红松不同种源的遗传多样性分析
2017-05-11张巍王清君郭兴
张巍,王清君,郭兴
(伊春林业科学院,黑龙江 伊春 153000)
红松不同种源的遗传多样性分析
张巍,王清君,郭兴
(伊春林业科学院,黑龙江 伊春 153000)
红松是小兴安岭林区的主要建群树种,在小兴安岭总体生态系统中具有重要的生态地位。为了保证小兴安岭林区红松的遗传丰富度,为小兴安岭林区提供优质的红松造林苗木。研究运用10个经过筛选后的引物,用ISSR分析方法对10个不同红松天然林种源进行了分子标记分析。结果显示:①10个ISSR引物共检测到的位点数在2~13之间,各位点的 DNA 片段长度介于 150~2 000 bp 之间。②共检测到79个位点,平均每个 ISSR 引物检测到9.9个位点。③多态性条带为65条,多态性位点百分率为42.63%。④各种源在分子层面存在明显差异,而这一差异与地理分布呈正相关。⑤小兴安岭林区红松群落的遗传丰富度较高,这也有效的保证了小兴安岭红松群落的林分质量。
红松;遗传丰富度;ISSR
0 引言
红松(Pinuskoraiensis)是我国珍贵的用材树种[1-2],而天然阔叶红松林是经过长期自然选择形成的原始群落[3],同时也是小兴安岭林区的地带性顶级群落[4]。小兴安岭伊春林区是我国红松的主分布区之一,曾经分布着大面积的红松原始林,但20世纪60、70年代经受了严重破坏。虽然现在每年都有大面积的造林计划进行,但伊春林区的红松原始林目前仍然处于恢复阶段。而随着造林任务的深入,不同种源的遗传丰富度表现出一定差异,造林质量也存在差异。近年来,如何有效的保存现有红松原始林的遗传多样性,提高阔叶红松林的遗传丰富度,一直是林业工作者共同努力的方向。为了揭示红松不同种源间遗传多样性的差异,本研究收集了包括小兴安岭、长白山等红松主分布区在内的天然红松母树林优树子代,用ISSR-PCR分子标记分析方法对各种源及种源内家系进行了分子标记分析,希望可以为红松不同种源的综合评价及利用提供参考和理论依据。
1 材料与方法
1.1 试验材料选择
试验共选定10个地区的红松原始林为种源选择区,各种源区具体情况见表1。
依据种源区域,选择铁力林业局种源家系8株,五营林业局种源家系10株,汤旺河林业局种源家系12株,带岭林业局种源家系9株,新青林业局种源家系10株,吉林省长白山1号种源家系10株(吉林,安图),吉林省长白山2号种源家系10株(吉林,蛟河),双鸭山市种源家系10株,友好林业局种源家系10株,牡丹江市种源家系10株,共99株单株样本。

表1 种源区立地条件统计
1.2 反应体系建立
1.2.1 红松总DNA提取
试验参考冯富娟等的试验方法[4-9]。采用CTAB提取红松针叶总DNA,从上清液中获取的DNA得率高,DNA电泳条带清晰。
1.2.2 ISSR-PCR
反应体系为20 μL,包括1×Buffer,1.5 mmol/L MgCl2,引物1.5 ng/(μL),dNTP的浓度为0.2 mmol/L,TaqDNA聚合酶的用量为0.5U,模板DNA的用量为20~40 ng,TaqDNA聚合酶、引物、dNTP由上海生工生物工程公司提供。见表2。
PCR反应程序为:94℃ 预扩增7 min;94℃变性 30s,52℃退火 45s,72℃延伸 2 min,40个循环;72℃ 7 min;4℃保存。
1.2.3 电泳检测及数据处理
PCR产物用1.5%琼脂糖凝胶,在不超过5 V/cm的电压下电泳,电极缓冲液为1×TAE,凝胶成像系统拍照记录。
1.2.4 DNA指纹图谱建立
将琼脂糖凝胶上出现DNA片段的记为1,不出现的为0,统计后输入电脑,用NTSYS聚类分析软件进行UPGMA法聚类分析,用POPGENE32软件进行数据处理。

表2 ISSR引物编号及序列
2 结果与分析
2.1 种源间ISSR的多态性
对10个种源进行PCR扩增。扩增片段分子量主要集中在200~1 500 bp之间。共扩增片段条带501条,其中多态性条带289条。各引物扩增出的条带数目从20至84不等。引物p13扩增出的条带数最多,为49条。引物P807扩增出891个遗传位点,为10个引物中最多(图1~图4)。以上数据表明红松种间在分子水平上的多态性是丰富的,利用ISSR分子标记能够检测红松多种源间的亲缘关系[10-13](表3)。

图1 天然红松ISSR电泳图谱,引物p5Fig.1 Pine ISSR gel electrophoresis primer P5

图2 天然红松ISSR电泳图谱,引物p11Fig.2 Pine ISSR gel electrophoresis primer P11

图3 天然红松ISSR电泳图谱,引物p13Fig.3 Pine ISSR gel electrophoresis primer P13

图4 天然红松ISSR电泳图谱,引物p807Fig.4 Pine ISSR gel electrophoresis primer P807
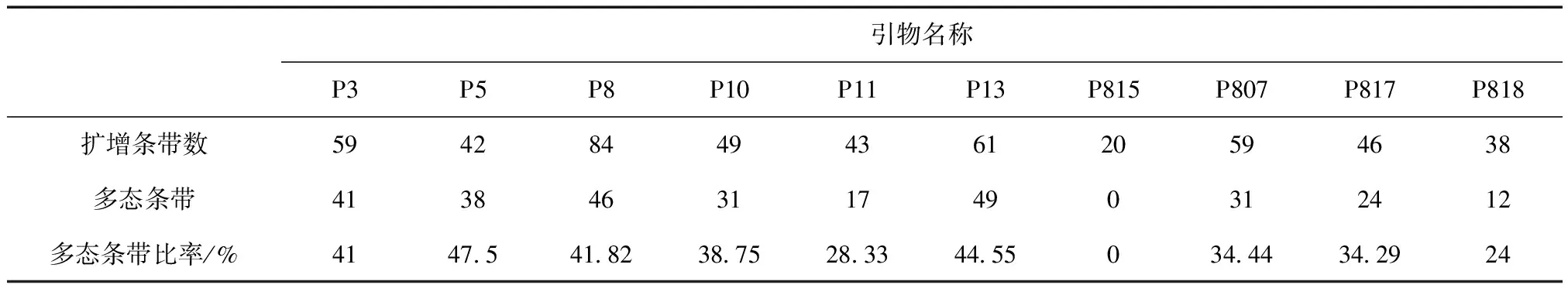
引物名称P3P5P8P10P11P13P815P807P817P818扩增条带数59428449436120594638多态条带4138463117490312412多态条带比率/%41475418238752833445503444342924
2.2 遗传多态位点分析
遗传多样性是生物多样性的基础,也是生物多样性最重要的部分[13-15]。通过对10个种源的遗传多样性进行统计分析表明,汤旺河种源区遗传性最为丰富,其总条带数为34条,多态性条带数42条,多态性条带百分比53.16%,该种区为各种源最高,其次为带岭种源区,总条带数为44条,多态性条带35条,多态性条带百分比为44.30%。五营种源区总条带数46,其中多态性条带数33,多态性条带比率为41.77%,相对较高(表4)。
从表4可知,不同种群的多态位点比率在25.32%~53.16%。其中,在10个种源区划中,汤旺河种源区和带岭种源区的遗传多样性最为丰富。五营种源区遗传多态位点比率也达到了41.77%,在本次所收集的种源中多态位点比率较高。长白山2号种源区、新青种源区和双鸭山种源区的多态位点比率最少,分别为25.32%、27.85%、29.11%,证明其遗传丰富度较差。
比较中,小兴安岭种群区域和长白山种群区域的多态位点比率都高于做为两个红松主分布区中间带的双鸭山和牡丹江种群区,证明做为我国国内的红松主分布区,两种群都在长期的进化中保证了种群自身的遗传丰富度。丁庆祝等[16]以5年生红松种源为实验材料,证明了红松不同地理种源间遗传差异是极显著的。

表4 各生态区和种群多态位点统计
2.3 遗传变异分析
Nei指数(H)、Shannon指数(I)和有效等位基因数(Ne)是衡量种源遗传多样性的重要参数。为了分析红松各种群内和种群间的遗传变异,对红松10个种源进行了遗传多样性分析(表5)。
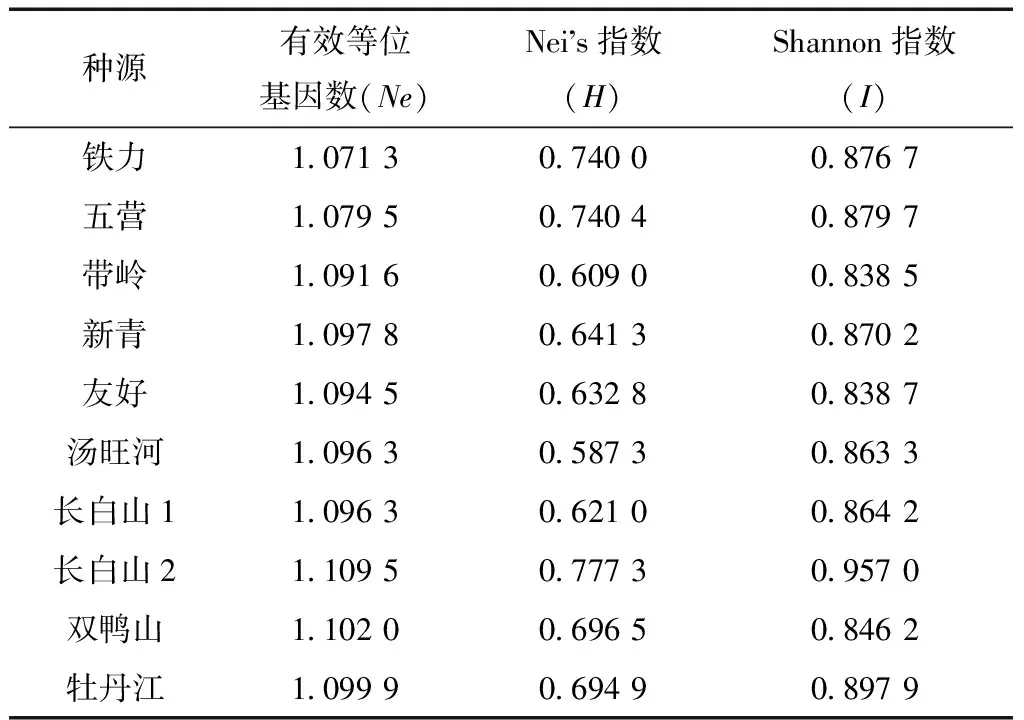
表5 红松各种源遗传多样性
从表5来看,除Shannon指数I比Nei指数H稍高一点外,各种源遗传多样性指数基本一致。其中,带岭种源的Shannon指数I最低,为0.838 5,其次为友好种源。Shannon指数I最高的种源为长白山2号种源和新青种源。结果表明,红松种源Nei遗传多样性指数总体区间在0.403 3~0.874 4。Shannon遗传多样性指数总体区间在0.514 2~1.025 3。各种源总的Nei遗传多样性指数为0.631,总Shannon多样性指数I为0.839 1。
2.4 各种源间的遗传距离和亲缘关系
对红松10个种源的99株单株个体进行聚类分析表明,10个种源基本可以分为两大类群,即包括铁力、带岭、友好、新青、五营、汤旺河等在内的小兴安岭红松群落和包括长白山1号种源、长白山2号种源、牡丹江、双鸭山等在内的长白山红松群落。铁力,友好,新青,五营,带岭、汤旺河等6个种源组内的单株个体在遗传距离上并没有明显的集群分类。
3 结论
(1)对10个种源99株单株优树进行遗传多样性及遗传分化的分析,共检测出有效位点7 821个,其中多态性位点数3 334个。分析结果证明,10个种源基本可以划分为小兴安岭林区、长白山林区和林区过渡带三个大的种源区划。而以双鸭山种源区、牡丹江种源区为代表的过渡带,遗传距离更接近于长白山林区。
(2)试验分析证明,小兴安岭红松区组总扩增位点数为4 661个,多态位点数2 135个。总的多态位点比率为45.81%。长白山红松区组总扩增位点数为1 580个,总多态位点率39.30%。双鸭山和牡丹江种源区组总扩增位点数为1 580个,总多态位点数为576个,总多态位点比率为36.48%。由此看出,无论是各种源组间比较或组内比较,小兴安岭林区总的多态位点比率都相对较高,证明具有更高的遗传丰富度,组内存在更好的基因流动性,可以更为有效的保证种源间基因杂合的丰富度,从而近一步保证林分质量。
(3)对所收集的10个红松种源进行分子标记分析证明,红松各种源的遗传差异在分子层面是显著的。而这种差异的显著性在一定程度上与地理分布关系密切。由此也证明,与其它植物相比,红松种源间的遗传距离受空间限制机率更小,其自身具有较高水平的基因流动。这也是红松在长时间的进化过程中为有效防止基因空间异质性而产生的生理对策。
4 讨论
红松自然分布地域狭长,从小兴安岭到长白山,几千公里,地理及气候条件差异很大,不同亚区的优树子代具有不同的生态适应性[16]。冯富娟等对露水河红松种子园内7个无性系种源进行聚类分析,表明黑龙江的4个种源遗传关系较近,而长白山的3个种源遗传关系同样较近[9],这与本文研究基本相同。但本次研究得到的多态位点比率较高,最终确定的种源区划也稍有差异,这可能是由于试验条件及选择样本方法不同产生的。本次种源选择完全是在红松天然母树林完成,样本树龄从128~444 a不等,共分4个层级进行采集,树龄跨度较大。这一龄级跨度的红松原始林基本未受人为干扰,物种丰富度可以被更好的保留。其次,本次研究选择种源区较多,且每个种源区在采集的过程中都对不同树龄层级的样本进行了采集,样本数的增加增大了位点被检测到的几率。
小兴安岭和长白山红松林地理距离较远,而红松林的群落组成及垂直植被层各有特点,差异较大[17]。张恒庆等在对天然红松林时间尺度遗传多样性变化和遗传分化进行分析后,证明红松在时间尺度具有一定波动[17]。丁庆祝等[18]以5 a生红松种源为实验材料,证明红松不同地理种源间遗传差异是显著的。而做为主分布区之一的小兴安岭红松林群落,由于包括土壤、气候等自然因子和种群内基因流动及杂合程度的复杂性,而形成了自身的群落稳定性。
[1]马建路,庄丽文,陈动,等.红松的地理分布[J].东北林业大学学报,1992,30(9):40-48.
[2]周以良,董世林,聂绍荃.黑龙江树木志[M].哈尔滨:黑龙江科学技术出版社,1986:29-65.
[3]安慧君.阔叶红松林空间结构研究[D].北京:北京林业大学,2003.
[4]郭兴,张 巍,任广明.小兴安岭红松原始林的遗传多样性研究[J].森林工程,2012,28(1):14-16.
[5]张振,张含国,莫迟,等.红松转录组SSR分析及EST-SSR标记开发[J].林业科学,2015,51(8):114-120.
[6]Ge X J,Sun M.Reproductive biology and genetic diversity of a cryptoviviparous mangrove aegiceras corniculatum (Myrsinaceae) using allozyme and intersimple sequence repeat (ISSR) analysis[J].Molecular Ecology,1999,8(12):2061-9.
[7]Prevost A,Wilkinson M J.A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars[J].Theoretical and Applied Genetics,1999,98(1):107-112.
[8]冯富娟,隋心,张冬东.不同种源红松遗传多样性的研究[J].林业科技,2008,38(1):11-4.
[9]冯富娟,王凤友,刘彤.红松ISSR-PCR实验系统影响因素[J].植物学通报,2004,21(3):326-331.
[10]Lester D T.Variation in cone morphology of balsam fir,abies balsamea[J].Rhodorad,1968,70(781):83-94.
[11]Khalil M A K.Genetics of cone morphology in white spruce (Picea glauca)[J].Canadian Journal of Botany,1974,52(1):15-21.
[12]Maley M L,Parker W H.Phenotypic variation in cone and needle characters of Pinus banksiana[J].Canadian Journal of Botany,1993,71(1):43-51.
[13]Parker M H,Maze J.Intraspecific variation in Aies lasioearpa from British Columbia and Washington.Am.J.Bot.[J].Oires,1984,71:1051~1059.
[14]张东冬.SRAP对不同地理种源红松遗传结构的研究[D].哈尔滨:东北林业大学,2008.
[15]马顺兴.日本落叶松无性系遗传变异及早期选择研究[D].郑州:河南农业大学 2006.
[16]徐化成.中国红松天然林[M].北京:中国林业出版社,2000:39-41.
[17]刘得利.天然红松遗传多样性在时间尺度上变化的RAPD和ISSR分析[D].沈阳:辽宁师范大学,2004.
[18]丁庆祝,张彩霞,江淑敏,等.红松不同种源的遗传差异与地理变异趋势[J].林业科技,1994,23(5):1-4
Study on the Genetic Diversity ofPinusKoraiensisin Different Provenances
Zhang Wei,Wang Qingjun,Guo Xing
(Yichun Academy of Forestry sciences,Yichun,153000)
Pinus koraiensis is the main tree species in Xiaoxing'anling forest,and of important ecological status in the overall ecological system of Xiaoxing'anling area.In order to ensure the genetic richness of Pinus koraiensis in Xiaoxing'anling forest region,and provide high quality Pinus koraiensis afforestation seedlings for Xiaoxing’anling forest area,10 primers were screened and 10 different Korean pine natural provenances were analyzed by ISSR in this study.The results showed that the points were detected by 10 ISSR primers were between 2~13loci,DNA fragment length of each point was between 150~2000 bp.79 loci were detected,and 9.9 loci was detected by ISSR primers on the average.Polymorphic bands for the 65 polymorphic loci was 42.63%.There was a significant difference in the molecular level of the various sources,and this difference was positively correlated to the geographical distribution.The genetic richness of Pinus koraiensis community was higher in Xiaoxing'anling forest area,which effectively ensured the quality of Pinus koraiensis community in Xiaoxing'anling.
korean pine;genetic richness;ISSR
2016-10-24
黑龙江省森林工业总局科技攻关项目(sgzjy2010005)
张巍,硕士,工程师。研究方向:森林特产和森林经营。E-mail:zw81025259@163.com
张巍,王清君,郭兴.红松不同种源的遗传多样性分析[J].森林工程,2017,33(2):17-21.
S 791
A
1001-005X(2017)02-0017-05
