野外采样、室内实验及数据分析等方面提供的帮助。
[1]Lake P S. Drought and Aquatic Ecosystems Effects and Responses [M]. Wiley-Blackwell A Jhon Wiley & Sons, Ltd., Publication. 2011, 2—3
[2]Duan X, Tao Y, Duan C C. A fine mesh climate division and the selection of representative climate stations in Yunnan Province [J]. Journal of Nanjing Institute of Meteorology, 2011, 34(3): 336—342 [段旭, 陶云, 段长春. 云南省细网格气候区划及气候代表站选取. 大气科学学报, 2011, 34(3): 336—342]
[3]Yang H, Song J, Yan H M, et al. Cause of the Severe Drought in Yunnan Province during Winter of 2009 to 2010 [J]. Climatic and Environmental Research, 2012, 17(3): 315—326 [杨辉, 宋洁, 晏红明, 等. 2009/2010年冬季云南严重干旱的原因分析. 气候与环境研究, 2012, 17(3): 315—326]
[4]Wang W, Wang W J, Li J S, et al. The Impact of Sustained Drought on Vegetation Ecosystem in Southwest China Based on Remote Sensing [J]. Procedia Environmental Sciences, 2010, 2(6): 1679—1691
[5]Zhang L, Xiao J F, Li J, et al. The 2010 spring drought reduced primary productivity in southwestern China [J]. Environmental Research Letters, 2012, 7(4): 45706
[6]Wang H, Yang Z X, Wang L, et al. The application of TVDI in drought monitoring over Yunnan province during 2009 to 2010 [J]. Journal of Yunnan University (Natural Sciences), 2014, 36(1): 59—65 [王海, 杨祖祥, 王麟,等. TVDI在云南2009/2010年干旱监测中的应用. 云南大学学报(自然科学版), 2014, 36(1): 59—65]
[7]Fu B J, Niu D, Zhao S D. Study on global change and terrestrial ecosystems: history and prospect [J]. Advances in Earth Science, 2005, 20(5): 556—560 [傅博杰, 牛栋, 赵士洞. 全球变化与陆地生态系统研究: 回顾与展望. 地球科学进展, 2005, 20(5): 556—560]
[8]Zhang B, Zhu J J, Liu H M, et al. Effects of extreme rainfall and drought events on grassland ecosystems [J]. Chinese Journal of Plant Ecology, 2014, 38(9): 1008—1018 [张彬, 朱建军, 刘华民, 等. 极端降水和极端干旱事件对草原对草原生态系统的影响. 植物生态学报, 2014, 38(9): 1008—1018]
[9]Tian H Q, Xu X F, Song X. Drought impacts on terrestrial ecosystem productivity [J]. Journal of Plant Ecology (Chinese Version), 2007, 31(2): 231—241 [田汉勤, 徐小锋, 宋霞. 干旱对陆地生态系统生产力的影响. 植物生态学报, 2007, 31(2): 231—241]
[10]Guo H C, Wang X Y, Yi X. Study on eutrophication control strategy based on the succession of water ecosystem in the Dianchi Lake [J]. Geographical Research, 2013, 32(6): 998—1006 [郭怀成, 王心宇, 伊璇. 基于滇池水生态系统演替的富营养化控制策略. 地理研究, 2013, 32(6): 998—1006]
[11]Dai G Y, Li J, Li L, et al. The spatio-temporal pattern of phytoplankton in the north basin of Lake Dianchi and related environmental factors [J]. Acta Hydrobiologica Sinica, 2012, 36(5): 946—956 [代龚圆, 李杰, 李林, 等. 滇池北部湖区浮游植物时空格局及相关环境因子. 水生生物学报, 2012, 36(5): 946—956]
[12]Shi H B, Chen G J, Liu Y Y, et al. Long-term pattern and driving factors of cladoceran community changes in Lake Xingyun, Yunnan [J]. Chinese Journal of Ecology, 2015, 34(9): 2464—2473 [施海彬, 陈光杰, 刘园园, 等. 星云湖枝角群落变化的长期特征与驱动因素. 生态学杂志, 2015, 34(9): 2464—2473]
[13]Smol J P. Pollution of Lakes and Rivers: A Paleoenvironmental Perspective. 2nd edn [M]. Blackwell Publishing, Oxford. 2008, 13—22
[14]Wu D, Zhou A, Liu J B et al. Changing intensity of human activity over the last 2000 years recorded by the magnetic characteristics of sediments from Xingyun Lake, Yunnan, China [J]. Journal of Paleolimnology, 2016, 53(1): 47—60
[15]Stoermer E F, Smol J. The Diatoms: Applications for the Environmental and Earth Sciences [M]. Cambridge: Cambridge University Press. 1999, 15—65
[16]Hargan K E, Nelligan C, Jeziorski A et al. Tracking the long-term responses of diatoms and cladoceran to climate warming and human influences across lakes of the Ring of Fire in the Far North of Ontario, Canada [J]. Journal of Paleolimnology, 2016, 56(2): 153—172
[17]Rü hland K M, Paterson A M, Smol J P. Lake diatom responses to warming: reviewing the evidence [J]. Journal of Paleolimnology, 2015, 54(1): 1—35
[18]Smol J P, Wolfe A P, Birks H J, et al. Climate-driven regime shifts in the biological communities of arctic lakes [J]. Proceedings of the National Academy of Sciences, 2005, 102(12): 4397—4402
[19]Lu Y B, Rioual P, Yang X P. An Assessment of Diatom Presence/Absence in the Lakes of the Badain Jaran Desert [J]. Quaternary Sciences, 2010, 30(3): 625—627 [卢彦斌, Rioual P, 杨小平. 巴丹吉林沙漠湖泊表层硅藻分布的初步研究. 第四季研究, 2010, 30(3): 625—627]
[20]Chen X, Yang X D, Dong X H, et al. Effects of environmental changes on the succession of diatom assemblage during the last 50 years in Lake Chaohu [J]. Journal of Lake Science, 2011, 23(5): 665—672 [陈旭, 羊向东, 董旭辉, 等. 近50年来环境变化对巢湖硅藻组合演替的影响. 湖泊科学, 2011, 23(5): 665—672]
[21]Chen C H, Zhao L Y, Zhu C, et al. Response of diatom community in Lugu Lake (Yunnan-Guizhou Plateau, China) to climate change over the past century [J]. Journal Paleolimnology, 2014, 51(3): 357—373
[22]Hu Z J, Li Y L, Li S X. Spatial and temporal distributions of diatom communities and their relationships with environmental factors in Lake Erhai [J]. Journal of Lake Science, 2012, 24(3): 400—408 [胡竹君, 李艳玲, 李嗣新. 洱海硅藻群落结构的时空分布及其与环境因子间的关系. 湖泊科学, 2012, 24(3): 400—408]
[23]Yang L, Li H, Yang X J, et al. Wetlands of Yunnan [M]. Beijing: China Forestry Publishing House. 2010, 153—154 [杨岚, 李恒, 杨晓君, 等. 云南湿地. 北京: 中国林业出版社. 2010, 153—154]
[24]The Compilation Committee of Yanshan County. Yanshan Year Book [M]. Yunnan: Yunnan People’s Publishing Press. 2010, 66 [砚山县志编纂委员会. 2000. 砚山县志. 云南: 云南人民出版社. 2010, 66]
[25]Qin B Q, Xu H, Dong B L. Eutrophication of Lakes Governance Theory and Practice [M]. Beijing: Higher Education Press. 2011, 2—3 [秦伯强, 许海, 董百丽. 富营养化湖泊治理的理论与实践. 北京: 高等教育出版社. 2011, 2—3]
[26]Appleby P G. Chronostratigraphic Techniques in Recent Sediments. In: Last W M, Smol J P (Eds.), Tracking Environmental Change Using Lake Sediments. Vol 1: Basin Analysis, Coring and Chronological Techniques, vol 1 [M]. Kluwer, Dordrecht. 2001, 171—203
[27]Zhu Y, Lei G L, Lin Y Y, et al. Element analysis-isotope mass spectrometry system: an alternative method for measuring total nitrogen contents [J]. Journal of Fujian Normal University (Natural Science Edition), 2013, 29(1): 58—62 [朱芸, 雷国良, 林燕语, 等. 元素分析仪-同位素质谱测定全氮含量的方法研究. 福建师范大学:自然科学版, 2013, 29(1): 58—62]
[28]Meyers P A. Preservation of elemental and isotopic source identification of sedimentary organic matter [J]. Chemical Geology, 1994, 114(3—4): 289—302
[29]Battarbee R W, Jones V J, Flower R J, et al. Diatoms Terrestrial, Algal and Siliceous Indicators. Tracking Environmental Change Using Lake Sediments [M]. Dordrecht: Kluwer Academic Publishers. 2001, 155—202
[30]Krammer K, Lange-Bertalot H. Bacillariophyceae (1—4 Teil). In: Ettl H, Gedof J, H eynig H, et al (Eds.), Sü sswasserflora von Mitteleuropa [M]. Stuttgart and Jena: Gustav Fischer Verlag. 1986—1991, 876, 596, 576, 473
[31]Stevenson R J, Pan Y. Assessing environmental conditions in rivers and streams with diatoms. In: E F Stoermer, J P Smol (Eds.), The Diatoms: Applications for the Environmental and Earth Sciences [M]. Cambridge, UK: Cambridge University Press. 1999, 11—40
[32]ter Braak C J F, Prentice I C. A theory of gradient analysis [J]. Advances in Ecological Research, 1988, 18(2004): 271—313
[33]ter Braak C J F, Šmilauer P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination [Version 4.5] [M]. Microcomputer Power, Ithaca, NY. 2002, 69—77
[34]Magurran A E. Measuring Biological Diversity [M]. Oxford: Blackwell Publication. 2004, 48—67
[35]Ding Z Y, Yang H, Wang X L, et al. Determination of Sedimentation Rates in Higher-up Lake Using137Cs,241Am and210Pb Dating Techniques [J]. Geography and Geo-Information Science, 2012, 28(5): 90—94 [丁兆运,杨浩, 王小雷, 等. 基于137Cs、241Am和210Pb计年的上级湖沉积速率研究. 地理与地理信息科学, 2012, 28(5): 90—94]
[36]Zhang X B, Long Y, Wen A B, et al. Discussion on applying137Cs and210Pbex for lake sediment dating in China [J]. Quaternary Sciences, 2012, 32(3): 430—440[张信宝, 龙翼, 文安邦, 等. 中国湖泊沉积物137Cs和210Pb断代的一些问题. 第四纪研究, 2012, 32(3): 430—440]
[37]Zeng L, Wu F C, Wan G J, et al. The distribution characteristic and environmental significance of Cesium-137 deposit profile in Chinese lacustrine sediment [J]. Journal of Lake Sciences, 2009, 21(1): 1—9
[38]Tao J S, Chen G J, Chen X L, et al. Long-term pattern of diatom community structure changes in response to multiple environmental stressors at Datun Lake, Southeast Yunnan, China [J]. Chinese Journal of Applied Ecology, 2015, 26(8): 2525—2533
[39]Liu J J, Wu J L. Environmental information of recent 100 years recorded sediments of Dapu area in Taihu Lake [J]. Journal of Palaeogeography, 2006, 8(4): 559—564 [刘建军, 吴敬禄. 太湖大浦区近百年来湖泊记录的环境信息.古地理学报, 2006, 8(4): 559—564]
[40]Xiao J L, Fan J W, Zhou L et al. A model for linking grain-size component to lake level status of a modern clastic lake [J]. Journal of Asian Earth Science, 2014, 355(12): 149—158
[41]Yi L, Yu H J, Ortiz J D et al. A reconstruction of late Pleistocene relative seal level in the south Bohai Sea, China, based on sediment grain-size analysis [J]. Sedimentary Geology, 2012, 281(24): 88—100
[42]Wang H Y, Liu H Y, Cui H T, et al. Terminal Pleistocene/Holocene palaeoenvironmental changes revealed by mineral-magnetism measurements of lake sediments for Dali Nor area, southeastern Inner Mongolia Plateau, China [J]. Palaeogeography Palaeoclimatology Palaeoecology, 2001, 170(1—2): 115—132
[43]Zhou Z H, Li J, Zhu Z Z. Environmental evolution of Longgan Lake sediments recorded by carbon and nitrogen isotopes [J]. Chinese Journal of Ecology, 2007, 26(5): 693—699 [周志华, 李军, 朱兆洲. 龙感湖沉积物碳、氮同位素记录的环境演化. 生态学杂志, 2007, 26(5): 693—699]
[44]Wu J L, Lin L, Liu J J, et al. Environmental significance and stable isotope signature from sedimented organic matter in Lake Taihu [J]. Marine Geology and Quaternary Geology, 2005, 25(2): 25—30 [吴敬禄, 林琳, 刘建军, 等. 太湖沉积物碳氮同位素组成特征与环境意义.海洋地质与第四纪地质, 2005, 25(2): 25—30]
[45]Meyers P A, Teranes J L. Sediment Organic Matter [M]. In: Last W M, Smol J P (Eds.), Developments in Paleoenvironmental Research. 2001, 239—269
[46]Bunting L, Laevitt R P, Gibson C E, et al. Degradation of water quality in Lough Neagh, Northern Ireland, by diffuse nitrogen flux from a phosphorus-rich catchment [J]. Limnology and Oceanography, 2007, 52(52): 354—369
[47]Lu H B, Chen G J, Cai Y F, et al. Cladoceran community responses to eutrophication, fish introduction and macrophyte degration over the past century in Lake Erhai [J]. Journal of Lake Science, 2016, 28(1): 132—140 [卢慧斌,陈光杰, 蔡燕凤, 等. 近百年来枝角群落响应洱海营养水平、外来鱼类引入以及水生植被变化的特征. 湖泊科学, 2016, 28(1): 132—140]
[48]Hu X L, Wang C H, Ren Y Q, et al. Modern diatom combination characters and an influence factor analysis of the surface sediment of lakes in Tibet [J]. Journal of Lanzhou University (Natural Sciences), 2014, 50(4): 455—464 [胡晓兰, 王彩红, 任雅琴, 等. 西藏湖泊表层沉积物硅藻组合及其环境影响因素. 兰州大学学报(自然科学版), 2014, 50(4): 455—464]
[49]Chen X L, Chen G J, Lu H B, et al. Long-term diatom biodiversity responses to productivity in lakes of Fuxian and Dianchi [J]. Biodiversity Science, 2015, 23(1): 89—100 [陈小林, 陈光杰, 卢慧斌, 等. 抚仙湖和滇池硅藻生物多样性与生产力关系的时间格局. 生物多样性, 2015, 23(1): 89—100]
[50]Zou Y F, Yan Y, Zhang J Y, et al. The relationship between water depth and diatom biodiversity of Yunlong Lake, Yunnan Province [J]. Quaternary Sciences, 2015, 35(4): 988—996 [邹亚菲, 严瑶, 张佼杨, 等. 云龙天池湖泊水深与硅藻生物多样性的关系. 第四纪研究, 2015, 35(4): 988—996]
[51]Laird K R, Kingsbury M V, Cumming B F. Diatom habitats, species diversity and water-depth inference models across surface-sediment transects in Worth Lake, northwest Ontario. Canada [J]. Journal of Paleolimnology, 2010, 44(4): 1009—1024
[52]Vilmi A, Karjalainen S M, Hellsten S, et al. Bioassessment in a metacommunity context: Are diatom communities structured solely by species sorting [J]? Ecological Indicators, 2015, 2016(62): 86—94
[53]Liu Y Y, Chen G J, Shi H B, et al. Responses of a diatom community to human activities and climate changes in Xingyun Lake [J]. Acta Ecologica Sinica, 2016, 36(10): 3063—3075 [刘园园, 陈光杰, 施海彬, 等. 星云湖硅藻群落响应近现代人类活动与气候变化的过程. 生态学报, 2016, 36(10): 3063—3075]
[54]Laird K R, Haig H A, Ma S, et al. Expanded spatial extent of the Medieval Climate Anomaly revealed in lakesediment records across the boreal region in northwest Ontario [J]. Global Change Biology, 2012, 18(9): 2869—2881
[55]Hoagland K D, Peterson C G. Effects of light and wave disturbance on vertical zonation of attached microalgae in a large reservoir [J]. Journal of Phycolagy, 1990, 26(3): 450—457
[56]Stevenson R J, Stoermer E F. Quantitative differences between benthic algal communities along a depth gradient in Lake Michigan [J]. Journal of Phycology, 1981, 17(1): 29—36
[57]Dodson S I, Arnott S E, Cottingham K L. The relationship in lake communities between primary productivityand species richness [J]. Ecology, 2000, 81(10): 2662—2679
[58]Mcclain C R, Barry J P, Eernisse D, et al. Multiple processes generate productivity-diversity relationships in experimental wood-fall communities [J]. Ecology, 2016, 97(4): 885—898
[59]Forrester D I, Bauhus J. A review of processes behind diversity-productivity relationships in forests [J]. Current Forestry Reports, 2016, 2(1): 45—61
[60]Zhang M, Yu Y, Qian S Q, et al. Phytoplankton community structure and biodiversity in summer Yunnan-Guizhou Plateau lakes [J]. Journal of Lake Sciences, 2010, 22(6): 829—836
[61]Cantonati M, Scola S, Angeli N, et al. Environmental controls of epilithic diatom depth-distribution in an oligotrophic lake characterized by marked water-level fluctuations [J]. European Journal of Phycology, 2009, 44(1): 15—29
[62]Punning J M, Puusepp L. Diatom assemblages in sediments of Lake Juusa, Southern Estonia with an assessment of their habitat [J]. Hydrobiologia, 2007, 586(1): 27—41
[63]Hobbs J P A, Frisch A J, Allen G R. Marine hybrid hotspot at Indo-Pacific biogeographic border [J]. Biology Letters, 2009, 5(2): 258—261
[64]Connell J H. Diversity in tropical rain forests and coral reefs [J]. Science, 1978, 199(4335): 1302—1310
[65]Chase J M. Drought mediates the importance of stochastic community assembly [J]. Proceedings of the National Academy of Sciences (USA), 2007, 104(44): 17430—17434
SEDIMENT RECORDS OF EXTREME DROUGHTS AND ECOLOGICAL CONSEQUENCES IN YUXIAN LAKE, SOUTHEAST YUNNAN
HU Kui, CHEN Guang-Jie, HUANG Lin-Pei, CHEN Xiao-Lin, LIU Yuan-Yuan, LU Hui-Bin, TAO Jian-Shuang and KANG Wen-Gang
(Key Laboratory of Plateau Geographical Processes and Environmental Change, School of Tourism and Geography, Yunnan Normal University, Kunming 650500, China)
The increased intensity and frequency of extreme droughts due to global warming have led to the dedine of water levels and significant shortage of water resources in many lakes and reservoirs, which could severely threaten the socio-economic development and ecological security. To date the ecological evaluation of extreme droughts have been mainly focused on terrestrial ecosystems in China. This study aim to assess the impact of extreme droughts on diatom assemblages and diversity patterns through multi-proxy sediment analyses in Yuxian Lake, which has experienced significant water level change during the period of 2009 to 2013. The grain size records showed an increase in sediment particles starting from about 2011, which corresponded to the occurrence of the most recent drought event. The isotopic and elementary records indicated that the trophic state remained oligotrophic over the past century. Diatom assemblages were dominated by benthic and limnetic diatoms (i.e. Fragilaria construens/pinnata), but these taxa were there after replaced by fluvial and riverine taxa (i.e. Nitzschia and Navicula) with the most recent decline of lake levels. Our results demonstrated that the diatom PCA axis was strongly associated with extreme climate, which accounted for 55.8% of the total variance for diatom assemblages. Our study provided sediment evidence for the role of extreme droughts in regulating freshwater environment and biota and highlighted the urgency of climate mitigation actions in maintaining water security and ecosystem functioning for this climate-sensitive region.
Yuxian Lake; Extreme drought; Sediments; Diatoms; Biodiversity; Grain size
X524
A
1000-3207(2017)03-0724-11
10.7541/2017.90
2016-05-27;
2016-10-25
国家自然科学基金(U1133601、41171048、41302151); 科技部国家重点基础研究发展计划(2014CB460607); 教育部科学技术研究项目(213034A)资助 [Supported by the National Natural Science Foundation of China (U1133601, 41171048, 41302151); Ministry of Science and Technology of China (2014CB460607); Ministry of Education of China (213034A)]
胡葵(1991—), 女, 云南德宏人; 硕士研究生; 主要从事淡水生态研究。E-mail: hukuian@163.com
陈光杰, E-mail: guangjiechen@gmail.com
