深海大洋最小含氧带(OMZ)及其生态环境效应
2017-05-04李学刚宋金明袁华茂段丽琴王启栋
李学刚, 宋金明, 袁华茂, 李 宁, 段丽琴, 王启栋
深海大洋最小含氧带(OMZ)及其生态环境效应
李学刚1, 2, 3, 宋金明1, 2, 3, 袁华茂1, 2, 3, 李 宁1, 2, 3, 段丽琴1, 2, 3, 王启栋1
(1. 中国科学院 海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071; 2. 中国科学院大学, 北京 100049; 3. 青岛海洋科学与技术国家实验室 海洋生态与环境科学功能实验室, 山东 青岛 266237)
大洋最小含氧带(Oxygen minimum zone, OMZ)通常是指大洋水体中氧含量缺乏的水层, 一般在水深200~1000 m之间, 其形成主要与厌氧细菌降解有机物导致的溶解氧消耗有关。但到目前为止对OMZ的浓度及水层都没有一个统一的标准, 如以DO低于20 μmol/L为标准, 低于此标准的海域面积可占全球大洋面积的8%, 水体体积可达大洋体积的7%, 分布的主要区域包括东北太平洋(ENP)、东南太平洋(ESP)、阿拉伯海(AS)和孟加拉湾(BB)等海域。OMZ在全球海洋氮循环中有极为重要的作用, 在那里不同化学形态的氮(NH4+、NO2–、NO3–、N2O、N2)在微生物的参与下会发生多种复杂反应。在OMZ的上部混合层, 硝化作用将NH4+转化为NO3–, 但在OMZ核心区主要进行着反硝化作用和厌氧氨氧化作用过程, 可以将NO3–、NO2–和NH4+转化为气态氮(如N2、N2O), 释放到大气中, 造成大洋生物可利用氮含量更加不足。大洋OMZ区的存在不仅影响浮游生物的丰度、分布、多样性, 而且影响生物的生存和迁移行为, 但低氧区并不意味着是生物的贫乏区。同时, OMZ有利于有机质在沉积物中的保存, 影响Fe、Mn、S等对氧化还原环境敏感元素的迁移与转化。尽管已有研究揭示了大洋OMZ对全球物质循环、大洋生态系统和极端环境下生物演化过程有重要作用, 但对大洋OMZ可能带来的复杂生态环境效应的认识仍然很有限, 目前亟须深入探讨大洋OMZ的形成过程及其生态环境效应。
最小含氧带(Oxygen minimum zone, OMZ); 分布与成因; 生态环境效应; 大洋
氧是生物地球化学循环的关键要素, 也是大洋碳、氮循环的主要参与者, 海洋溶解氧含量的变化, 与全球气候系统关系密切。自从Schmidt于1925年第一次报道在太平洋的巴拿马远洋海域存在“氧含量低到没有”的远洋海域以来, 相似条件的海域在东部南北太平洋、阿拉伯海或强度稍低的南大西洋的纳米比亚外海相继被发现[1]。于是Cline和Richards[2]在1972年提出最小含氧带(Oxygen minimum zone, OMZ)的概念, 专指大洋水体中氧含量缺乏的水层。OMZ被认为在整个地质时期具有显著的变化, 很可能改变了大洋携带碳、氮的能力, 因而备受关注。
OMZ在全球氮循环中起着极为重要的作用, 在那里氮的不同化学形态(NH4+、NO2–、NO3–、N2O、N2)在微生物的参与下会发生多种复杂反应。在氧化条件下, 也就是在OMZ的上边界层, 硝化作用将NH4+转化为NO3–, 但在氧含量不足的OMZ区, 反硝化作用和厌氧氨氧化作用可以将NO3–、NO2–和NH4+转化为气态氮(如N2、N2O), 并释放到大气中, 造成大洋氮含量更加不足。特别是OMZ也与影响气候变化的主要气体有关: (i)大洋产生的N2O有50%在这里形成; (ii)形成H2S和CH4, 尤其在OMZ的沉积物界面; (iii)限制大洋吸收大气CO2(直接影响: 有机质矿化的最终产物——CO2的生成。间接影响:因氮的流失限制总初级生产, 从而影响对大气CO2的吸收); (iv)因较高的微生物活性导致DMS的消耗。在化学方面, OMZ还与海水酸化、物质的还原等有关。另外, OMZ还可以看作是孕育生命的初期缺氧的大洋, OMZ区从高到低的不同氧含量的转变可以模拟不同古环境条件下的生物多样性。OMZ可以作为适应低氧环境的特殊生物躲避捕食或其他种竞争的避难所(如巨辫硫菌)在OMZ的下边界甚至成了许多大型生物的栖息地。作为呼吸的屏障, OMZ也与生物的垂直迁移有关(如浮游动物)。在地质历史时期, 海洋缺氧事件与大量物种的灭绝有关(如白垩纪中期)。OMZ对气候和海洋生态系统的潜在影响取决于OMZ的强度。这种强度因各种原因而变化, 如气候变化(因分层导致的交换不畅, O2溶解度的降低)、上升流和河流输入或大气沉降输入的营养盐或金属所导致的自然和人为的施肥等。在当前条件下, 根据最近几十年的观察OMZ的强度一直在增加或加强。
从理论上来说, 低氧区的形成主要有两个基本机制, 一是水体中的溶解氧被不断消耗, 二是被消耗的溶解氧得不到及时的补充。对大洋来说, 消耗溶解氧的过程有很多, 如水体中有机物的降解、无机物的氧化反应、底泥耗氧及浮游植物呼吸作用等; 而氧的补充主要依靠大洋环流, 但目前有关海洋环流供氧和生物地球化学耗氧间可能的相互作用仍不清楚, 预测OMZ的位置、强度和他们的时空变化仍是一个具有挑战性的任务。
由于大洋最小含氧带(OMZ)具有独特的水动力环境和氧含量特征, 其内部的生物活动和生物地球化学循环过程和周围大洋明显不同, 导致大洋最小含氧带的研究越来越受到关注。本文通过对大洋最小含氧带的分布、以及其对海洋碳、氮循环及生态系统的影响进行了系统总结, 以期对大洋最小含氧带的生态环境效应有一个全面的认识。
1 大洋最小含氧带的分布特征与成因
1.1 大洋最小含氧带的分布
大洋最小含氧带(Oxygen minimum zone, OMZ)通常是指大洋水体中溶解氧含量不足的水层, 但到目前为止还没有一个统一的标准值来确定OMZ。常用的描述低氧环境的名词有多种, 如亚氧化(Suboxia)是生物学家和生物地球化学家用来定义氧化物从O2到NO3–的转换层, 阈值在0.7~20 μmol/kg之间。缺氧(Hypoxia)是指大型生物不能生存的含氧环境(8 μmol/kg), 但依据的生物不同最高可到40 μmol/kg, 如鳀鱼。贫氧(Dysoxia,O2<4 μmol/kg) 和微氧(microxia, O2<1 μmol/kg) 主要与大型生物, 如鱼类的氧需求有关。无氧(Anoxia, (O2<0.1 μmol/kg)被定义为由NO3–向硫酸盐被还原的转换的条件。不同的研究者往往由于研究的目的不同而选用不同的标准, 所以在文献中描述的每一个OMZ的标准可能是不同的, 例如: Helly和 Levin[3]在研究全球陆架边缘海低氧区时采用的临界值是20 μmol/kg (0.5 ml/L), Karstensen等[4]在研究大西洋和太平洋最小含氧带时使用的标准是低氧(suboxic level, 4.5 μmol/kg), 稍严格的标准(45 μmol/kg)和比较宽松的标准(90 μmol/kg)。
根据the World Ocean Atlas 2001(WOA01)调查数据, 全球大洋海水中溶解氧的含量分布如图1所示,其中溶解氧含量小于20 μmol/kg的水体占全球海洋体积的1%, 而≤10 μmol/kg的水体占比<0.3%。但是Fuenzalida等[5]认为这个数值被严重低估了, 估值被低估了约90%。造成这种结果的原因之一就是数据量太少[6]。
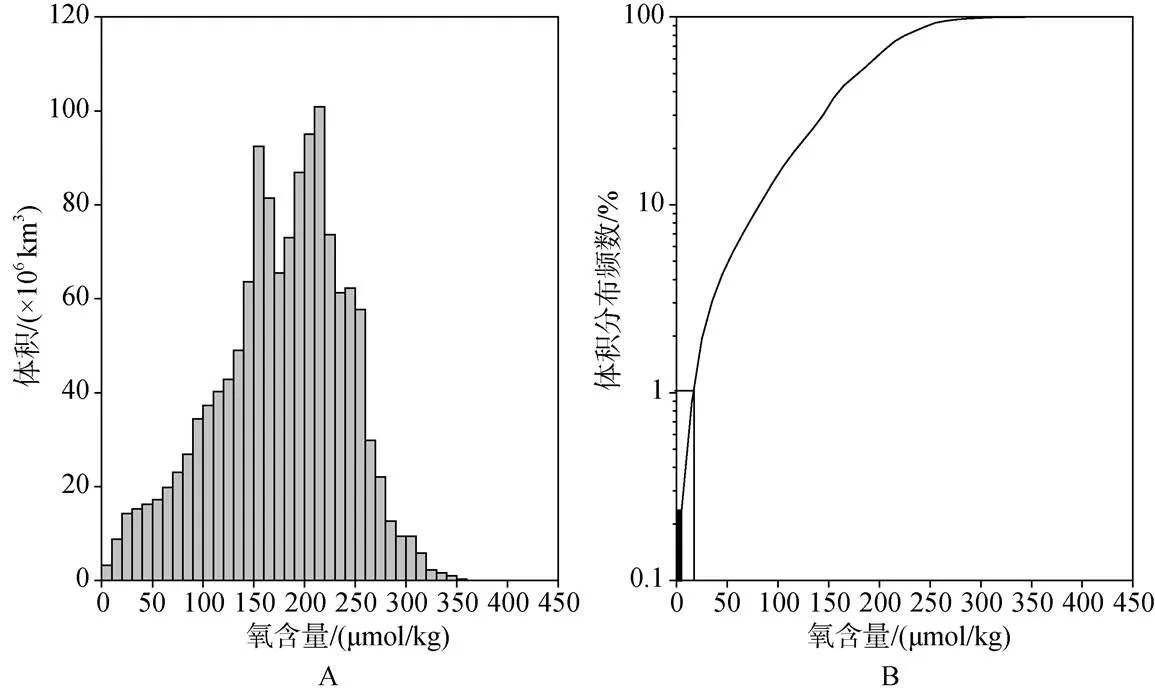
图1 全球大洋中溶解洋含量的分布[6]
A: 体积分布; B: 体积分布频数; 黑色方块代表溶解氧含量≤10 μmol/kg的海水体积百分比; 白色方块代表溶解氧含量≤20 μmol/kg的海水体积百分比
A: The volume Distribution of the dissolved oxygen content; B: Cumulative volume frequency for the oxygen concentration. The black box represents the percentage of waters with≤10 μmol/kg dissolved oxygen and the white box the percentage of waters with ≤20 μmol/kg.
Paulmier和Ruiz-Pino[2]根据WOA2005获得的1894–2004年间全球大洋溶解氧数据绘制了全球OMZ分布图(图2), 可以看到大洋最小含氧带普遍存在, 但以东北太平洋(ENP, Eastern North Pacific)、东南太平洋(ESP, Eastern South Pacific)、阿拉伯海(AS, Arabian Sea) 和孟加拉湾(BB, Bay of Bengal)含量最低。为O2含量低于20 μmol/kg的低氧区的面积可占全球大洋面积的8%, 水体体积可占大洋体积的7%。
图2 O2含量最小层中氧的分布[2]
Fig. 2 O2distribution (μmol/kg) at depth where O2concentration is minimal[2]
比例尺棒中, 在0~20 μmol/kg的间隔为1 μmol/kg± 2 μmol/kg; 在30~340 μmol/kg的间隔为20 μmol/kg ± 2 μmol/kg。等深线是OMZ核心区深度, 间隔为100 m; AS: 阿拉伯海; BB: 孟加拉湾; BLS: 黑海; BS: 巴拉提克海; ESP东南太平洋; ENP: 东北太平洋; ESTNP: 热带北太平洋东南部; ETNP: 热带北太平洋东部; GA: 阿拉斯加湾; PG: 波斯湾; RS: 红海; SWACM: 西南非洲陆架边缘; WBS: 白令海西部
The color barscale corresponds to a 1 μmol/kg ± 2 μmol/kg interval between 0 and 20 μmol/ kg, and a 20 μmol/kg ± 2 μmol/kg interval between 20 and 340 μmol/kg. The isolines indicate the limit of the upper OMZ coredepth in meters with a 100-m contour interval. AS: Arabian Sea; BB: Bay of Bengal; BlS: Black Sea; BS: Baltic Sea; ESP: Eastern South Pacific; ENP: Eastern North Pacific; ESTNP: eastern subtropical North Pacific; ETNP: Eastern Tropical North Pacific; GA: Gulf of Alaska; PG: Persian Gulf; RS: Red Sea; SWACM: Southwest African continental margin; WBS: western Bering Sea.
特别是Karstensen等[4]的研究也证实, 在大西洋和太平洋的东赤道海域的100~900 m 深度广泛分布着最小含氧带(OMZ)。在东太平洋的300~500 m深度溶解氧含量达到最小值, 可低于1 μmol/kg, 形成亚氧化环境(suboxic, DO<4.5 μmol/kg)。在东大西洋不会形成亚氧化环境, 具有相对较高的最小溶解氧值; 在南大西洋约为17 μmol/kg, 在北大西洋约高于40 μmol/kg。当以45 μmol/kg(或90 μmol/kg)为OMZ溶解氧含量的上限时, 大约20%(40%) 的北太平洋水体为OMZ海域, 但在南太平洋OMZ的面积相对较小, 相对占比不到北太平洋的一半, 仅有7%(13%);南大西洋的OMZ区域占总海域的1%(7%), 北大西洋只有约0%(5%)。
根据最近几十年的观察, OMZ的强度在近年来一直在增加或加强。而根据“未来海洋”模型的预测, 在全球变暖条件下溶解氧的含量会降低, OMZ区域会扩展。但在季节或年际间仍然会有较大的波动, 如Helly和Levin[3]的调查发现, 在1997–1998年的厄尔尼诺期间, 秘鲁和智利北部外海的东南太平洋OMZ区域(<20 μmol/kg)的面积缩减了60%。至于产生这种波动的原因仍不清楚。
大洋最小含氧带(OMZ)根据其氧含量的垂直变化特征可以划分为3个层: 上部氧跃层(UO, 氧含量由正常急剧减少)、核心层(OMZ, 氧含量最低)、下部氧跃层(LO, 氧含量从最低逐渐恢复)(图3)。在OMZ的上方是混合层(ML, 氧含量较高), 下方是亚氧跃层(SO, 氧含量继续升高)。其中, 上部氧跃层被认为是OMZ的引擎, 大部分高强度的矿化在那里发生, 导致OMZ增强, 同时也可发生硝化作用和反硝化作用; 在OMZ核心区, 反硝化作用等典型的厌氧过程在这里发生; 在下部氧跃层, 硝化作用是其主要过程, 总之, 在OMZ的不同区域因氧化还原的能力不同, 致使发生的地球化学反应不同。但Köllner等[7]在热带北大西洋氧最小带进行的示踪扩散实验发现, OMZ核心区域氧的扩散速率和氧跃层区域并没有明显的降低。
1.2 OMZ形成的原因
大洋中最小含氧带(OMZ)一般在水深200~1000 m, 其在大洋的形成主要与厌氧细菌降解有机物导致氧的消耗, 而所消耗的氧又得不到及时补充有关。但不同海域导致氧消耗和补充的具体过程又有所不同。
在热带海域, 最小含氧带(OMZ)主要分布在水交换不畅的区域。在印度洋OMZ主要分布在阿拉伯海和孟加拉湾, 那里水交换时间为30年或更长。阿拉伯海OMZ(ASOMZ)是世界上热带海域第二大的OMZ, 主要分布在200~1000 m水深, 氧含量低于5 μmol/kg, 远低于发生反硝化作用发生的含量, 氧含量最低的区域对应于初级生产力最高的区域。孟加拉湾OMZ (BBOMZ)的强度小于阿拉伯海OMZ, 氧含量高于发生反硝化作用的水平。次表层水如果没有氧的补充, 沉降到次表层水中有机碎屑的持续分解将消耗光水层中所有的氧。阿拉伯海和孟加拉湾中的氧主要来自南半球, 沿南极绕极流北部边缘形成的高溶解氧的中层水贯穿整个印度洋。其他来源包括来自波斯湾和红海的高氧水, 波斯湾水可进入阿拉伯海温跃层的下部, 而红海水可到达中层(300~1000 m)。另外, 印度尼西亚贯穿流也影响印度洋的上层水, 包括影响阿拉伯海的温跃层特征, 中尺度涡也可能促进氧的交换与扩散[9]。除了位于水交换不畅的区域外, 到目前为止对保持印度洋OMZ的关键过程仍没有定论。如是什么物理和生物过程决定了低氧区的强度和结构?环流和涡流在高氧水的扩散和混合过程中起什么作用?
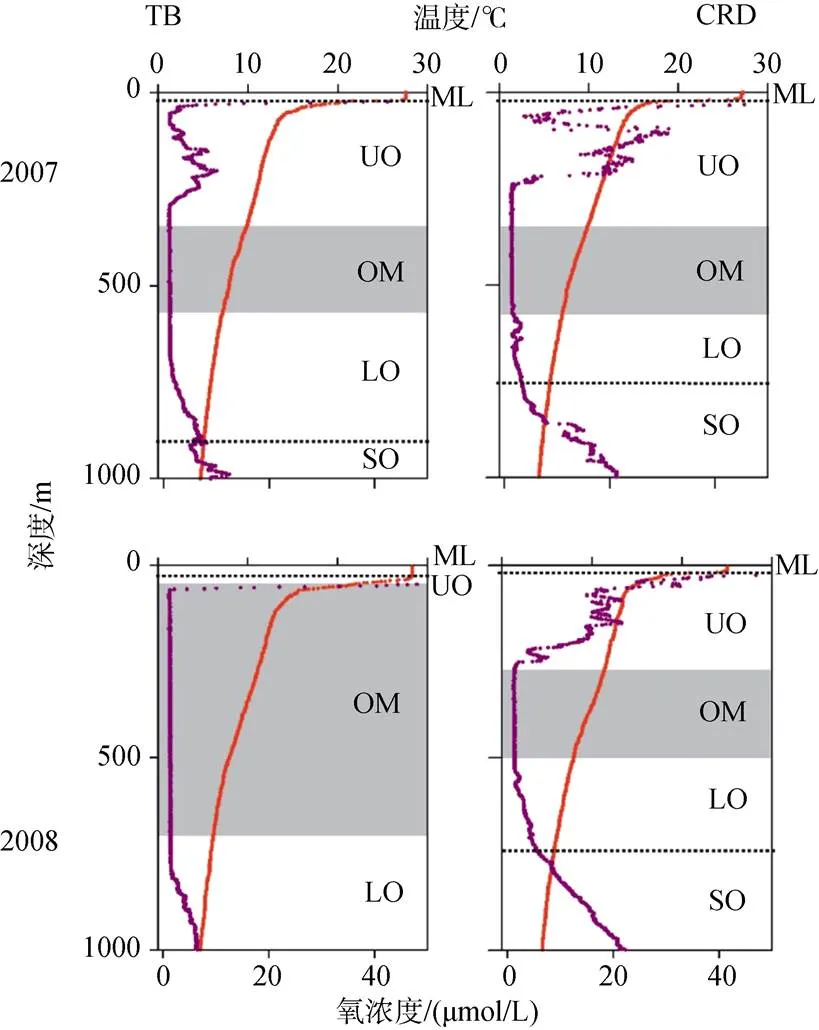
图3 OMZ的垂直分区[8]
红色曲线为温度曲线, 紫色曲线为溶解氧曲线; ML混合层、UO上部氧跃层、OM 核心低氧带、LO下部氧跃层、SO亚氧跃层
The red lines indicate the temperature, the purple lines indicate the dissolved oxygen; ML: mixed layer; UO: Upper oxycline; OM: OMZ core; LO: Lower oxycline; SO: Suboxycline
除了水文条件外, 生物是OMZ形成的关键因素之一。当地有机质的微生物降解是OMZ溶解氧消耗的主要因素, 而浮游动物的呼吸也是一个因素。按平均消耗量, 有机质的微生物降解消耗约80%的氧, 浮游动物呼吸消耗约20%的氧[10]。
2 大洋最小含氧带的生态环境效应
2.1 大洋最小含氧带对氮循环的影响
除了溶解氧浓度低外, OMZ的另一个特征是相对于无机磷来说更加缺乏无机氮。在表层形成的有相对固定氮磷比(Redfield比)的有机质沉降和矿化后, 释放出相同比例的N和P, 所以大洋中氮磷浓度的摩尔比接近有机质中的比值(16︰1)。但是, 在OMZ区, 通过异养反硝化作用和厌氧氨氧化作用生成N2而逸出, 导致无机氮(主要是NO2–+NO3–)显著偏离Redfield比, 造成氮缺乏。
OMZ在全球氮循环中起着关键作用, 在那里氮的多种化学形态(NH4+、NO2–、NO3–、N2O、N2)在微生物的参与下会发生多种复杂反应。在氧化条件下, 也就是在OMZ的上部混合层, 硝化作用将NH4+转化为NO3–, 但是OMZ主要与反硝化作用有关。反硝化作用将NO3–转化为气态氮(如N2、N2O), 可以释放到大气中, 造成大洋氮含量更加不足。但最近发现的厌氧氨氧化过程, 在OMZ区无论是在沉积物还是水体中都可以将NO2–和NH4+转化为氮气。Farias等[11]研究了OMZ对N2O循环过程的影响, 发现水体中N2O和NO3–的最大值位置与紧邻真光层的强烈的氧跃层有关。NO2–在氧跃层下部积累, 可以达到9 μmol/L。在氧跃层的上部即混合层底部和氧跃层的中间点之间, O2从250 μmol/L下降到50 μmol/L, O2和NO3–都有较大的浓度梯度(二者是反向的), 表明在真光层中的硝化作用产生N2O和NO3–; 在氧跃层的下部即OMZ的上边界区, O2从50 μmol/kg下降到11 μmol/kg, NO3–开始被还原, 会生成约41%~68%的N2O, 在这一层N2O循环中涉及硝化作用和NO3–的还原作用。在OMZ核心区, O2的浓度比较稳定(O2<11 μmol/kg), 在这一层N2O和NO3–持续减少, 但生成大量的NO2–。
厌氧氨氧化是氨被NO2–或NO3–厌氧氧化为N2的过程, 在氧缺乏体系中会导致氮的流失。厌氧氨氧化是被一组属于浮霉菌门的特殊菌完成的, 但目前对厌氧氨氧化菌的分布、活动性和控制因素了解不多[12]。Galan等[12]公布了智利北部海域厌氧氨氧化菌的系统进化的多样性、垂直分布和活动性特征, 该海域是第一个被报道的东太平洋OMZ。他们发现厌氧氨氧化菌的微观多样性比早前报道的高, 他们描述了一个新的代表性的海洋微生物系统发生世系。在秘鲁和阿拉伯海也有类似的报道。Galan等[12]也定量报道了这些细菌的丰度、垂直分布和活性, 在OMZ的上层部分达到一个峰值, 可能与这一层强烈的生物地球化学循环有关。
在厌氧氨氧化发现之前, 氨的生物氧化被认为是在硝化过程中被厌氧氨氧化菌专门发生的。然而, 因为这一过程需要氧, 所以被认为是在氧跃层而不是在OMZ核心区发生的硝化作用。但分子学研究显示氨氧化细菌基因在表层氧化性水体和OMZ海域都存在。Molina和Farias[13]发现, 无论是在氧跃层还是在OMZ核心区的硝化速率, 都足以满足氨氧化和二氧化碳暗固定(dark carbon fixation)的需要。Molina和Farĺas[13]对智利北部热带南太平洋低氧区的研究发现, 在核心低氧区, 有氧氨氧化速率高于低氧区上部的氧跃层, 通过有氧氨氧化固定的碳占弱光层总固碳量的33%~57%, NO2–在核心低氧区和其上部的氧跃层消耗较高, 但有氧氨氧化可以贡献8%~ 76%的NO2–产量, 也就是说对反硝化作用贡献较大。总之, 有氧氨氧化在低氧带及其上部边界层中的氮和碳循环中起着重要的作用。
虽然厌氧氨氧化作用对氮损失贡献的确很大, 但反硝化作用仍然是低氧区氮损失的重要途径, 在一些区域内反硝化作用仍占主导地位。Dalsgaard等[14]利用15N 示踪法研究了南太平洋东部的智利和秘鲁沿岸海域的N2O和N2的来源, 发现72%的N2是由反硝化作用产生的。Mashifane等[15]对非洲西南沿岸本格拉OMZ区的研究认为, 硝化作用和厌氧氨氧化、反硝化作用对本格拉海域的高氮通量的贡献相当。
Bohlen 等[16]的研究发现, 在秘鲁陆架和上陆坡(水深80~260 m, 低氧带)沉积物中存在大面积的细菌席, 硝酸盐异化还原成氨(DNRA)在总氮的转化中占主导地位, 大约有≥65%的NO3–+NO2–被沉积物吸收, 但沉积物并不是溶解无机氮(NO3–+NO2–+NH4+)的汇, 因为DNRA将NO3–, 甚至部分NO2–还原为NH4+, 因此, 由于相对低的反硝化速率和较高的DNRA释放氨的速率和有机质氨化速率, 陆架和上陆坡沉积物成为DIN的再循环场所。这一发现与目前的主流观点——OMZ下覆沉积物是DIN的强汇相反。在这里, 反硝化作用是主要过程, 可移除占沉积物中55%~73%的NO3–和NO2–, DNRA和厌氧氨氧化占剩余的部分。厌氧氨氧化在陆架和陆坡并不算重要, 但在水深1 000 m处可贡献总生成氮的62%。这一研究结果表明海洋沉积物是DIN的汇还是再循环场所主要由氧化态氮(NO3–, NO2–)进行DNRA和反硝化过程来控制, 所以在低氧带氧化态氮的底层吸收速率对海洋环境中氮的亏损并不是必要的。
世界大洋中的最小含氧带(OMZ)具有强烈的厌氧氮的转化, 如最大的OMZ——阿拉伯海最小含氧带中氮损失的30%~50%为氮的厌氧转化。在氧含量很低的水体中通过N2生产而造成氮的流失被认为是造成过量氮气和硝酸盐匮乏的主要原因。通过添加15NH4+、15NO2–、46N2O的对阿拉伯海OMZ中的海水进行示踪培养实验, 发现反硝化作用速率与海水中NO2–浓度密切相关, 向海水中添加有机质和氨并不能促进反硝化速率, 反硝化作用是阿拉伯海OMZ产生N2的主要作用[17]。
De Pol-Holz等[18]分析了沉积物和水体中的N稳定同位素, 发现沉积物中的δ15N具有强烈的纬度效应, 其沿智利海岸向北不断增加。他们认为智利沿岸沉积物中的δ15N在41°S以南主要由浮游植物的同化作用控制, 在30°S以北主要由反硝化作用控制。Schwartz等[19]测定了巴基斯坦边缘海OMZ的沉积反硝化作用, 发现在整个阿拉伯海OMZ和紧邻海域(如400 m和1 200 m)内沉积反硝化作用是一个持续过程。在西南季风期间沉积反硝化作用加强。该区域沉积反硝化作用速率与其他陆架环境相当, 但高于深海。整个巴基斯坦边缘海OMZ的沉积反硝化通量可达1.1~10.5 Tg/a。
OMZ无论在水体还是沉积物都是氮流失的关键区域。Sommer等[20]发现秘鲁(OMZ)的O2含量在约500 m处最低, 氧含量在检测限以下。这里底部氮循环存在明显的不平衡, 沉积物释放出大量的NH4+, 远远超过氨化所形成的NH4+, 他们认为这是硝酸盐异化还原成铵(DNRA)造成的。秘鲁OMZ中的底部氮、硫循环表现出特别易受底层水中O2、NO3–和NO2–波动的影响, 当NO3–和NO2–被消耗后将加强低氧区的作用。
OMZ被认为是温室气体的强源N2O和/或CO2潜在的源。Paulmier等[21]对智利东南太平洋OMZ(大洋中强度最大的浅水OMZ之一)中N2O和CO2的海气界面通量进行了估算, 发现智利OMZ是大洋强的N2O和CO2的源。N2O的平均通量是太平洋和印度洋开放大洋区域OMZ的5~10倍。高温室气体源区与近岸富含N2O和CO2的上升流有关。
热带东南太平洋具有强的近岸上升流、狭窄的大陆架和广泛分布的中层水最小含氧带(OMZ)。这种水动力特征决定了中层水物质向表层的长久输送, 使富含营养盐的表层水具有较低的无机N︰P。为了研究影响上升流的OMZ对浮游植物生长、元素和生物组成的影响, Franz等[22]研究了从狭窄大陆架上部的上升流区到具有良好分层的大洋东部边界的广大东南热带太平洋区域中的水动力和生物地球化学参数, 沿岸上升流区的新生产力主要由大型浮游植物(如硅藻)贡献, 他们通常具有较低的N︰P(<16︰1)。在整个海域硝酸盐和磷酸盐并不限制浮游植物的生长, 但在远岸海域, 浮游植物的生长受硅限制。位于陆架坡折处上方次表层水处的温跃层叶绿素丰度最高, 与微型-微微型(聚球藻、鞭毛虫)和小型浮游植物的分布一致, 在温跃层N︰P比接近Redfield比值。在分层的宽阔大洋具有高的PON︰POP(20︰1), 与微型蓝细菌——原氯球藻的丰度一致。过量磷酸盐的存在并没有促进固氮浮游植物的生长, 在OMZ内生成的过量磷酸盐中, 大部分是被表层水中非Redfield比值的大型浮游植物消耗的。
2.2 OMZ对有机质降解的影响
虽然氧含量是影响有机质的降解的主要因素, 但在OMZ区有机质的降解过程中氧的具体作用仍不清楚。有机质的厌氧降解通常被认为比微生物有氧降解缓慢, 这是因为在陆架边缘缺氧区下面的有机质含量比较丰富, 同时厌氧过程产生的能量少于有氧过程。为了估算有氧和低氧条件下活性有机质的降解动力学过程, Pantoja等[23]利用沉积物捕集器研究了智利北部OMZ区颗粒有机碳的降解特征, 发现绝大部分(82%)光合作用产生的蛋白质在上部30 m内降解, 另外15%在30~300 m (弱氧化层内)降解, 只有大约1%能到达1 200 m深的表层沉积物。沉降的蛋白质在上部300 m仍保持着降解活性, 可以作为新鲜物质的降解指数和降解速率常数的指示指标。他们认为颗粒蛋白质的降解不受30~300 m间的弱氧化层存在的影响, 这一结论与颗粒蛋白质的降解受细胞外的蛋白水解酶控制而不依赖于氧的模式一致。活性溶解有机质在智利氧化和亚氧化海水中具有相似的微生物降解速率。肽水解和氨基酸吸收速率在氧被消耗的水体中并不低, 水解速率在上部20 m为65~160 nmol/(L·h), 在100~300 m(氧消耗区)为8~28 nmol/(L·h), 在600~800 m为14~19 nmol/(L·h);在同一深度溶解自由氨基酸吸收率分别为9~26、3~17和6 nmol/(L·h)[24]。这一结果与墨西哥湾浮游植物产生的活性物质的降解速率一致, 说明在高或低溶解氧含量水体中降解速率的不同可能是底物活动性不同造成的。对比水解和吸收速率的不同可以发现微生物肽降解的速率与氨基酸吸收速率接近或快一些, 肽水解不是活动大分子物质完成再矿化的限制速率。低氧水每小时可以处理大约10 t肽, 在表层氧水体中可以处理双倍的肽。在OMZ, 上升到表层的溶解有机质的低溶解度可能影响碳平衡。在受洪堡流系影响的高生产力的智利沿岸大洋的一个显著特征就是存在OMZ, 溶解有机质的很大一部分在OMZ被处理。浮游动物进出OMZ是将大量新生产的碳输入OMZ区最有效的机制[25]。Espinosa等[26]对智利水体悬浮颗粒物中来自脂肪酸甲酯的细菌磷脂的分析发现, Antofagasta外海水中脂肪酸甲酯的垂直分布不能说明微生物活性的隔离区, 可能因为风生沿岸上升流改变了环境特征, 使不同微生物群落混合在一起。智利北部水体中脂质生物标志物的垂直分布说明硫酸盐还原菌产生的脂质生物标志物不仅仅限于OMZ。这种混合可能是深部富营养盐及CO2饱和的冷水传输造成的垂直混合的结果。
海水中的颗粒有机碳沉降到海底后会继续发生着各种变化, 对其积累/保存进行的研究发现, OMZ区氧浓度与沉积物中有机质的保存密切相关。在研究较多的阿拉伯海OMZ区, 中陆坡沉积物受OMZ的影响使有机碳得以更好地沉积, 从而使这一区域成为世界大洋沉积有机碳含量最高的区域[27]。在水深300~1000 m的陡峭陆坡的海底, 因缺乏大型底栖生物、游泳生物, 故无生物扰动, 有机碳含量高、沉积层序良好等都说明该区域为近无氧条件[28]。位于低氧带内比低氧带外的沉积物具有显著低的氧化还原环境、高的碳酸盐含量、以及不同的岩土特征和不同的生物扰动特征, 而且这些差异与低氧程度和水深有关。在低氧带内, Eh、pH和碳酸盐含量随水深而增加, 而总有机质(TOM)和含水率随水深降低[29]。Weijden等[30]认为OMZ内部和OMZ下方沉积物中有机质的生物和微生物过程不同而导致二者存在有机质保存的差异。在氧含量较高的条件下, 生物扰动和活性Fe、Mn氧化物均是影响微生物介导的有机质降解的重要因素。而阿拉伯海OMZ下方的沉积物在低溶解氧条件下, 不仅减少了有机质的好氧矿化作用, 而且减少了底栖生物活动引起的OM降解作用和微生物利用Mn、Fe氧化物作为电子受体的低氧矿化作用。Vandewiele等[31]发现OMZ沉积物中有机碳含量和有机碳与矿物表面积的比值(OC/SA)在核心低氧区和低氧区下转换区明显升高, 而且这两个区域的沉积物含有更多的活性有机质, 比如含有更高浓度的水解性氨基酸(THAA), THAA中的氮对总氮的贡献更大。虽然沉积物中的有机质都发生了矿化, 但低氧区上部转换区和低氧区下部沉积物中的有机质分解更多。
2.3 OMZ对金属迁移的影响
OMZ的生态环境可由氧化性变为弱氧化性、甚至是无氧状态的还原性, 而对氧化还原环境最为敏感的元素, 如Fe、Mn、S等受OMZ的影响最为显著。例如, 东热带南太平洋海域具有较高的固氮速率, 但表层Fe常常太低不足以支撑固氮生物的生长, Kondo等[32]发现该区域OMZ内的Fe(II), 约占溶解铁的8%~68%, OMZ在Fe向上层400 m水体的输送过程中起着重要作用。在阿拉伯海的OMZ带没有检测到游离的H2S, 但检测到了金属硫化物。相对于表层海水, OMZ带内的硫化物含量稍高, 可能与有机质降解释放出硫有关。在低浓度的金属和S2–条件下, 硫化合物的存在形式有两种。首先, 金属如Mn、Fe、Co、Ni形成硫氢化物(HS–), 活动性强, 易于参加反应; 其次, 像Cu、Zn一类的金属与S2–形成多核化合物, 可以从稳定到易于分解, 比S2–(HS–)和活动的金属硫氢化物的活动性弱。Zn、Cu硫化物对低氧海水中H2S的存在至关重要[33]。
海水中的锰的地球化学特征受其氧化还原特性控制, 其中Mn(II)可溶于水, 而Mn(IV)形式不溶于水, 但二者通过一系列的氧化还原过程可以相互转化, 溶解Mn在还原条件下可以聚集而在氧化环境下被消耗。在秘鲁上升流区——以强还原条件下宽广的大陆架和远离海岸的强最小含氧带为特征, 是热带南太平洋东部潜在的最大锰的来源。但Vedamati等[34]对秘鲁上升流和OMZ中锰、氧和氮循环的研究发现令人诧异, 即溶解锰的消耗是在秘鲁大陆架较为还原的条件下进行的, 溶解锰的含量在近岸表层水中增加, 说明来自秘鲁大陆架海水的水平运移不是锰的主要来源, 在氧跃层而不是OMZ带内的次表层水中观察到锰的最大含量, 说明锰来自有机质的矿化释放, 而不是锰氧化物的还原。大陆架水体中锰含量比预期的低说明从陆源河流输入的锰是有限的, 其行为与铁明显不同, 铁在陆架水体中含量较高而且与氧化还原过程密切相关。Lewis和Luther III[35]发现阿拉伯海的OMZ强烈影响了锰的分布。在OMZ, 可以观察到两个明显的溶解锰最大值, 分别在200~ 300 m 和600 m。后者的峰值大约可以达到6 nmol/L。这个中等深度的最大值与OMZ的低氧核心([O2] <2 μmol/kg)有关。上部溶解锰最大值(200~300 m深)比600 m处的峰值分布宽的多, 溶解锰的浓度在3~ 8 nmol/L, 与NO2–的次最大值有关, 都处于反硝化区。雾状层也与这个深度有关。颗粒锰在相同的深度也表现出类似的最小浓度、低Mn/Al和活性锰/难溶锰比值, 表明锰氢氧化物的还原性溶解。所有的观测都表明原位微生物降解过程可能是OMZ上层溶解锰最主要的源, 而对深层水来说水平输送更重要。
Chakraborty等[36]研究了阿拉伯海OMZ带内外沉积物中金属存在的形式的变化。他们利用连续浸取法将沉积物中的金属分为水溶态、离子交换和碳酸盐态、Fe–Mn氧化物态、有机态和残渣态, 研究了底层水中溶解氧含量的变化对Cu、Zn等金属存在状态的影响。底层水溶解氧水平的变化(富氧到缺氧再到低氧)造成了不同结合态金属的再分配。沉积物中铜的活性随底层水溶解氧浓度的降低而逐渐降低, 即稳定性增加。底层水溶解氧含量的降低加强了Cu与沉积物中有机质的联系。但是Pb与Fe–Mn氧化物相的相关性随上覆水中溶解氧浓度的降低而降低。沉积物中Pb配合物的活动性随底层水中氧浓度的降低而增加。底层水中氧的浓度是控制沉积物中Cu、Pb配合物稳定性和活动性的一个关键因素。在阿拉伯低氧带沉积物中与有机质和Fe–Mn氧化物态结合的Cu、Pb是沉积物中最主要的存在形式。Chakraborty等[37]发现阿拉伯海低氧区有机碳的含量随上覆底层水DO水平的降低而增加, 而有机质含量的增加也会导致沉积物中Hg积累的增加。但上覆底层水中DO的消耗不会加强表层沉积物中甲基汞的含量。沉积物中甲基汞含量随有机质和总汞含量的增加而增加, 有机质是控制沉积物中Hg分布和存在形态的关键因子, 沉积物中有机质含量的增加会导致有机汞含量的增加, 但甲基化的速率会降低。总的来说有机质控制了阿拉伯低氧区内沉积物中甲基汞的行为。
2.4 大洋最小含氧带对生态系统的影响
OMZ广泛分布于世界大洋的中层水, 强烈影响了浮游生物和底栖生物的分布和多样性特征, 因此, 其对生态系统的影响也是OMZ研究的重点。Wishner等[8]认为全球气候变化或自然气候波动将使大洋OMZ在全球范围内的空间和垂直分布上进一步扩展, 这种扩展将对生态系统结构和功能产生更加显著的影响。
2.4.1 OMZ影响生物的分布
对东赤道北太平洋的研究发现, 无论OMZ的强度如何, 温跃层是浮游动物生物量最高的区域, 下部氧跃层具有独特的浮游动物组合和第二生物量峰值, 与特定的氧浓度2 μmol/kg密切相关。对阿曼近海OMZ的研究也发现浮游生物和弱游泳生物的生物量在上层100 m水深内的含量最高, 这可能与该层具有较高的氧含量有关。在这一层以胶质生物为主, 还含有一群游泳的蟹类。相当大量的蛇鼻鱼、发光鱼和十足目甲壳动物白天聚集在氧跃层的下方, 但在晚上迁移到表层[38]。Williams等[39]的研究发现热带北太平洋东部(ETNP)浮游植物的同位素组成和营养(级)生态的水平和垂直分布明显受低氧带的控制。最显著的是同位素组成在低氧带的上下界面处有明显垂直梯度。在低氧带的上部δ13C明显偏低, 可能与被浮游动物摄食的化能自养生物有关。在浮游动物的δ15N在下部氧跃层内随深度有一个显著的降低, 说明在紧靠核心低氧带的下方营养物质增加。浅层POM(0~110 m)是混合层、上部氧跃层、核心低氧带浮游动物最重要的食物源, 而深层POM是下部氧跃层浮游动物最主要的食物来源(除了季节性迁移的桡足类)。
浮游动物的分布与溶解氧浓度和梯度关系密切。Saltzman和Wishner[40-41]对东赤道太平洋一海山OMZ浮游动物的分布进行了研究, 发现OMZ是影响温跃层下浮游动物类群垂直分布的重要因素, 在OMZ底层界面区域出现浮游动物第二峰值似乎是OMZ区域的独特现象。不同类群组具有4种垂直分布模式, 说明他们具有不同的低氧耐受能力。最常见的垂直分布模式是在混合层和温度跃层区有最大丰度, 在OMZ下界面区(600~1000 m)出现第二峰值,、、、和是这种分布模式。低氧浓度不限制这些物种的分布, 因为他们在整个OMZ都可出现。第二种垂直分布模式是浮游动物在温跃层和OMZ之间垂直迁移的结果,表现出这种分布模式, 晚上在温跃层区具有最大丰度, 白天在OMZ核心区丰度最大。特别是在在OMZ下界面区(600~1000 m)出现第二峰值。第三种类型的分布是桡足类在OMZ的上、下界面区丰度高。、和就是这种分布模式的主要类群, 他们在混合层中缺失或在混合层和上部OMZ具有类似的丰度。第四种分布模式是桡足类在白天和夜晚主要生活在OMZ上方。在其他OMZ区也发现类似的分布, 表明低氧是主要控制因素。
2.4.2 OMZ影响生物的迁移
Antezana[42]对秘鲁寒流生态系统中OMZ区物种昼夜垂直迁移的特性进行了研究, 发现OMZ决定了迁移强度:只向浅层氧跃层迁移, 而、、和则向OMZ的核心区迁移,向OMZ核心区的底部迁移。在定时迁移和持续迁移中的向上、向下和在浅层或深层的驻留时间也有一些差异。、和在日出时正常向下迁移, 在日落时向上迁移。和在日出时也向下迁移, 但是在下午就开始向上迁移, 缩短了他们在深层水的驻留时间表现出在浅层水中的最长驻留时间和最短的垂直迁移路径。浮游生物定期迁入和迁出OMZ区在白天和夜晚的垂直分层和差异说明群落结构建立在一个通过避免在时间和空间上共存而形成的栖息地分离机制。Escribano等[25]在智利北部也发现类似的分布规律, 如大型桡足类生活在氧跃层的下部, 可通过昼夜迁移进入OMZ, 而丰度很大的在表层水和OMZ核心(200 m)进行昼夜迁移, 甚至穿越OMZ。
2.4.3 不同生物对OMZ的响应不同
Cook等[43]发现低溶解氧对线虫的分布没有影响, 而食物则是影响线虫分布的主要因素。东热带太平洋独有的大型深海鱿鱼——巨型鱿鱼能垂直迁移进入深海OMZ, 但在低氧区鱿鱼的代谢率降低(68%)[44]。Wishner等[45]发现在阿拉伯海OMZ核心区、下部氧跃层和氧跃层底部具有不同的桡足类群落。桡足类在含氧的环境中具有最大的多样性, 物种丰度的排序和下部氧跃层和氧跃层底部类似。在OMZ核心区一些种类缺失或相当少。桡足类在OMZ的垂直分带受低氧、潜在捕食者和潜在食物源的综合控制。总体来说, 生物的粒径越小越有利于在低氧区生存, 因为较小的生物能更好的满足自身新陈代谢的需求[46]。但Teuber等[47]认为无论是深度还是当前的OMZ的低氧条件对浮游动物的呼吸都没有很强的和持久的影响。
2.4.4 低氧区并不意味着是生物贫乏区
虽然从理论上来讲, 低氧不利于生物的生长与生存, 但实际调查结果经常有令人诧异的发现。如Davies等人[48]对东赤道太平洋墨西哥外海从OMZ的上边界(约0.2 mL/L)到海表的仔鱼栖息地的分布进行了分析, 发现氧最低栖息地大部分位于温跃层下面的低氧带(<1 mL/L; 约70 m深)和无氧区(<0.2 mL/L,约80 m深), 而且在低氧栖息地具有最多的种和最高的仔鱼丰度。同样, 一般认为低氧区底栖生物的丰度和多样性都会大大降低, 但在OMZ区底栖生物对低氧的响应和我们想象的完全不同。例如智利中部Concepcion Bay陆架和陆坡的OMZ(122~206 m)大型生物密度较高(16478~21381 individualsm2), 69%~89%的生物为软体生物, 在陆架拐点处(206 m)生物密度最高(21381 individuals/m2), 但生物量最低(16.95 g wet weight/m2), 生物个体最小(0.07 mg C/individuals),多毛类占总生物丰度71%、甲壳类占 16%、软体生物只占2%。在OMZ下方的陆坡中部(约–840 m)甲壳动物49%, 多毛类占43%。虽然有文献表明在OMZ区生物的密度和生物量都低于OMZ下方的区域, 但在该调查区只有生物密度较低。生物量的分布随水深呈凹型, 高值出现在OMZ的上部边缘(122 m)和下方(840 m)。说明大型生物的结构与底层溶解氧浓度、叶绿素a、脱镁叶绿酸和硫化物浓度有关, 与沉积物的粒度、C、N、泥、孔隙度、氧化还原条件、底层水温度等无关[49]。在巴基斯坦半深边缘海OMZ区大型动物的多样性、优势种和洞穴数量在氧浓度(5~9 μmol/kg)时达到极限, 在这个浓度之下表现为较少的生物和较好的沉积层序, 在这个浓度之上表现为较多生物和充分扰动的沉积物。长久洞穴的形成和腐食性决定了OMZ区大型底栖生物的生活方式, 允许沉积层在高生物密度和生物量的条件下存在并保留下来。在永久严重缺氧区, 食物的可利用性决定了动物的丰度和生物结构的深度。在巴基斯坦边缘海氧影响模式的多样性和主导性及其与有机质的相互作用决定了动物区系的分布特征[50]。Levin等[51]对阿拉伯海OMZ内部和下部大型底栖生物的调查也发现了类似的规律, OMZ内的生物密度和生物量要比OMZ下部的海域高, 且个体大小没有显著差异。但Hughes等[52]对巴基斯坦边缘海OMZ大型底栖生物的研究发现, 大型底栖生物的密度在OMZ浅水140m水深站位最高, 但在OMZ下边界(1200 m)密度并未升高, 在OMZ核心(300 m)相当低, 这与阿拉伯其他区域OMZ的调查结果明显不同, 说明在OMZ大型底栖生物的组成受局地因素的影响更重要。另外, Zettler等[53]对纳米比亚北部上升流区域底栖生物的调查发现, 与近岸高生物多样性的区域相比, OMZ底栖生物种类的数量急剧降低, 但是在氧含量0.06~0.88 mL/L的区域内, 生物密度和生物量非常高, 也说明OMZ大型底栖生物的组成与分布受局地因素的影响。
3 大洋最小含氧带(OMZ)研究之思考
大洋最小含氧带(OMZ)一般在水深200~1000 m, 其在大洋的形成主要与厌氧细菌降解有机物导致氧的消耗有关。在深度>1000 m处, 大洋再次复氧, 主要因底层流带来冷的、富氧、极地水有关。在大陆平台, 与OMZ接触的沉积物也是缺氧环境, 有利于埋藏有机质的保存。OMZ在控制全球生态群落结构方面起着重要作用。在这一区带内重要的代谢活动控制着几种与生命有关的元素的循环, 如氮循环。在缺氧带, 生物必须发展出特殊的生存策略, 如反硝化菌、异养厌氧菌能通过反硝化作用消耗硝酸盐中的氧, 后者是将NO3–转化为N2的主要代谢过程, 与全球大气圈-水圈中氮循环有关。另外, 对生活在OMZ的原核生物群落的研究还可以发现能在类似缺氧的环境中生存的特殊生命演化和特殊的分解/代谢活动的重要线索。虽然当前已认识到大洋OMZ在全球物质循环、大洋生态系统和极端环境下生物演化的重要作用, 但对大洋OMZ的认识仍存在许多疑问, 未来几年应当在如下几个方面进行重点研究。
(1) 大洋最小氧带(OMZ)标准的确定。由于当前获得的大洋溶解氧的数据较少, OMZ标准的选取常常依赖于研究者的兴趣(如低氧生物化学过程研究采用的是O2浓度低于20 μmol/kg, 但是物理过程认为这又包括了弱氧化和还原条件), 而且当前在每一个OMZ采取的标准可能是不同的, 有的标准过于苛刻, 使部分溶解氧含量较低的区域不在OMZ的范围之内。所以建立统一的OMZ的标准是对大洋OMZ进行深入研究的基础, 也是必须完成的工作。
(2) 大洋最小氧带(OMZ)的成因。虽然从理论上来说OMZ的形成是中层溶解氧被消耗又得不到及时补充造成的, 但具体到每一个海域又有其特殊成因, 如消耗溶解氧的有机质从哪里来?表层生产力的年变化强度较大, 为什么OMZ的年变化较弱?是什么物理和生物过程决定了低氧区的强度和结构?环流和涡流在高氧水的扩散和混合过程中起什么作用?大洋OMZ强度和全球变化如何相互作用?导致不同区域OMZ强度不同的主要因素是什么?
(3) 大洋OMZ中的氮循环。虽然OMZ被认为是全球氮的汇区, 但异养反硝化作用相对于厌氧氨氧化的相对贡献仍然没有解决。厌氧氨氧化有助于解释OMZ水体中氨的缺乏, 对N2的流失有明显的贡献, 但不能解释N2O的分布和循环, 也不能解释传统的反硝化作用涉及的高基因多样性。另外, 最新的模型结果显示大洋OMZ在全球大气N2固定中起着重要作用, 但对其与氮气流失的耦合关系知之甚少。可见, 对大洋OMZ氮循环过程的研究还需进一步加强。
(4) 大洋OMZ中其他元素的生物地球化学循环过程。大洋低氧环境下氮循环过程也会影响其他元素的生物地球化学循环过程, 比如碳、硫、铁、锰等元素, 加强不同元素迁移转化之间的关系研究, 有助于更好的理解海洋生物地球化学循环。
(5) 大洋OMZ对生态系统的影响。当前对大洋OMZ生态系统中研究最多的是浮游生物和底栖生物, 但对OMZ水体和沉积物中微生物的生态学和生物多样性知之甚少。即使是研究最多的浮游生物, 对他们的新陈代谢/遗传对低氧环境的适应性和他们的生物地球化学作用也知之不多, 更不用说是微生物了。加强大洋OMZ生态系统研究是了解低氧环境下生物地球化学关键过程的前提。
[1] Thamdrup B, Dalsgaard T, Revsbech N P. Wide spread functional anoxia in the oxygen minimum zone of the Eastern South Pacific[J]. Deep-Sea Research I, 2012, 65: 36-45.
[2] Paulmier A, Ruiz-Pino D. Oxygen minimum zones (OMZs) in the modern ocean[J]. Progress in Oceanography, 2009, 80: 113-128.
[3] Helly J, Levin L. Global distribution of naturally occurring marine hypoxia on continental margins[J]. Deep- Sea Research I, 2004, 51: 1159-1168.
[4] Karstensen J, Stramma L, Visbeck M. Oxygen minimum zones in the eastern tropical Atlantic and Pacific oceans[J]. Progress in Oceanography, 2008, 77: 331-350.
[5] Fuenzalida R, Schneider W, Garcés-Vargas J, et al. Vertical and horizontal extension of the oxygen minimum zone in the eastern South Pacific Ocean[J]. Deep- SeaResearchII, 2009, 56: 992-1003.
[6] Ulloa O, Pantoja S. The oxygen minimum zone of the eastern South Pacific[J]. Deep-Sea Research II, 2009, 56: 987-991.
[7] Köllner M, Visbeck M, Tanhua T, et al. Diapycnal diffusivity in the core and oxycline of the tropical North Atlantic oxygen minimum zone[J]. Journal of Marine Systems, 2016, 160: 54-63.
[8] Wishner K F, Outram D M, Seibel B A, et al. Zooplankton in the eastern tropical north Pacific: Boundary effects of oxygen minimum zone expansion[J]. Deep-Sea Research I, 2013, 79: 122-140.
[9] McCreary J P, Yu Z J, Hood R R, et al. Dynamics of the Indian-Ocean oxygen minimum zones[J]. Progress in Oceanography, 2013, 112-113: 15-37.
[10] Raateoja M, Kuosa H, Flinkman J, et al. Late summer metalimnetic oxygen minimum zone in the northern Baltic Sea[J]. Journal of Marine Systems, 2010, 80: 1-7.
[11] Farias L, Paulmier A, Gallegos M. Nitrous oxide and N-nutrient cycling in the oxygen minimum zone off northern Chile[J]. Deep-Sea Research I, 2007, 54: 164- 180.
[12] Galán A, Molina V, Thamdrup B, et al. Anammox bacteria and the anaerobic oxidation of ammonium in the oxygen minimum zone off northern[J]. Chile Deep-Sea Research II, 2009, 56: 1021-1031.
[13] Molina V, Farĺas L. Aerobic ammonium oxidation in the oxycline and oxygen minimum zone of the eastern tropical South Pacific off northern Chile (~20°S)[J]. Deep-Sea Research II, 2009, 56: 1032-1041.
[14] Dalsgaard T, Thamdrup B, Farías L, et al. Anammox and denitrification in the oxygen minimum zone of the eastern South Pacific[J]. Limnology and Oceanography, 2012, 57(5): 1331-1346.
[15] Mashifane T B, Vichi M, Waldron H N, et al. Modelling nitrite dynamics and associated feedback processes in the Benguela oxygen minimum zone[J]. Continental Shelf Research, 2016, 124: 200-212.
[16] Bohlen L, Dale A W, Sommer S, et al. Benthic nitrogen cycling traversing the Peruvian oxygen minimum zone[J]. Geochimicaet Cosmochimica Acta, 2011, 75: 6094- 6111.
[17] Bulow S E, Rich J J, Naik H S, et al. Denitrification exceeds anammox as a nitrogen loss pathway in the Arabian Sea oxygen minimum zone[J]. Deep-Sea Research I, 2010, 57: 384-393.
[18] De Pol-Holz R, Robinson R S, Hebbeln D, et al. Controls on sedimentary nitrogen nisotopes along the Chilemargin[J]. Deep-SeaResearch II, 2009, 56: 1042- 1054.
[19] Schwartz M C, Woulds C, Cowie G L. Sedimentary denitrification rates across the Arabian Sea oxygen minimum zone[J]. Deep-Sea Research II, 2009, 56: 324- 332.
[20] Sommer S, Gier J, Treude T, et al. Depletion of oxygen, nitrate and nitrite in the Peruvian oxygen minimum zone cause an imbalance of benthic nitrogen fluxes[J]. Deep-Sea Research I, 2016, 112: 113-122.
[21] Paulmier A, Ruiz-Pino D, Garcon V. The oxygen minimum zone (OMZ) off Chile as intense source of CO2and N2O[J]. Continental Shelf Research, 2008, 28: 2746-2756.
[22] Franz J, Krahmann G, Lavik G, et al. Dynamics and stoichiometry of nutrients and phytoplankton in waters influenced by the oxygen minimum zone in the eastern tropical Pacific[J]. Deep-Sea Research I, 2012, 62: 20-31.
[23] Pantoja S, Sepúlveda J, González H E. Decomposition of sinking proteinaceous material during fall in the oxygen minimum zone off northern Chile[J]. Deep-Sea Research I, 2004, 51: 55-70.
[24] Pantoja S, Rossel P, Castro R, et al. Microbial degradation rates of small peptides and amino acids in the oxygen minimum zone of Chilean coastal waters[J]. Deep-Sea Research II, 2009, 56: 1055-1062.
[25] Escribano R, Hidalgo P, Krautz C. Zooplankton associated with the oxygen minimum zone system in the northern upwelling region of Chile during March 2000[J]. Deep-Sea Research II, 2009, 56: 1083-1094.
[26] Espinosa L F, Pantoja S, Pinto L A, et al. Water column distribution of phospholipid-derived fatty acids of marinemicroorganisms in the Humboldt Current System off northern Chile[J]. Deep-Sea Research II, 2009, 56: 1063-1072.
[27] Gonsalves M J, Paropkari A L, Fernandes C E G, et al. Predominance of anaerobic bacterial community over aerobic community contribute to intensify ‘oxygen minimum zone’ in the eastern Arabian Sea[J]. Continental Shelf Research, 2011, 31: 1224-1235.
[28] Rad U, Schulz H, Khan A A, et al. Sampling the oxygen minimum zone off Pakistan: glacial-interglacial variations of anoxia and productivity (preliminary results, Sonne 90 cruise)[J]. Marine Geology, 1995, 125: 7-19.
[29] Meadows A, Meadows P S, West F J C, et al. Bioturbation, geochemistry and geotechnics of sediments affected by the oxygen minimum zone on the Oman continental slope and abyssal plain, Arabian Sea[J]. Deep- Sea Research II, 2000, 47: 259-280.
[30] Weijden C H, Reichart G J, Visser H J. Enhanced preservation of organic matter in sediments deposited within the oxygen minimum zone in the northeastern Arabian Sea[J]. Deep-Sea Research I, 1999, 46: 807-830.
[31] Vandewiele S, Cowie G, Soetaert K, et al. Amino acid biogeochemistry and organic matter degradation state across the Pakistan margin oxygen minimum zone[J]. Deep-Sea Research II, 2009, 56, 318-334.
[32] Kondo Y, Moffett J W. Iron redox cycling and subsurface offshore transport in the eastern tropical South Pacific oxygen minimum zone[J]. Marine Chemistry, 2015, 168: 95-103.
[33] Theberge S M, Luther G W, Farrenkopf A M. On the Existence of free and metal complexed sulfide in the Arabian Sea and its oxygen minimum zone[J]. Deep-SeaResearch Ⅱ, 1997, 44(6-7): 1381-1390.
[34] Vedamati J, Chan C, Moffett J W. Distribution of dissolved manganese in the Peruvian Upwelling and oxygen minimum zone[J]. Geochimicaet Cosmochimica Acta, 2015, 156: 222-240.
[35] Lewis B L, Luther III G W. Processes controlling the distribution and cycling of manganese in the oxygen minimum zone of the Arabian Sea[J]. Deep-Sea Research I, 2000, 47: 1541-1561.
[36] Chakraborty P, Chakraborty S, Jayachandran S, et al. Effects of bottom water dissolved oxygen variability on copper and lead fractionation in the sediments across the oxygen minimum zone, western continental margin of India[J]. Science of the Total Environment, 2016, 556: 1052-1061.
[37] Chakraborty P, Mason R P, Jayachandran S, et al. Effects of bottom water oxygen concentrations on mercury distribution and speciation in sediments below the oxygen minimum zone of the Arabian Sea[J]. Marine Chemistry, 2016, 186: 24-32.
[38] Herring P J, Fasham M J R, Weeks A R, et al. Across- slope relations between the biological populations, the euphotic zone and the oxygen minimum layer off the coast of Oman during the southwest monsoon (August, 1994)[J]. Progress in Oceanography, 1998, 41: 69-109.
[39] Williams R L, Wakeham S, McKinney R, et al. Trophic ecology and vertical patterns of carbon and nitrogen stable isotopes in zooplankton from oxygen minimum zone regions[J]. Deep-Sea Research I, 2014, 90: 36-47.
[40] Saltzman J, Wishner K F. Zooplankton ecology in the eastern tropical Pacific oxygen minimum zone above a seamount: 1. General trends[J]. Deep-Sea Research I, 1997, 44(6): 907-930.
[41] Saltzman J, Wishner K F. Zooplankton ecology in the eastern tropical Pacific oxygen minimum zone above a seamount: 2. Vertical distribution of copepods[J]. Deep- Sea Research I, 1997, 44(6): 931-954.
[42] Antezana T. Species-specific patterns of diel migration into the oxygen minimum zone by euphausiids in the Humboldt Current Ecosystem[J]. Progress in Oceanography, 2009, 83: 228-236.
[43] Cook A A, Lambshead P J D, Hawkins L E, et al. Nematode abundance at the oxygen minimum zone in the Arabian Sea[J]. Deep-Sea Research II, 2000, 47: 75-85.
[44] Trübenbach K, Teixeira T, Diniz M, et al. Hypoxia tolerance and antioxidant defense system of juvenile jumbo squids in oxygen minimum zones[J]. Deep-Sea Research II, 2013, 95: 209-217.
[45] Wishner K F, Gelfman C, Gowing M M, et al. Vertical zonation and distributions of calanoid copepods through the lower oxycline of the Arabian Sea oxygen minimum zone[J]. Progress in Oceanography, 2008, 78: 163-191.
[46] Quiroga E, Quin˜ ones R, Palmab M, et al. Biomass size-spectra of macrobenthic communities in the oxygen minimum zone off Chile[J]. Estuarine, Coastal and Shelf Science, 2005, 62: 217-231.
[47] Teuber L, Kiko R, Séguin F, et al. Respiration rates of tropical Atlantic copepods in relation to the oxygen minimum zone[J]. Journal of Experimental Marine Biology and Ecology, 2013, 448: 28-36.
[48] Davies S M, Sánchez-Velasco L, Beier E, et al. Three- dimensional distribution of larval fish habitats in the shallow oxygen minimum zone in the eastern tropical Pacific Ocean off Mexico[J]. Deep-Sea Research I, 2015, 101: 118-129.
[49] Gallardo V A, Palma M, Carrasco F D, et al. Macrobenthic zonation caused by the oxygen minimum zone on the shelf and slope off central Chile[J]. Deep-Sea Research II, 2004, 51, 2475-2490.
[50] Levin L A, Whitcraft C R, Mendoza G F, et al. Oxygen and organic matter thresholds for benthic faunal activity on the Pakistan margin oxygen minimum zone (700—1100 m)[J]. Deep-Sea Research II, 2009, 56: 449-471.
[51] Levin L A, Gage J D, Martin C, et al. Macrobenthic community structure within and beneath the oxygen minimum zone, NW Arabian Sea[J]. Deep-Sea Research II, 2000, 47: 189-226.
[52] Hughes D J, Lamont P A, Levin L A, et al. Macrofaunal communities and sediment structure across the Pakistan margin oxygen minimum zone, North-East Arabian Sea[J]. Deep-Sea Research II, 2009, 56: 434-448.
[53] Zettler M L, Bochert R, Pollehne F. Macrozoobenthos diversity in an oxygen minimum zone off northern Namibia[J]. Mar Biol, 2009, 156: 1949-1961.
The oxygen minimum zones (OMZs) and its eco-environmental effects in ocean
LI Xue-gang1, 2, 3, SONG Jin-ming1, 2, 3, YUAN Hua-mao1, 2, 3, LI Ning1, 2, 3, DUAN Li-qin1, 2, 3, WANG Qi-dong1
(1. Chinese Academy of Sciences Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Laboratory for Marine Ecology and Environmental Sciences, Qingdao National Laboratory for Marine science and Technology, Qingdao 266237, China)
Oxygen minimum zones (OMZs) are the O2-deficient layers in the ocean water column that develops between 200 and 1000 m below the sea level. The development of the OMZ in oceans is mainly related to the consumption of oxygen by aerobic bacteria degrading dead organisms descending through the water column from the above oxygenated ocean layer. There is no currently agreed upon threshold in oxygen that defines an OMZ. If the threshold were chosen as 20 μmol/L, all OMZs surface account for 8% of the present global ocean surface and a mean volume of OMZs account for 7% of the ocean volume. The main regions of OMZ are in the Eastern North Pacific, Eastern South Pacific, Arabian Sea, and Bay of Bengal. In addition, OMZs have been mainly known to play an essential role in the global nitrogen cycle in which various chemical species, according to their degree of oxidation (e.g., ammonium, NH4+; nitrite, NO2; nitrate, NO3; nitrous oxide, N2O; dinitrogen, N2), and different bacterial processes intervene. Under oxic conditions and at the upper boundary (oxycline) of an OMZ, nitrification transforms NH4+into NO3–. However, the core of OMZs is especially associated with denitrification and anaerobic ammonium oxidation (anammox) that can convert NO3–, NH4+, NO2–into gaseous nitrogen (Ngas, for example, N2O, N2) that is lost to the atmosphere and contributes to the oceanic nitrate deficit. The OMZ affects the abundance, distribution, and diversity of plankton and affects the survival and migration of organisms. However, the low oxygen zone does not mean the lack of living organisms. At the same time, the oxygen deficiency in OMZ enhances the preservation of organic matter in sediments and affects the migration and transformation of elements, such as Fe, Mn, and S, that are most sensitive to the redox environment. Although the OMZ has been recognized the important role in the global material cycle and the biological evolution in the ocean ecosystem and extreme environmental conditions, there are still many questions to understand the ocean OMZ. Consequently, further research is needed.
oxygen minimum zones (OMZs); distribution and formation; eco-environmental effects; ocean
(本文编辑: 康亦兼)
[Qingdao National Laboratory for Marine Science and Technology (the Preliminary Deep Sea Research)(2016ASKJ14); Strategic Priority Research Program of the Chinese Academy of Sciences, No. XDA11030202]; Science & Technology Basic Resources Investigation Program of China (2017TY100802)]
Aug.21, 2017
李学刚(1969-), 男, 河南汝南人, 研究员, 博士, 主要从事海洋化学研究, 电话: 0532-82898891, E-mail: lixuegang@qdio.ac.cn; 宋金明,通信作者, 研究员, 博士, 主要从事海洋化学研究, 电话: 0532-82898809, E-mail: jmsong@qdio.ac.cn
P734.2
A
1000-3096(2017)12-0127-12
10.11759/hykx20170821003
2017-08-21;
2017-10-19
青岛海洋科学与技术国家实验室鳌山科技创新计划项目(2016ASKJ14); 中国科学院战略性先导科技专项(XDA11030202); 科技基础资源调查专项(2017FY100802)
