蓝身大斑石斑鱼染色体核型分析
2017-05-04杨宇晴张海发吴锦辉黄锦雄石和荣
刘 苏, 杨宇晴, 2, 张海发, 张 勇, 2, 吴锦辉, 黄锦雄, 石和荣
蓝身大斑石斑鱼染色体核型分析
刘 苏1, 杨宇晴1, 2, 张海发1, 张 勇1, 2, 吴锦辉1, 黄锦雄1, 石和荣1
(1. 广东省海洋渔业试验中心, 广东 惠州 516081; 2. 中山大学 水生经济动物研究所 暨广东省水生经济动物良种繁育重点实验室, 广东 广州 510275)
为了解蓝身大斑石斑鱼()的细胞生物学特征, 作者采用体内注射PHA培养肾细胞法对蓝身大斑石斑鱼染色体核型进行了研究。结果表明, 其染色体核型为2=48t, 属于石斑鱼属鱼类典型的高位类中原始类群, 是石斑鱼属进化过程中出现较早的物种。
蓝身大斑石斑鱼(); 核型; 染色体
鱼类是脊椎动物在自然界中分布最广、种类最多的类群, 中国鱼类资源丰富, 多样性水平高, 经济价值大。染色体作为遗传信息的载体, 其核型具有一定的物种特异性, 对鱼类染色体的研究可以探讨鱼类种群的进化和亲缘关系, 为现代遗传学提供有价值的理论数据, 为鱼类资源的开发利用提供科学的指导。
蓝身大斑石斑鱼()是中国沿海养殖经济鱼类之一, 其生长速度快、营养丰富, 具有很高的市场价值, 养殖前景十分广阔。蓝身大斑石斑鱼属中大型鱼类, 性成熟时间较晚, 一般需要3~4龄鱼才能达到初次性成熟, 亲鱼的培育难度大, 其苗种来源主要靠天然捕捞, 严重制约了养殖产业的发展。目前对蓝身大斑石斑鱼的研究较少, 仅见于线粒体基因克隆[1]、神经坏死病毒传播研究[2]及产卵和幼苗培育的研究[3-4]。本研究利用体内注射PHA肾细胞培养法对蓝身大斑石斑鱼染色体核型进行了分析, 旨在开展蓝身大斑石斑鱼繁殖生物学的基础研究, 可为其繁育提供数据支持, 为其繁育研究奠定基础。
1 材料和方法
1.1 材料
试验用蓝身大斑石斑鱼取自广东省海洋渔业试验中心, 共取样2尾, 鱼体质量分别为922.4 g和1 341.3 g。
1.2 实验方法
1.2.1 前处理
试验鱼按5 mL/kg鱼体质量进行抽血刺激, 按10 μg/g鱼体质量进行胸腔PHA一次性注射, 5 h后注射1 μg/g的秋水仙素, 1.5 h后剪开鳃部动脉放血20 min,取头肾于0.9%生理盐水中洗涤, 剪碎, 用纱布过滤细胞悬液至离心管中, 1 000 r/min离心10 min收集沉淀细胞, 用生理盐水洗涤细胞2次。
1.2.2 制片
低渗: 向离心管中加入5 mL的0.075 mol/L的KCl低渗液, 30℃处理23~33 min, 1 000 r/min离心10 min,弃上清, 留底部沉淀。
固定: 加入5 mL预冷新配的卡诺氏固定液(甲醇︰冰醋酸=3︰1), 用吸管轻轻吹打沉淀至充分混匀, 固定30 min后1 000 r/min离心10 min, 弃上清, 反复2次, 第3次固定于冰上进行, 弃上清, 留底部沉淀, 加入2~4 mL固定液重新悬浮细胞, 冰浴10 min。
滴片: 冷滴片法制片。取出预冻于无水乙醇的干净玻片, 于滤纸上稍微擦干, 立刻进行滴片。滴片前先混匀悬液, 然后用滴管吸取少量悬液, 于距离玻片约50 cm高处滴下, 自然风干。
染色: 待玻片完全干透后用10% Giemsa染液倒扣染色30 min, 用蒸馏水冲洗干净并晾干。
1.2.3 核型分析
用光学显微镜(Olympus)观察并拍照, 选取清晰的染色体中期分裂相计数确定染色体众数; 选取10个分散效果良好、着丝粒清晰、长度适中、两条单体适度分开、形态清楚的分裂相放大, 确认着丝点位置, 测量并统计每条染色体的相对长度、臂比, 按照Levan等(1964)的标准进行配对, 分类排列核型。
2 结果
对100个图像清晰, 染色体分散良好的中期分裂相细胞进行统计得出, 蓝身大斑石斑鱼染色体众数为48, 众数出现频率为68%(表1)。对10个中期分裂相进行镜检和测量, 得出蓝身大斑石斑鱼染色体核型公式为2=48t, 由48条端部着丝粒染色体组成(图1)。

表1 蓝身大斑石斑鱼染色体众数统计
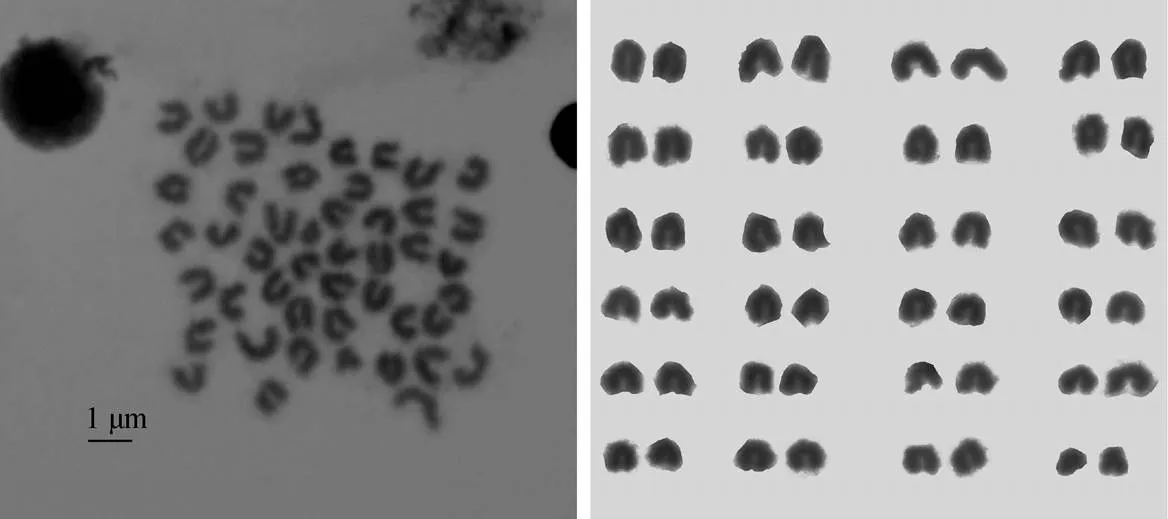
图1 蓝身大斑石斑鱼染色体核型
由表2可见, 在蓝身大斑石斑鱼48条染色体中, 相对长度最大的为2.69%±0.11%; 相对长度最小的为1.38%±0.09%, 明显短于其他染色体。相对长度最大的染色体与相对长度最小的染色体比值为1.95。
3 讨论
3.1 石斑鱼染色体制备
无论是何种生物, 获取新陈代谢活动旺盛、分裂增生较快的组织都是染色体制片取得成功的基本前提[5], 生命活动越旺盛, 则染色体标本的制备就越简单, 获得的细胞分裂指数也越高。大量的研究证实PHA能够促进细胞的增殖[6-8], 但每种鱼类对PHA的处理浓度和时间差异较大[9-12], 在本研究中, 为获得更多的分裂相, 除利用10 μg/g鱼体质量的PHA进行腹腔注射外, 还对试验鱼进行了抽血刺激。结果表明, 抽血和PHA双重刺激的处理方法能有效促进细胞增殖, 获得数量较多、效果较好的染色体中期分裂相, 与王德祥等[13]研究结果相一致。
3.2 石斑鱼染色体核型与系统分类和种质鉴定的关系
中国已发现的海水鱼类染色体数目为26~56, 大部分海水鱼类染色体数目为2=48, 根据原始性状普遍性原则, 2=48被认为是海水鱼类原始核型数目[14-15]。
现有研究表明, 石斑鱼二倍体种类的染色体数目均为2=48, 染色体数目具有高度一致性, 但不同鱼种类的核型结果不同, 可用作鉴别不同种属的重要依据[16-17]。目前, 石斑鱼的分类主要依据其外部形态特征, 但其体色和斑纹易受到环境、年龄和生理变化的影响而发生改变, 给分类鉴定造成极大的困难。许多石斑鱼的外形特征极为相似, 特别是在应激状态下, 难以根据外部形态将它们区分开来, 如七带石斑鱼()和褐石斑鱼()、斜带石斑鱼()和点带石斑鱼()等,然而它们染色体核型差异明显, 七带石斑鱼的核型为2=48t, 褐石斑鱼的染色体核型为2=2m+4sm+42t, 斜带石斑鱼的核型为2=2sm+46t, 点带石斑鱼的染色体核型为2=48t[18-19]。可见形态差异大的石斑鱼染色体核型不同, 染色体核型相同的形态差异较大, 因此染色体核型的多样化对石斑鱼的系统分类和种类鉴定有着重要的辅助作用。
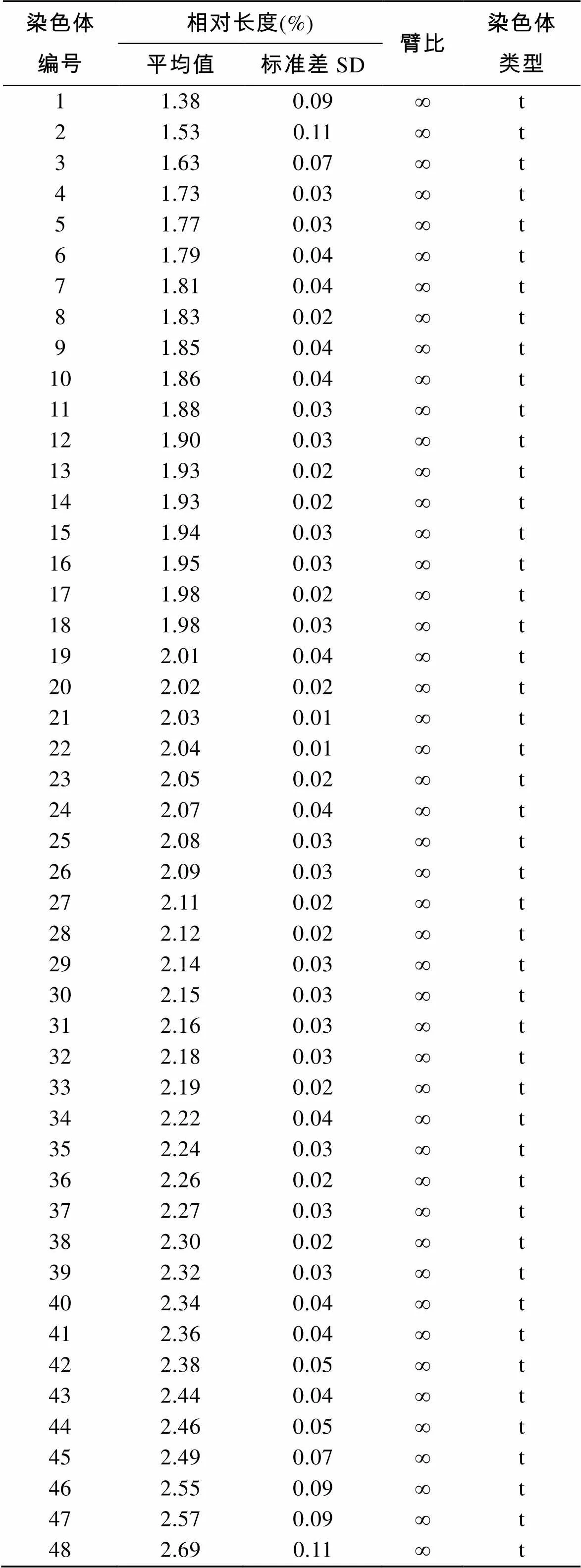
表2 蓝身大斑石斑鱼染色体相对长度和臂比
在本研究的试验过程中, 作者发现蓝身大斑石斑鱼在强光照射或应激情况下, 鱼体色变深, 体表的斑纹会发生变化, 鱼体侧会出现隐约的条带性斑纹, 与斜带石斑鱼、褐石斑鱼、青石斑鱼()等的外表特征相似, 不易根据外部形态区分它们, 但蓝身大斑石斑鱼和斜带石斑鱼、褐石斑鱼的染色体核型组成有较大的差异, 蓝身大斑石斑鱼的染色体核型为2=48t, 斜带石斑鱼的染色体核型为2=2sm+46t, 褐石斑鱼的染色体核型为2=2sm+46t, 因此根据染色体核型能很好地将其区分开来。但是, 石斑鱼种类繁多, 从现有的研究结果来看, 仅依据染色体的核型分析进行石斑鱼类种质鉴定是远远不够的, 若能进一步结合染色体的C带、银染等多种方法, 将提高石斑鱼类的系统分类的准确性, 为石斑鱼类种质鉴定提供更有力的佐证。
3.3 蓝身大斑石斑鱼染色体核型与种类进化的关系
鱼类的演化程度与其细胞染色体类型是一致的, 进化中处于上位的, 染色体数目越收敛, 端部着丝粒染色体多, 臂数少[20]。李树深[21]认为在特定分类阶元中, 具有较多中部或亚中部着丝粒染色体的是特化类群, 而具有较多端部着丝粒染色体的为原始种群, 也就是说染色体臂数多的比染色体臂数少的种类更特化。王世锋[19]在对6种石斑鱼染色体核型进行比较分析后指出, 原始类群的石斑鱼在进化过程中, 其染色体结构未发生大的变化, 基本保持了原始核型特征, 特化类群的石斑鱼在进化过程中染色体结构发生了易位、倒位等重组, 形成染色体特化核型特征。在本研究中, 蓝身大斑石斑鱼的染色体核型为2=48t, 染色体相对长度从1.38±0.09到2.69± 0.11, 相差不大, 收敛性强, 染色体臂数少, 推测其应属于典型的高位类中原始类群, 是石斑鱼属演化过程中出现较早的物种。由于石斑鱼种类繁多, 而其染色体数目具有高度一致性, 因此有必要利用染色体显带和荧光原位杂交等技术对蓝身大斑石斑鱼的同源染色体进行更为精确的分析和鉴定, 为其种类进化提供更为翔实的数据资料。
[1] Yang Y Q, Xie Z Z, Peng C, et alThe complete mitochondrial genome of the(Perciformes, Serranidae)[J]. Mitochondrial DNA Part A, 2016, 27(1): 520-522.
[2] Yue G H, Li Y , Chen F, et al. Comparison of three DNA marker systems for assessing genetic diversity in Asian arowana ()[J]. Electrophoresis, 2002, 23: 1025-1032.
[3] Yeh S L, Dai Q C, Chu Y T, et al. Induced sex change, spawning and larviculture of potato grouper,[J]. Aquaculture, 2003, 228: 371-381.
[4] 郑飞, 韩丽娜, 王永波, 等. 金钱斑亲鱼培育技术[J]. 现代农业科技, 2016, 14: 261-270.
Zheng Fei, Han Lina, Wang Yongbo, et al. Culture technology of[J]. Modern Agricultural Science and Technology, 2016, 14: 261-270.
[5] 吴彪, 杨爱国, 周丽青, 等. 几种鱼类染色体制备方法的比较[J]. 安徽农业科学, 2012, 40(12): 7168-7170.
Wu Biao, Yang Aiguo, Zhou Liqing, et al. Comparison of several preparation methods for fish chromosome[J]. Journal of Anhui Agricultural Sciences, 2012, 40(12): 7168-7170.
[6] 林义浩. 快速获得大量鱼类肾细胞中期分裂相的PHA体内注射法[J]. 水产学报, 1982, 6(3): 201-208.
Lin Yihao. A PHA injection methodfor the rapid obtainment of large numbers of metaphase figures from kidney cells of teleosts[J]. Journal of Fisheries of China, 1982, 6(3): 201-208.
[7] 牛文涛, 蔡泽平. 中国海水鱼类核型研究概述[J]. 厦门大学学报(自然科学版), 2006, 45(2): 125-131.
Niu Wentao, Cai Zeping. Review on karyotype study of marine fishes in China[J]. Journal of Xiamen University (Natural Science), 2006, 45(2): 125-131.
[8] 张伟明, 吴萍, 吴康, 等. 两种鱼类染色体制片方法的比较研究[J]. 水利渔业, 2003, 23(5): 9-10.
Zhang Weiming, Wu Ping, Wu Kang, et al. Comparison of two preparation methods for fish chromosome[J]. Reservoir Fisheries, 2003, 23(5): 9-10.
[9] 舒琥, 蔡晓阅, 刘峰, 等. 鲀形目3种鱼的染色体组性分析[J]. 动物学杂志, 2010, 45(2): 101-106.
Shu Hu, Cai Xiaoyue, Liu Feng, et al. Karyotypes analysis for three species of tetraodontiformes fishes[J]. Chinese Journal of Zoology, 2010, 45(2): 101-106.
[10] 蔡岩, 周永灿, 张本, 等. 海南野生三斑石斑鱼染色体核型分析[J]. 热带生物学报, 2011, 2(2): 97-100.
Cai Yan, Zhou Yongcan, Zhang Ben, et al. Chromosome karyotype analysis of(Valenciennes, 1828)[J]. Journal of Tropical Organisms, 2011, 2(2): 97-100.
[11] 李锡强, 彭跃东. 斑带石斑鱼与黑边石斑鱼核型的研究[J]. 湛江海洋大学学报, 1994, 14(2): 22-26.
Li Xiqiang, Peng Yuedong. Studies on karyotype ofand[J]. Journal of Zhanjiang Fisheries College, 1994, 14(2): 22-26.
[12] 霍蕊, 张本, 陈国华, 等. 波纹唇鱼染色体核型分析[J]. 海洋科学, 2009, 33(4): 94-97.
Huo Rui, Zhang Ben, Chen Guohua, et al. The karyotype of[J]. Marine Sciences, 2009, 33(4): 94-97.
[13] 王德祥, 苏永全, 王世锋, 等. 宽额鲈染色体核型研究及制作方法的比较[J]. 台湾海峡, 2003, 22(4): 465- 468.
Wang Dexiang, Su Yongquan, Wang Shifeng, et al. Study on the karyotype of[J]. Journal of Oceanography in Taiwan Strait, 2003, 22(4): 465-468.
[14] 王世锋, 王德祥, 苏永全, 等. 双棘黄姑鱼染色体组型分析[J]. 厦门大学学报(自然科学版), 2003, 42(5): 682-684.
Wang Shifeng, Wang Dexiang, Su Yongquan, et al. The karyotypes of[J]. Journal of Xiamen University(Natural Science), 2003, 42(5): 682-684.
[15] 陈淑芬, 汤泽生. 脊椎动物染色体数目及其变化趋势[J].四川师范学院学报(自然科学版), 1997, 18(1): 18-23.
Chen Shufen, Tang Zesheng. The number and changing tendency of the vertebrate chromosome[J]. Journal of Sichuan Teachers College(Natural Science), 1997, 18(1): 18-23.
[16] 郑莲, 刘楚吾, 李长玲. 4种石斑鱼染色体核型研究[J]. 海洋科学, 2005, 29(4): 51-55.
Zheng Lian, Liu Chuwu, Li Changling. Studies on the karyotype of 4 groupers[J]. Marine Sciences, 2005, 29(4): 51-55.
[17] 舒琥, 魏秋兰, 罗丽娟, 等. 广东沿海4种石斑鱼的染色体组型分析[J]. 广东农业科学, 2012, 8: 124-127.
Shu Hu, Wei Qiulan, Luo Lijuan, et al. Karyotypes analysis of four grouper fishes from coastal waters of Guangdong[J]. Guangdong Agricultural Sciences, 2012, 8: 124-127.
[18] 钟声平, 陈超,王军. 七带石斑鱼染色体核型研究[J]. 中国水产科学, 2010, 17(1): 150-155.
Zhong Shengping, Chen Chao, WANG Jun, et al. Chromosome karyotype of sevenband grouper(Thunberg, 1793)[J]. Journal of Fishery Sciences of China, 2010, 17(1): 150-155.
[19] 王世锋. 六种石斑鱼核型特征比较和染色体进化研究[D]. 厦门: 厦门大学, 2007.
Wang Shifeng. Comparative karyotype characterization and chromosomal evolution of six groupers[D]. Xiamen: Xiamen University, 2007.
[20] 小岛吉雄.水产生物及遗传育种[M]. 水交出版社, 1979: 46-62.
Ojima. Genetics and breeding of aquatic life[M]. Tokyo: Back Water Press, 1979: 46-62.
[21] 李树深. 鱼类细胞分类学[J]. 生物科学动态, 1981, 2: 8-15.
Li Shusen. Taxonomy of fish cell[J]. Bio-science, 1981, 2: 8-15.
The karyotype of
LIU Su1, YANG Yu-qing1, 2, ZHANG Hai-fa1, ZHANG Yong1, 2, WU Jin-hui1, HUANG Jin-xiong1, SHI He-rong1
(1. Marine Fisheries Development Center of Guangdong Province, Huizhou 516081, China; 2. Institute of Aquatic Economic Animals and Guangdong Provincial Key Laboratory for Aquatic Economic Animals, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China)
This study aims to assess the cell biological characteristics of. We analyzed the karyotype offrom their renal tissue using the PHA and colchicine injection methods. The results revealed 48 telocentric chromosomes in the diploid number, and their karyotypic formula was 2= 48t. The study deduces thatcomprises the characteristics representative of the ancestral karyotypic pattern and belongs to the primitive groups.
; karyotype; chromosome
(本文编辑: 谭雪静)
[Special Fund for Agro-scientific Research in the Public Interest, No.201403011; Special Fund for Marine Fisheries Development in Guangdong Province, No.201743; Special Fund for Fishing Port Construction and Fishery Development in Guangdong Province, No.201611; Special Fund for Marine Fishery Science and Industry Development in Guangdong Province, No.A201400A01, A201600A02; Science and Technology Planning Project of Guangdong Province, No.2015A020208002]
Jun. 13, 2017
刘苏(1985-), 男, 湖南省宁远人, 工程师, 硕士, 主要从事海洋生物繁育学研究, 电话: 0752-5574826, E-mail: 306151183@qq.com
Q343.22
A
1000-3096(2017)12-0046-05
10.11759//hykx20170613002
2017-06-13;
2017-09-13
公益性行业(农业)科研专项(201403011); 广东省海洋和渔业发展专项资金(201743); 广东省渔港建设和渔业发展专项资金(201611); 广东省海洋渔业科技与产业发展专项(A201400A01, A201600A02); 广东省科技计划项目(2015A020208002)
