鱼类学习记忆能力及其在鱼类驯化养殖中应用的研究进展
2017-05-04曹小欢胡佳宝乐琪君陶顺顺匡思雯徐万土徐芳君王亚军
余 娜, 曹小欢, 胡佳宝, 乐琪君, 陶顺顺, 匡思雯, 张 曼, 徐万土, 徐芳君, 王亚军
鱼类学习记忆能力及其在鱼类驯化养殖中应用的研究进展
余 娜1, 2, 曹小欢1, 2, 胡佳宝1, 2, 乐琪君1, 2, 陶顺顺3, 匡思雯1, 2, 张 曼1, 2, 徐万土3, 徐芳君3, 王亚军1, 2
(1. 宁波大学 应用海洋生物学教育部重点实验室, 浙江 宁波 315211; 2. 宁波大学 海洋生物工程省重点实验室, 浙江 宁波 315211; 3. 象山港湾水产苗种有限公司, 浙江 宁波 315702)
随着鱼类养殖业的发展, 人们对鱼的研究已经不单只涉及形态特征、繁殖习性的层面, 还涉及它们的学习记忆能力。虽然鱼类的大脑结构和组织在某些方面与其他脊椎动物差异较大, 但是它们也能表现出某些高等脊椎动物的大脑功能。作者基于国内外有关鱼类学习记忆能力的研究, 分析了鱼类学习记忆能力的类型, 概括了鱼类接收信息的感官知觉和影响鱼类学习记忆能力的因素, 针对鱼类学习记忆能力在人工驯化技术上的应用提出建议, 并对未来鱼类学习记忆能力在人工驯养技术研究方面的应用提出了展望。
鱼; 感官知觉; 学习记忆能力; 驯化养殖
从20世纪70年代起, 中国渔业资源开始衰退, 人们开始密切关注渔业资源的合理利用与保护, 同时也引发了对鱼类行为的研究热潮。有实验表明, 鱼会习惯于定点投喂, 喂养6个月后, 鱼不仅可以辨认喂食者, 还能在3 min内完成喂食[1]。一个经典的鱼类行为学试验——即通过对鱼进行定点投喂, 来研究鱼类的学习记忆能力, 研究者早晚分别在水族箱的两端投喂, 并且记录鱼的进食位置, 发现鱼在喂食前就已经到了投喂点, 如果鱼可以预先在投喂点聚集, 那么就表明它们已经学会这个任务。花鳉()和江南乳鱼()可以在大约两周的时间内学会这个任务[2]。鳊鱼()和淡水神仙鱼()要花3~4周的时间学会这个任务[3-4]。小鼠()要花19 d的时间来学习这个任务[5]。这些都证明鱼类和其他脊椎动物一样具有学习与记忆的能力。因此, 作者以鱼类学习记忆能力的研究现状为中心, 从鱼类的学习记忆行为、感官知觉、影响鱼类学习记忆能力的因素等方面加以综合评述, 以期对今后鱼类的人工驯化有所启示。
1 鱼类学习与记忆的行为
1.1 条件反射
条件反射是人和动物在个体发育过程中后天获得的, 即出生后通过训练或学习而形成的反射, 是在非条件反射的基础上逐渐形成的, 它具有较大的易变性和适应性。一般, 条件反射又可以分为经典条件反射和操作性条件反射。两种反射有两种明显不同的区别, 经典条件反射不需要条件刺激, 它的形成是源于动物的随意活动, 是欲求行为的结果。操作性条件反射是先接受刺激, 再做出反应, 最后才得到报偿[6]。
鱼类的大脑两半球皮层尚未形成, 产生条件反射的器官在其他部位, 如前脑、中脑等, 这主要取决于刺激的种类, 鱼类可以经过人工驯化和通过在生长环境中获得的经验形成条件反射。但是, 与其他高等脊椎动物相比较, 鱼类的条件反射活动还不完善。即使这样, 鱼类仍表现出较强的学习能力[7]。Hollis等[8]对毛腹鱼()的研究证明经典条件反射能够使鱼对环境建立起积极的适应性。1901年Triplett[9]最先研究鱼类的操作性条件反射, 他以河鲈()为对象, 将河鲈与一些鲤科()小鱼养在一个水族箱内, 中间用玻璃隔开, 开始河鲈有激烈的攻击行为, 但是屡次的失败使得在移开玻璃后, 河鲈也不再攻击小鱼, 说明河鲈已经在多次的尝试中建立起了操作性条件反射。鱼类行为学研究证明了鱼类具有经典条件反射和操作性条件反射能力, 鱼体会随着环境条件的改变而发生适应调整。
1.2 习惯化
习惯化是指对动物反复施加某种刺激后, 其反应强度降低并且最终消失的现象。如果经常对动物进行这种无害的刺激, 动物会逐渐减弱对这种刺激的反应强度, 但这种习惯化往往比较短暂, 如果引起习惯化的刺激长时间不出现, 原来的应激反应就会恢复[7]。例如, 花鳉喜欢在水中上层游泳觅食, 会对水面上的阴影做出逃避反应, 这种反应会随着阴影的反复出现而衰减, 若阴影在较长时间内不再出现, 花鳉的这种习惯化就会消失[10]。另外, 鱼类对光照的反应会随着时间的延长而产生适应性变化, 这种适应性分为暗适应和明适应。在灯光诱捕远东拟沙丁鱼()时, 集鱼时间以30 min最适, 太长或太短均会导致捕捞量下降[11]。在实际围网作业中, 渔船开动主机投网的声音会使鱼群受到惊吓而驱散或下潜, 不易捕捞。为了解决这个问题, 渔船往往会在鱼群的周围绕几圈, 让鱼群习惯这种声音后, 再进行投网捕捞, 充分运用了鱼类对声音的习惯化。这些习惯化所导致的行为都表明鱼类具有一定的学习记忆能力。
1.3 印记学习
印记学习, 指发生在动物生活的早期阶段, 由直接印象形成的学习行为。许多印记学习仅在动物出生后的极短时间内完成, 一旦动物的神经系统在早期接受了这类刺激, 那么在之后的发育过程中, 神经系统便不再进行印记学习了。并且这种学习对后期行为也有影响, 即使不进行任何强化, 也不易消失[12]。鱼类也有印记学习, 其中大马哈鱼()的生殖洄游就是最典型的例子, Sharma等[13]对在海洋和河流生活的大鳞大马哈鱼()的生活史和洄游路线间的关系进行了研究, 他们将大鳞大马哈鱼分成海洋类(在它们出生后的第一年就游到大海中)和河流类的(在河流中生活一年后再游到大海中)两种类型, 不同类型的大鳞大马哈鱼的洄游模式不同, 海洋类的大鳞大马哈鱼在南沿海河流物种中占主导地位, 而河流类的大鳞大马哈鱼主要集中在内部和向北的河流。大鳞大马哈鱼的不同洄游模式, 主要是其发生初期有了对出生河流气味的印记, 因此它一生都不会忘记这种气味, 成熟后的大鳞大马哈鱼回到接近出生河流的沿岸后可依靠出生河流气味的诱导进入出生河流。
2 鱼类的感官知觉对鱼类学习记忆能力的影响
动物的行为反应首先需要接收环境信息, 因此研究鱼类的大脑和行为, 就必须了解鱼类是如何观察到它们周围的环境。鱼类的种类多样而且生活环境复杂, 很难进行归纳。鱼类和人类对世界的感知能力不同, 人们所认为的鱼类的生活环境并非其真正的生活环境, 而是应该依赖于鱼体对环境的感知。因此, 了解鱼类的感官知觉对研究鱼类的学习记忆能力有很重要意义。
2.1 鱼类的视觉
大多数的鱼都有标准脊椎动物的眼睛, 整个眼球近似呈椭圆形, 位于眼眶内, 由眼球壁和内部的折光物质所组成。眼球壁由巩膜、脉络膜和视网膜3层构成, 巩膜有保护眼的作用。巩膜在眼球前方形成透明的角膜, 角膜有折光的作用, 光线透过角膜落到水晶体上。紧贴在巩膜内面的一层为脉络膜, 具有高度的新陈代谢机能, 不但可以供给视网膜营养, 又可防止光的散射[7]。视觉器官构造的不同会影响鱼类的习性, 鱼类视网膜中有两种视细胞, 一种是视柱细胞, 另一种是视锥细胞。视柱细胞对光线的强度敏感[14]。视锥细胞对光线的波长即光照颜色敏感, 主要用以分辨颜色[15]。
Neumeyer[16]研究发现金鱼()的视锥细胞能吸收400、450、530和620 nm的光线。Kamermans等[17]研究发现雀鲷()、慈鲷()、大马哈鱼和金鱼能够看到偏振光, 这种对偏振光的敏感性有助于它们在觅食小的食物时增加对比度, 进行空间定位和视觉传播。有趣的是, 鱼类还知道根据视幻觉来逃跑, 这意味着它们不仅会利用视觉信息, 而且在知觉组织层次上会进行复杂的处理[18]。例如金鱼()和鲨鱼()感知幻觉轮廓的方式与灵长类动物相类似[19-22]。一些鱼眼不仅能感觉光的颜色和明暗, 而且还能感知物体的形状、大小、运动等。雄性鲫鱼()和大马哈鱼在接近产卵期时为了引起雌性个体的注意, 体色会发生变化, 即出现婚姻色, 婚姻色有助于对异性的刺激及认识, 具有催化生殖的功能。在研究雌性剑尾鱼()择偶偏好性时, 发现雌性杂色剑尾鱼更喜欢身体和鳍大的雄性鱼[23-24]。由此可见, 鱼类能够通过视觉以辨别生活环境中各种物体的亮度、颜色和大小、距离、方位、移动情况等, 从而在视觉刺激中获得信息。
2.2 鱼类的化学感觉
与人类的嗅觉和味觉相似, 大多数鱼类也有发达的化学感应能力。它们能够接收并利用化学信息来进行各种活动, 例如觅食、逃避捕食者、择偶和辨认位置等。鱼类的嗅觉感受系统由嗅觉感受器、嗅神经、嗅球组成。嗅上皮的感觉细胞是嗅觉感觉的基本单位[25]。味觉感受器与其他脊椎动物的味蕾相似, 由神经上皮细胞、支持细胞、黏液细胞组成, 神经上皮细胞是味觉感觉细胞[26]。人类有5个主要的味觉(酸、甜、咸、鲜、苦), 而鱼类在生物学上可能只能感受到酸和苦[27]。
Frisch等[28]研究发现幼鱼群中的单个受惊个体就会引起整个鱼群的恐惧感, 他认为是某种或某类化学物质引起了群体性恐惧, 帮助幼鱼躲避危险的捕食者, 一些肉食性鱼类如骨鳔鱼类(Ostariophysi)特殊的皮肤细胞中就有这种物质。当同类受到捕食者的袭击时, 警报危险的物质就会释放到水中, 周围的鱼群就会获得避敌的信息, 并用相应的生物学上的应答方式, 对捕食者的外形进行标记。最近几年的研究表明, 鱼类在胚胎期就能区分捕食者[29]。Nelson等[30]研究发现, 一旦鱼卵开始孵化, 在胚胎发育期就能够将警报危险的物质与捕食者相配对, 这样就能够提高鱼类的避敌能力。
正如上面所提到剑尾鱼的例子, 雌性剑尾鱼对伴侣的选择很慎重, 它们通过一些视觉信息, 来选择潜在的健康雄性(例如大小或颜色)。同时鱼类也能够依靠化学线索找出适合的伴侣。Milinski等[31]对三刺鱼()进行研究, 发现雌性三刺鱼通过化学线索找到的雄性三刺鱼具有更多不同的MHC等位基因(MHC等位基因多样性决定了个体的免疫系统的强弱), 雌性三刺鱼选择最佳的伴侣是为了使它们的后代MHC多样性最优化而不是最大化。Thornhill等[32]对人类进行的一些研究中也发现了类似的结果。
2.3 鱼类的听觉
鱼类对外界环境中声音信息的接收主要靠听觉系统, 鱼类的听觉系统主要由耳、气鳔及其他外周附属结构和听觉中枢组成, 鱼耳是听觉器官的主要部分, 有3个半规管和椭圆囊的前庭管, 包括球状囊和听囊。鱼耳可以感觉平衡, 鱼鳔能起到辅助内耳的作用, 除了传统意义上的听觉系统, 侧线也能感觉到水波的振动, 接受低音和超低音频率, 通过内耳将信息传递到大脑中处理[33]。这使得鱼类对环境因素变化具有高度的敏感性, 也使鱼群之间的联系更加紧密。
一般运用行为学试验来研究鱼类的听觉能力。迄今, 研究发现具有听力能力的物种不到100种。例如鲈鱼()和鲑鱼()可以检测到次声波(35 Hz), 而太平洋鲱鱼()等可以听到180kHz以上的声音, 虽然对鱼类是否能够检测到声音所在的位置一直有争议, 但是已经有证据表明某些鱼类可以对声音进行定位[34]。

2.4 其他的感官
鱼的感觉机制是多种多样的, 其中还有许多人类尚未研究清楚的模式, 其中包括侧线感知、声呐电流、电磁场感应等感知系统。许多鱼类依靠其侧线器官来感知水压的变化, 在水中激发电流[38]。白鲟()和刀鱼()有一种类似于声呐的系统, 这种系统能够发出电流并且检测周围环境的电场变化, 以助于它们觅食、导航和与同伴交流, 这些鱼都会发出电流, 并且能够发现和回应电流。Mouritsen等[39]研究表现, 许多鱼还可以像鸟类一样, 检测到地球的磁场, 并利用磁场进行大范围或小范围领域的导航。鲨鱼和其他软骨鱼有专门的器官来检测磁场的变化。早在120 a前就提出鱼可以探测到地球的磁场并且利用磁场进行洄游的观点, 但直到1980年才证实这个猜想, 但是对于如何利用磁场和磁场感应的作用问题仍有相当大的争论[40]。
3 影响鱼类学习记忆能力的因素
鱼类的生命周期相比于高级脊椎动物来说要短暂, 但是它们所面临的环境却错综复杂, 它们需要探索识别优质的食物资源, 需要躲避敌害的追踪和解决种间的冲突, 以及找到合适的配偶, 这些都要求它们在面临具体情况时能够表现出适当的行为, 而学习与记忆能力在指导鱼类的行为方面扮演着重要角色。不同鱼种间的学习记忆能力具有差异是很常见的, 特别是当鱼群来自不同的生长环境时, 这种差异更加明显。遗传会造成其中的一些差异(这是不同环境下自然选择的结果), 不同鱼类的生活经历也会造成这种差异, 这两种差异形成机制都与鱼类生存的环境条件相关联。总体来说, 鱼类的学习记忆能力与鱼类适应环境, 满足其生存和生殖相一致。
环境直接影响鱼类的生理特征, 进而决定它们的学习记忆能力, 如表1。
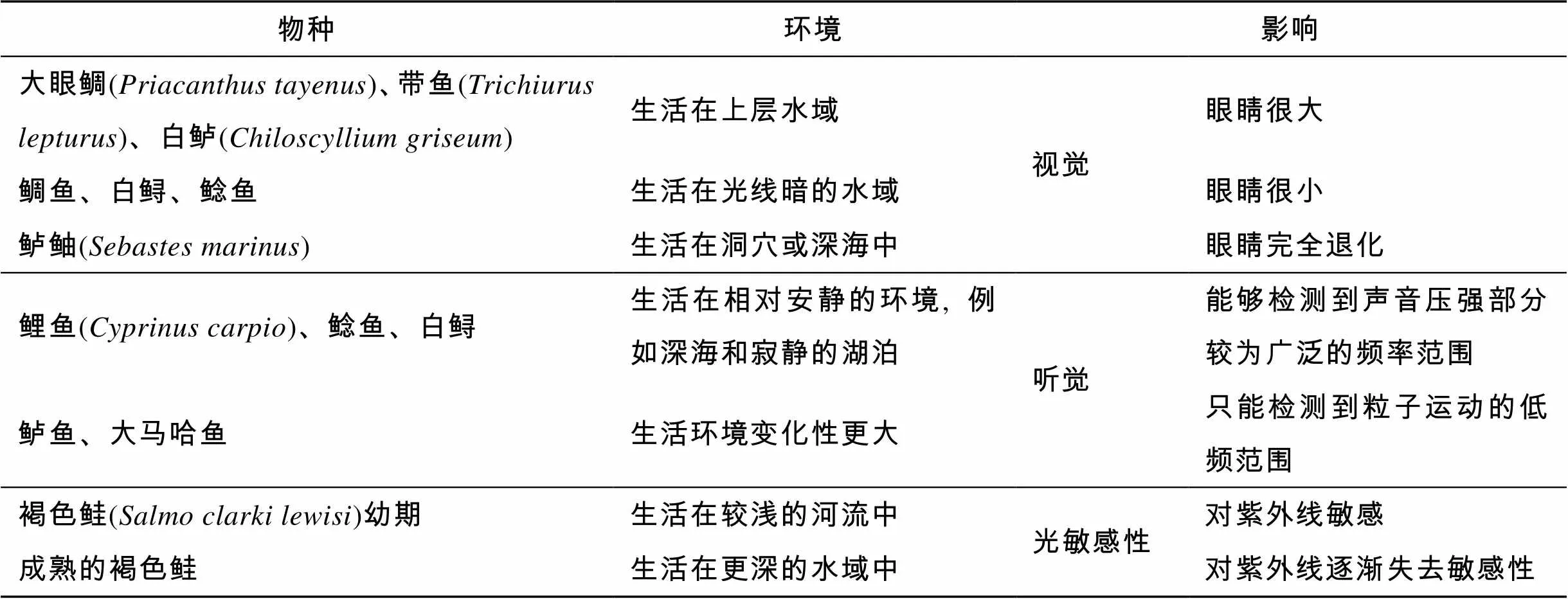
表1 环境对鱼类学习记忆能力的影响
尽管鱼类的生命周期短暂, 但是与其他动物相似, 其生活史对学习记忆能力的形成有着重要的意义[45-47]。有学者专门针对鱼类早期饲养环境进行过研究, 发现高密度的实验室饲养环境能够使鱼类觅食更加灵活, 侵略行为下降, 在恶劣的环境下恢复力更快, 能够提高鱼类的学习能力[48-51]。野生的花鳉比较敏感, 具有较显著的避敌行为, 然而在实验室饲养一段时间后, 来自高密度敌害取得的花鳉, 相对于野生的鱼群来说, 它们避敌行为的强度明显变弱[52]。并且野生花鳉在7 d多的时间内能够显示出快速的学习能力(14 d的试验), 而养殖的花鳉虽然开始的学习能力比较缓慢, 但最终它们能够赶上野生花鳉的学习记忆能力[53]。训练澳大利亚淡水鱼从有洞的网中通过, 只训练5次, 鱼就学会了逃脱, 如果把鱼网撤掉, 11个月后鱼仍然可以很快的完成逃脱任务[54]。训练一群杜氏虹银汉鱼()穿过有洞的网, 只有5尾杜氏虹银汉鱼知道逃脱路线的位置。当试验时隔1 a后, 即使它们在干预期间并没有看到网, 可还是能从中逃脱出来[55]。鱼类能够对不好的经历形成长时间的记忆, 被勾住过的鱼, 在1 a多的时间里还是会对钩子产生恐惧[56]。三刺鱼亲鱼经常会追击幼鱼, 以促使其回巢, 相比于单个生活的三刺鱼, 它们对敌害的应激反应更加激烈[57]。
此外, 同种鱼群中, 不同性别的个体学习记忆能力有差异, 花鳉群体中, 雌鱼所占比例较小, 因此, 鱼群的生殖成功率在很大程度上依赖于雌性的存活率。因此雌性花鳉的学习记忆能力要优于雄性, 这样才能维持种族的延续[58-59]。这与物种种族延续的目的相吻合, 是自然选择的结果。
4 鱼类学习记忆能力在鱼类驯化养殖中的应用
4.1 音响驯化
音响驯化是控制鱼类行为的关键技术之一, 是利用鱼类的听觉, 结合诱食性饵料的适量投喂对其进行条件反射训练, 从而达到控制鱼类行为的目的。在鱼类养殖业中音响驯化技术被广泛使用。日本在20世纪中期, 就已经开始研究音响驯化对真鲷()养殖的影响, 通过建立声音与摄食之间的条件反射, 使鱼记住摄食地点, 以提高饵料的利用率[60]。在欧洲, 文献记载较多的音响驯化对象是大西洋鲑鱼[61]、虹鳟()[61-62]、加利略帚齿非鲫()[63]。Tlusty[61]用50 Hz~400 Hz的音频对大西洋鲑鱼、虹鳟进行驯化, 发现250 Hz时, 刚好能够控制两种鱼摄饵, 大西洋鲑鱼的最大聚集率为85%, 虹鳟的最大聚集率为96%。
4.2 饵料诱食驯化
饵料诱食驯化主要是利用饵料的化学性状对鱼类形成有效的刺激, 鱼类通过嗅觉和味觉器官感受这种刺激, 从而找到食物[64]。中国对饵料诱食驯化的研究甚多, 特别是对配合饵料的研究应用, 极大地提高了中国养殖渔业的经济效益。宋阳[65]等通过对野生肉食性鱼类进行肠胃解剖分析, 以确定鱼的摄食喜好, 为鱼食性驯化的成功提供可靠依据。丁绍雄等[66]在西江斑鳜()的驯养过程中, 将斑鳜驯养分为摄食驯养和食性驯养两个阶段。唐文家等[67]在齐口裂腹鱼()驯化过程中, 混养容易被驯化而且适应能力强的大渡软刺裸裂尻鱼(), 觅食能力强的大渡软刺裸裂尻鱼在3~5 d后就能够主动到固定的饵料台觅食, 齐口裂腹鱼也开始随着大渡软刺裸裂尻鱼进食饵料, 从而极大地加快了驯化速度。但这些试验都是针对池塘、水和网箱等鱼类养殖, 并没有推广到海洋等自然放养环境中。
4.3 灯光干扰

表2 不同诱鱼灯的特征
5 展望
尽管研究人员目前还不能够提供明确的证据解释非人类脊椎动物的意识水平, 但是许多关于鱼类的行为和学习记忆能力的研究均对鱼类的驯化有着重要的意义。目前, 日本和欧美等国主要利用音响驯化的方式, 以实现对鱼群的高效管理。中国的驯化更侧重于通过饵料诱食来改变鱼种的食性。这两种不同层面的驯化方法, 均给养殖渔业带来了良好的经济效益。在工厂化鱼类养殖业中, 更是将光对鱼类学习记忆能力的影响运用到了生产中。但其实根据鱼类感官知觉和学习记忆能力的关系, 电磁场也可用于鱼类的驯化。尽管电诱捕鱼技术已经非常成熟, 但是对电磁场的研究并不深入。因此今后在鱼类养殖驯化方面应结合诱食性饵料, 音响驯化, 声、光诱捕技术开展全方位研究, 以期提高海洋牧场的渔获效率, 建立更加完善的渔场机制, 增加养殖鱼类的生长效益等。
[1] Klausewiz W. Ein bemerkenswerter Zähmungsversuch an freilebenden Fischen[J]. Natur Volk, 1960, 90: 91-96.
[2] Brown C, Warburton K. Social mechanisms enhance escape responses in shoals of rainbowfish,[J]. Environmental Biology of Fishes, 1999, 56(4): 455-459.
[3] Reebs S G. Time-place learning in golden shiners (Pisces: Cyprinidae)[J]. Behavioural Processes, 1996, 36(3): 253- 262.
[4] Gómez-laplaza L M, Morgan E. Time-place learning in the cichlid angelfish,[J]. Behavioural Processes, 2005, 70(2): 177-181.
[5] Means L W, Ginn S R, Arolfo M P, et al. Breakfast in the nook and dinner in the dining room: time-of-day discrimination in rats[J]. Behavioural Processes, 2000, 49(1): 21-33.
[6] 尚玉昌. 动物的经典条件反射和操作条件反射学习行为[J]. 生物学通报, 2005, 40(12): 7-9.
Shang Yuchang. Classical conditioning and conditioned reflex learning behavior in animals[J]. Bulletin of Biology, 2005, 40(12): 7-9.
[7] 何大仁, 蔡厚才. 鱼类行为学[M]. 厦门: 厦门大学出版社, 1998: 12-18.
He Daren, Cai Houcai. Fish behavior[M]. Xiamen: Xiamen University Press, 1998: 12-18.
[8] Hollis K L, Pharr V L, Dumas M J, et al. Classical conditioning provides paternity advantage for territorial male blue gouramis ()[J]. Journal of Comparative Psychology, 1997, 111(3): 219-225.
[9] Triplett N. The educability of the perch[J]. American Journal of Psychology, 1901, 12(3): 354-360.
[10] 郭江涛, 毋全富. 孔雀鱼人工养殖技术[J]. 河南水产, 2009, 4: 19-21.
Guo Jiangtao, Wu Quanfu. Guppy artificial breeding technology[J]. Henan Fisheries, 2009, 4: 19-21.
[11] 罗会明. 光刺激时间与鱼趋光反应变化的关系[J]. 海洋渔业, 1981, 1: 16-17.
Luo Huiming. The relationship between the time of light stimulation and the change of the response of fish to light[J]. Marine Fisheries, 1981, 1: 16-17.
[12] 尚玉昌. 动物的印记学习行为[J]. 生物学通报, 2005, 40(6): 13-15.
Shang Yuchang. Study on the behavior of animal imprinting[J]. Bulletin of Biology, 2005, 40(6): 13-15.
[13] Sharma R, Quinn T P. Linkages between life history type and migration pathways in freshwater and marine environments for[J]. Acta Oecologica, 2012, 41(41): 1-13.
[14] 王子仁, 高岚. 鱼类视神经损伤和再生的研究[J]. 兰州文理学院学报(自然科学版), 1999, 1: 39-45.
Wang Ziren, Gao Lan. Studies on optic nerve crush and regeneration[J]. Journal of Gansu Lianhe University (Natural), 1999, 1: 39-45.
[15] 白艳勤, 陈求稳, 许勇, 等. 光驱诱技术在鱼类保护中的应用[J]. 水生态学杂志, 2013, 34(4): 85-88.
Bai Yanqin, Chen Qiuwen, Xu Yong, et al. The application of light drive and trap technique in fish species protection[J]. Journal of Hydroecology, 2013, 34(4): 85-88.
[16] Neumeyer C. Tetrachromatic color vision in goldfish: evidence from color mixture experiments[J]. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology, 1992, 171(5): 639- 649.
[17] Kamermans M, Hawryshyn C. Teleost polarization vision: how it might work and what it might be good for[J]. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 2011, 366(1565): 742-756.
[18] Agrillo C, Miletto Petrazzini M E, Dadda M. Illusory patterns are fishy for fish, too[J]. Frontiers in Neural Circuits, 2013, 7(5): 137-137.
[19] Wyzisk K, Neumeyer C. Perception of illusory surfaces and contours in goldfish[J]. Visual Neuroscience, 2007, 24(3): 291-298.
[20] Sovrano V A, Bisazza A. Recognition of partly occluded objects by fish[J]. Animal Cognition, 2008, 11(1): 161- 166.
[21] Sovrano V A, Bisazza A. Perception of subjective contours in fish[J]. Perception, 2009, 38(4): 579-590.
[22] Theodora Fuss H B V S. The brain creates illusions not just for us: sharks () can “see the magic” as well[J]. Frontiers in Neural Circuits, 2014, 8(8): 24-41.
[23] Rosenthal G G, Evans C S. Female preference for swords inreflects a bias for large apparent size[J]. Proceedings of the National Academy of Sciences, 1998, 95(8): 4431-4436.
[24] MacLaren R D, Gagnon J, He R. Female bias for enlarged male body and dorsal fins in[J]. Behavioural Processes, 2011, 87(2): 197- 202.
[25] 赵红月, 薛敏, 解绶启. 鱼类的摄食感觉[J]. 水产科技情报, 2010, 37(5): 230-234.
Zhao Hongyue, Xue Min, Xie Shouqi. Feeding sensation of fish[J].Fisheries Science & Technology Information , 2010, 37(5): 230-234.
[26] Fishelson L, Delarea Y. Taste buds on the lips and mouth of some blenniid and gobiid fishes: comparative distribution and morphology[J]. Journal of Fish Biology, 2004, 65(3): 651-665.
[27] Hara T J. Olfaction and gustation in fish: an overview[J]. Acta Physiologica Scandinavica, 1994, 152(2): 207-217.
[28] Frisch K V. Über einen schreckstoff der fischhaut und seine biologische bedeutung[J]. Zeitschrift Für Vergleichende Physiologie, 1942, 29(1-2): 46-145.
[29] Oulton L J, Haviland V, Brown C. Predator recognition in rainbowfish,embryos[J]. Plos One, 2013, 8(10): 271-272.
[30] Nelson A B, Alemadi S D, Wisenden B D. Learned recognition of novel predator odour by convict cichlid embryos[J]. Behavioral Ecology and Sociobiology, 2013, 67(8): 1269-1273.
[31] Milinski M. The function of mate choice in sticklebacks: optimizing MHC genetics[J]. Journal of Fish Biology, 2003, 63(s1): 1-16.
[32] Thornhill R, Gangestad S W, Miller R, et al. Major histocompatibility complex genes, symmetry, and body scent attractiveness in men and women[J]. Behavioral Ecology, 2003, 14(5): 668-678.
[33] 胡运燊, 石小涛, 刘德富, 等. 声音导鱼技术的分析与展望[J]. 水生态学杂志, 2013, 34(4): 89-94.
Hu Yunshen, Shi Xiaotao, Liu Defu, et al. Technology analysis and outlook for the sound guidance fish[J]. Journal of Hydroecology, 2013, 34 (4): 89-94.
[34] Popper A N, Lu Z. Structure-function relationships in fish otolith organs[J]. Fisheries Research, 2000, 46(1): 15-25.
[35] Hawkins A D, Amorim M C P. Spawning sounds of the male haddock,[J]. Environmental Biology of Fishes, 2000, 59(1): 29-41.
[36] Johnston C E, Johnson D L. Sound production in(Rafinesque)(Cyprinidae)[J]. Copeia, 2000, 2: 567-571.
[37] Simpson S D, Meekan M G, Jeffs A G. Attraction of settlement-stage coral reef fishes to reef noise[J]. Marine Ecology Progress, 2004, 276(1): 263-268.
[38] Allen C. Fish cognition and consciousness[J]. Journal of Agricultural & Environmental Ethics, 2013, 26(1): 25-39.
[39] Mouritsen H, Ritz T. Magnetoreception and its use in bird navigation[J]. Current Opinion in Neurobiology, 2005, 15(4): 406-414.
[40] Walker M M, Diebel C E, Haugh C V, et al. Structure and function of the vertebrate magnetic sense[J]. Nature, 1997, 390(6658): 371-376.
[41] Gomahr A, Palzenberger M, Kotrschal K. Density and distribution of external taste buds in cyprinids[J]. Environmental Biology of Fishes, 1992, 33(1-2): 125- 134.
[42] Amoser S, Ladich F. Are hearing sensitivities of freshwater fish adapted to the ambient noise in their habitats[J]. Journal of Experimental Biology, 2005, 208(18): 3533-3542.
[43] Siebeck U E, Parker A N, Sprenger D, et al. A species of reef fish that uses ultraviolet patterns for covert face recognition.[J]. Current Biology Cb, 2010, 20(5): 407- 410.
[44] Deutschlander M E, Greaves D K, Haimberger T J, et al. Functional mapping of ultraviolet photosensitivity during metamorphic transitions in a salmonid fish,[J]. Journal of Experimental Biology, 2001, 204(14): 2401-2413.
[45] Jordan L A, Brooks R C. Recent social history alters male courtship preferences[J]. Evolution, 2012, 66(1): 280-287.
[46] Jordan L A, Kokko H, Kasumovic M. Reproductive foragers: male spiders choose mates by selecting among competitive environments[J]. American Naturalist, 2014, 183(5): 638-649.
[47] Hotta T, Takeyama T, Heg D, et al. The use of multiple sources of social information in contest behavior: testing the social cognitive abilities of a cichlid fish[J]. Frontiers in Ecology & Evolution, 2015, 3(85): 1-9.
[48] Strand D A, Utne-Palm A C, Jakobsen P J, et al. Enrichment promotes learning in fish[J]. Marine Ecology Progress, 2010, 412(6): 273-282.
[49] Lee J S F, Berejikian B A. Effects of the rearing environment on average behaviour and behavioural variation in steelhead[J]. Journal of Fish Biology, 2008, 72(7): 1736-1749.
[50] Salvanes A G V, Moberg O, Braithwaite V A. Effects of early experience on group behaviour in fish[J]. Animal Behaviour, 2007, 74(4): 805-811.
[51] Lee J S F, Berejikian B A. Effects of the rearing environment on average behaviour and behavioural variation in steelhead[J]. Journal of Fish Biology, 2008, 72(7): 1736-1749.
[52] Seghers B H, Magurran A E. Population differences in the schooling behaviour of the Trinidad guppy,: adaptation or constraint[J]. Canadian Journal of Zoology, 1995, 73(6): 1100-1105.
[53] Bibost A L, Brown C. Laterality influences cognitive performance in rainbowfish[J]. Animal Cognition, 2014, 17(5): 1045-1051.
[54] Brown C, Laland K N. Social learning in fishes: a review[J]. Fish and Fisheries, 2003, 4(3): 280-288.
[55] Brown C. Familiarity with the test environment improves escape responses in the crimson spotted rainbowfish,[J]. Animal Cognition, 2001, 4(2): 109-113.
[56] Beukemaj J J. Acquired hook‐avoidance in the pikeL. fished with artificial and natural baits[J]. Journal of Fish Biology, 1970, 2(2): 155-160.
[57] Tulley J J, Huntingford F A. Paternal care and the development of adaptive variation in anti-predator responses in sticklebacks[J]. Animal Behaviour, 1987, 35(5): 1570-1572.
[58] Seghers B H, Magurran A E. Population differences in the schooling behaviour of the Trinidad guppy,: adaptation or constraint[J]. Canadian Journal of Zoology, 1995, 73(6): 1100-1105.
[59] Reader S M, Laland K N. Diffusion of foraging innovations in the guppy[J]. Animal Behaviour, 2000, 60(2): 175-180.
[60] 毕庶万. 日本真鲷的音响驯化[J]. 海洋科学, 1982, 6(1): 58-59. Bi Shuwan. The Japanese sea bream acoustic taming [J]. Marine Sciences, 1982, 6(1): 58-59.
[61] Tlusty M F, Andrew J, Baldwin K, et al. Acoustic conditioning for recall/recapture of escaped Atlantic salmon and rainbow trout[J]. Aquaculture, 2008, 274(1): 57-64.
[62] Abbott R R. Induced aggregation of pond-reared rainbow trout () through acoustic conditioning[J]. Transactions of the American Fisheries Society, 1972, 101(1): 35-43.
[63] Zion B, Barki A, Grinshpon J, et al. Retention of acoustic conditioning in St Peter’s fish[J]. Journal of Fish Biology, 2011, 78(3): 838- 847.
[64] 王德平, 雷洪. 影响鱼类驯食成功的因素[J]. 科学养鱼, 2005, 6: 78-80.
Wang Deping, Lei Hong. Factors affecting the success of fish feeding[J]. Scientific Fish Farming, 2005, 6: 78-80.
[65] 宋阳, 范黎明. 肉食性鱼类食性驯化技术刍议[J]. 渔业现代化, 2002, 6: 18-19.
Song Yang, Fan Liming. Discussion on domestication of carnivorous fish[J]. fishery Modernization, 2002, 6: 18-19.
[66] 丁绍雄, 何美琼, 江林源. 西江斑鳜的驯养技术[J]. 广西水产科技, 2007, 4: 32-35.
Ding Shaoxiong, He Meiqiong, Jiang Linyuan. The domestication technology of[J]. Fisheries Science & Technology of Guangxi, 2007, 4: 32-35.
[67] 唐文家, 杨成. 野生齐口裂腹鱼驯化注意事项[J]. 科学养鱼, 2008, 5: 33.
Tang Wenjia, Yang Cheng. Attention to the domestication of[J]. Scientific Fish Farming, 2008, 5: 33.
[68] 杨吝. 诱鱼灯的原理及实际应用[J]. 江西水产科技, 1995, 3: 34, 41.
Yang Lin. Principle and application of fish lamp[J]. Jiangxi Fishery Sciences and Technology, 1995, 3: 34, 41.
[69] Schluessel V, Bleckmann H. Spatial memory and orientation strategies in the elasmobranch[J]. Journal of Comparative Physiology A Neuroethology Sensory Neural & Behavioral Physiology, 2005, 191(8): 695-706.
Progress in the ability of learning and domesticated breeding of fish
YU Na1, 2, CAO Xiao-huan1, 2, HU Jia-bao1, 2, YUE Qi-jun1, 2, TAO Shun-shun3, KUANG Si-wen1, 2, ZHANG Man1, 2, XU Wan-tu3, XU Fang-jun3, WANG Ya-jun1, 2
(1. Key Laboratory of Applied Marine Biotechnology, Ningbo University, Ningbo 315211, China; 2. Key Laboratory of Marine Biotechnology of Zhejiang Province, Ningbo University, Ningbo 315211, China; 3. The Xiangshan Harbor Aquaculture and Larva Company Limited, Ningbo 315702, China)
The development of aquaculture has led to the research of not only the morphological characteristics and breeding habits of fish but also their learning ability and memory. Despite being considered to exhibit significant differences in some aspects of the brain structure and organization compared to other vertebrates, fish demonstrate functional similarities simultaneously. Based on the current domestic and overseas studies on the learning ability and memory, this study aims to assess the learning ability and memory of fish, discuss sentience of fish, and determine factors that affect the learning and memory of fish. In addition, the study proposes some suggestions and prospects of domestication techniques based on the application of existing learning and memory concepts in the domestication technology.
fish; sentience; learning and memory ability; domestication technology
(本文编辑: 谭雪静)
[National Natural Science Foundation of China, No.31772869; The Project Supported by Zhejiang Provincial Natural Science Foundation ofChina, No.LY15C190003; Ningbo Agricultural Major Project, No.2015C110003; Ningbo Livelihood Key Project, No.2013C11010; Xiangshan County Agricultural Research Project, No.2016C0003]
Apr. 13, 2017
余娜(1993-), 女, 浙江省舟山人, 硕士研究生, 主要从事鱼类功能基因组学研究, 电话: 17858885662, E-mail: eduyuna@163.com;王亚军,通信作者, E-mail: wangyajun@nbu.edu.cn
S965.9
A
1000-3096(2017)12-0155-08
10.11759//hykx20170413002
2017-04-13;
2017-08-29
国家自然科学基金(31772869); 浙江省自然科学基金(LY15C190003); 宁波市农业重大专项(2015C110003); 宁波市民生重点项目(2013C11010); 象山县农业科研攻关项目(2016C0003)
