共表达磷脂酶C促进葡萄糖异构酶在大肠杆菌中的胞外表达
2017-05-03宿玲恰
姜 琪, 宿玲恰, 吴 敬, 陈 晟
(食品科学与技术国家重点实验室,江南大学,江苏 无锡 214122)
共表达磷脂酶C促进葡萄糖异构酶在大肠杆菌中的胞外表达
姜 琪, 宿玲恰, 吴 敬, 陈 晟*
(食品科学与技术国家重点实验室,江南大学,江苏 无锡 214122)
磷脂酶C(PLC)能够水解细胞膜主要成分磷脂,使细胞膜通透性增强,从而能够释放出胞内物质。作者构建了PLC与天然胞内定位蛋白葡萄糖异构酶(GI)共表达的重组大肠杆菌,摇瓶发酵胞外上清液中GIC酶活达到3.4 U/mL,占胞外和胞内总酶活的93%,表明GIC成功实现了胞外表达。将胞外上清液中的GIC进行分离纯化和酶学定性,发现其比活为12.1 U/mg,最适反应温度为80℃,最适pH为10,均与对照菌单独表达的GIO性质基本一致。在此基础上,对上述重组菌进行3 L发酵罐培养,发酵周期为24 h,酶活达到17.7 U/mL,表明其良好的工业化放大生产前景。
磷脂酶C,葡萄糖异构酶,共表达,胞外表达
大肠杆菌表达系统作为目前研究最深入和应用最广泛的表达系统,具备遗传背景清晰、细胞结构简单、蛋白质表达效率高、培养周期短、操作简便等天然优势,是规模化制备重组蛋白的首选表达系统之一[1-2]。大肠杆菌具有两层细胞膜结构,这一特点决定了蛋白质的定位方式除了膜定位外,还有胞内定位、周质空间定位和胞外基质定位。重组蛋白质分泌至周质空间或者胞外基质比定位于胞内更有助于蛋白质折叠、减少包涵体的形成、降低胞内杂蛋白质的污染以及简化下游分离提取过程,而胞外基质定位蛋白质的分离过程只需离心过滤除去细胞即可,在大规模生产生物制品中具有较大优势[3-4]。
对于胞内蛋白质来说,通常连接信号肽也无法利用宿主菌蛋白质转运系统实现胞外分泌,因此只能通过破碎细胞来获取。常用的方法有机械法,包括高压匀浆破碎、超声波破碎等;非机械法,包括渗透压冲击破碎、冻融破碎,另外还有化学法破碎等。此外,还有研究者采用共表达一些噬菌体来源的溶解细胞蛋白实现蛋白质的释放[5-7]。这些使细胞完全裂解破碎的方法在过程中无法避免的释放出细胞内蛋白质、核酸、多糖等杂质,这些杂质会给下游的分离提取过程带来不利影响。
本实验室前期研究表明[8],角质酶具有磷脂酶B的水解活性,当其在胞内重组表达时能够有限的水解细胞膜磷脂组分,在一定程度上对细胞膜形成破坏,提升其通透性,从而使内容物能够非正常释放,但不会引起细胞裂解,因此能够通过与角质酶共表达实现胞内目的蛋白质胞外表达的目的。然而,由于角质酶催化磷脂水解的作用位点为磷脂分子中1位和2位酯键[9],其产物之一是溶血磷脂,属于较强的表面活性剂,导致在发酵过程中大量起泡,不利于发酵调控和工业放大。
磷脂酶C (phospholipase C,PLC,EC3.1.4.3)是一种水解甘油磷脂C3位点磷酯酰键生成甘油二酯和磷酸胆碱、磷酸肌醇或磷酸乙醇胺等的水解酶[10],有报道[11]中提到Bacillus cereus来源的磷脂酶C(PLC)同样对细胞存在一定毒性。作者所在实验室前期工作构建了Bacillus cereus来源的磷脂酶C在大肠杆菌中重组表达的工程菌,研究表明,其能够在不引起细胞裂解的情况下通过对细胞膜磷脂的部分水解提升膜透性,并且能够避免角质酶促进蛋白质胞外表达过程中易起泡的缺陷。
葡萄糖异构酶(glucoseisomerase,GI,EC5.3.1.5),又称木糖异构酶,能将葡萄糖、木糖、核糖等醛糖催化异构为相应的酮糖[12]。目前其主要应用领域为将葡萄糖转化为果糖从而制备果葡糖浆。葡萄糖异构酶来源广泛,包括近百种细菌和放线菌[13],其中绝大部分为胞内酶,研究者在进行其重组表达时通常采取胞内定位的形式。作者所在实验室前期构建了Thermobifida fusca来源的葡萄糖异构酶在大肠杆菌中重组表达的基因工程菌,研究表明其酶学特性和应用性能良好,具备良好的工业化应用潜力[14]。
作者研究了通过共表达B.cereus来源的磷脂酶C(PLC)和T.fusca来源的葡萄糖异构酶,考察磷脂酶C是否能够通过限制性地破坏细胞膜促使葡萄糖异构酶的胞外“释放”,并考察在这一过程中,葡萄糖异构酶是否发生折叠不正常、性质改变等问题。
1 材料与方法
1.1 菌株与质粒
E.coli JM109、BL21(DE3)菌株、葡萄糖异构酶胞内表达的重组E.coli BL21(DE3)及分别带有磷脂酶C和葡萄糖异构酶基因的重组质粒pETDuet/plc和pET24a/glu:作者所在实验室保藏。
1.2 试剂与培养基
限制性内切酶NdeⅠ、XhoⅠ,碱性磷酸酶(calf intestine alkaline phosphatase,CIAP),T4DNA连接酶,DNA Marker及琼脂糖:宝生物工程(大连)有限公司;质粒小提试剂盒及琼脂糖凝胶DNA回收试剂盒:天根生化科技有限公司;异丙基硫代-β-D半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG)和氨苄青霉素(ampicillin,Amp):生工生物工程(上海)股份有限公司;分子级酵母粉和胰蛋白胨:英国Oxoid公司;其它试剂:均为国产分析纯试剂,上海国药集团化学试剂有限公司。
LB液体培养基 (g/L):酵母粉 5.0,胰蛋白胨10.0,NaCl 10.0。
LB固体培养基:LB液体培养基中添加1.5~2.0 g/dL的琼脂。
TB培养基(g/L):甘油5.0,胰蛋白胨12.0,酵母粉24.0,K2HPO4·3H2O 16.4,KH2PO42.3。
LB(TB)-Amp培养基:在灭菌后的LB(TB)培养基中添加终质量浓度为100 μg/mL的氨苄青霉素。
3 L罐发酵培养基 (g/L):甘油 8.0,KH2PO413.5,(NH4)2HPO44.0,柠檬酸1.7,MgSO4·7H2O 1.4,微量元素液 12.0 mL,工业级酵母粉2.4,工业级蛋白胨1.2,调pH 7.0。
微量元素液 (g/L):FeSO4·7H2O 10.0,ZnSO4· 7H2O 5.25,CuSO4·5H2O 3.0,MnSO4·4H2O 0.5,Na2B4O7·10H2O 0.23,CaCl22.0,(NH4)6Mo7O240.1。
补料液 (g/L):甘油 600,MgSO49.0,工业级酵母粉2.4,工业级蛋白胨1.2。
1.3 方法
1.3.1 大肠杆菌表达质粒的构建 提取作者所在实验室保藏的重组质粒pET24a/glu及pETDuet/plc,经NdeⅠ、XhoⅠ双酶切,割胶回收获得glu基因及线性化的pETDuet/plc载体,16℃连接过夜,转化E.coli JM109感受态细胞,氨苄青霉素抗性平板筛选。获得的转化子提取质粒经NdeⅠ、XhoⅠ双酶切验证正确,即质粒pETDuet/plc/glu。将验证正确的pETDuet/plc/glu质粒转化E.coli BL21(DE3)感受态细胞,氨苄青霉素抗性平板筛选,-80℃甘油管保存。
1.3.2 重组菌诱导表达 将葡萄糖异构酶和磷脂酶C共表达的重组菌(以下简称共表达菌)与葡萄糖异构酶单独表达的对照重组菌 (以下简称对照菌)置于LB培养基中,37℃、200 r/min振荡培养8~10 h,以5%接种体积分数转接至TB培养基中,25℃、200 r/min培养6 h后加入IPTG,至终浓度为0.1 mmol/L,继续在25℃、200 r/min培养至发酵液中GI酶活不再上升。
1.3.3 葡萄糖异构酶纯化及SDS-PAGE凝胶电泳检测
1)前处理:将共表达菌发酵液离心收集上清液。对照菌发酵液离心收集菌体,菌体用适量30 mmol/L磷酸盐缓冲液(Na2HPO4-KH2PO4,pH 7.5)复溶,超声破碎细胞,离心收集上清液。将上述收集的共表达菌发酵上清液和对照菌破壁上清液于75℃处理15 min,离心,收集上清液。
2)硫酸铵沉淀:在经过前处理的上清液中缓慢加入70 g/dL硫酸铵,盐析过夜,4℃、8 000 r/min离心30 min;用适量缓冲液A(30 mmol/L Na2HPO4-KH2PO4,pH 7.5)将沉淀充分溶解,4℃透析过夜,经膜过滤后即为上样样品。
DEAE-Sepharose阴离子交换柱用缓冲液A平衡后上样,依次用缓冲液A、含0~1 mol/L NaCl的缓冲液A、含1 mol/L NaCl的缓冲液A洗脱结合蛋白质,流速为1 mL/min。收集有酶活力组分进行酶活测定和蛋白质电泳分析。
1.3.4 酶活力检测方法 葡萄糖异构酶活力检测方法:以3 mol/L的葡萄糖溶液为底物,葡萄糖异构酶将底物异构化为果糖,进而采用咔唑-硫酸法测定果糖含量。异构反应为:0.1 mL的3 mol/L葡萄糖溶液,0.1 mL的50 mmol/L的MgSO4,0.1 mL的300 mmol/L pH 7.5 Na2HPO4-KH2PO4缓冲液,0.6 mL H2O混匀,70℃预热5 min,加入0.1 mL适当稀释的酶液(以水作空白),准确反应10 min后,立即用1 mL的0.5 mol/L HClO4终止反应。显色反应:将上述反应液稀释一定倍数后取0.5 mL加入到比色管中,再分别加入0.1 mL半胱氨酸盐酸盐溶液,3 mL 75%的H2SO4和0.1 mL咔唑-酒精溶液,振荡混匀后置于60℃水浴中反应10 min。置于冰浴冷却至室温后,于560 nm处测定吸光度(空白为以水代替反应液)。
酶活力单位定义:在上述反应条件下,每分钟生成1 μmol果糖所需的酶量为一个活力单位(U)。1.3.5 重组菌株生长曲线测定 从甘油管中接种共表达菌及对照菌于LB培养基中,37℃、200 r/min培养8~10 h,5%接体积分数转接于TB培养基中,37℃、200 r/min培养,定时取样测定OD600nm值及酶活力,做2组平行实验。
1.3.6 重组GI最适温度及最适pH测定 其他条件不变,分别在55、60、65、70、75、80、85、90℃测定纯化共表达重组GI酶活,以酶活最高的为100%。
其他条件不变,分别测定pH 5.5~12之间的酶活,以酶活最高的为100%。
1.3.7 3 L发酵罐培养方法 从甘油管中吸取100 μL菌液,接种于含有100 μg/mL氨苄青霉素的工业级LB培养基(装液量50/250 mL)中,37℃、200 r/min培养8 h。将种子以10%的接种体积分数接入Infors 3 L全自动发酵罐中,初始装液量1.2 L。诱导前培养温度37℃、pH 7.0,溶氧20%。当发酵初始碳源甘油基本消耗完时,开始流加补料液;流加方式为指数流加,比生长速率控制在μ=0.2 h-1[15]。当菌体干重约为7.5 g/L左右时,加入甘氨酸,加量为10 g/L;菌体干重约为25 g/L左右时,开始诱导,诱导方法为恒速流加乳糖,乳糖加量0.2 g/(L·h)。开始诱导后,每3小时取样一次,测定菌体干重和培养基中酶活变化,培养基中酶活不再上升时,发酵过程结束。
2 结果与讨论
2.1 重组表达质粒的构建
分别提取 pETDuet/plc质粒及 pET24a/glu质粒,经NdeⅠ、XhoⅠ双酶切,pETDuet/plc酶切之后再用CIAP去磷酸化处理以防止自连,目的片段回收后用T4DNA连接酶于16℃连接过夜,转化E.coli JM109感受态细胞,氨苄青霉素抗性平板筛选。获得的转化子提取质粒经NdeⅠ、XhoⅠ双酶切验证,得到目的条带,见图1。重组质粒pETDuet/plc/glu构建成功。

图1 重组质粒pETDuet/plc/glu NdeⅠ、XhoⅠ双酶切电泳图Fig.1 Restriction enzyme digestion analysis of pETDuet/ plc/glu by NdeⅠand XhoⅠ
bp
2.2 重组葡萄糖异构酶在大肠杆菌中的表达
将验证正确的重组质粒pETDuet/plc/glu转化感受态细胞E.coli BL21(DE3),筛选获得转化子,将该转化子与实验室前期构建的单独表达GI菌株按照上述1.3.2节方法进行诱导培养,结果见图2。共表达菌(以下简称共表达GI为GIC)与对照菌(以下简称单独表达GI为GIO)生长情况相似,在发酵过程的前8个小时,共表达菌培养基中检测不到GIC酶活,推测可能是此时PLC对细胞的破坏作用尚未达到能够使GIC渗漏的程度。发酵8 h后,随着时间的增加,培养基中GIC酶活逐渐增加,直至24 h,随着时间的推移酶活几乎不增长,发酵过程结束,此时培养基中GIC酶活达到3.4 U/mL,胞内酶活约为0.3 U/mL,GIC总酶活为3.7 U/mL,因此释放至胞外的酶活约占总酶活的93%。蛋白质电泳结果见图3。共表达菌培养基上清组分中在43 000处有明显的条带,与GI理论相对分子质量一致,表明与PLC共表达时,GI被高效释放到胞外上清液基质中。此外,对照菌发酵过程结束,酶活达到4.2 U/mL,共表达菌与对照菌葡萄糖异构酶表达量区别不大。
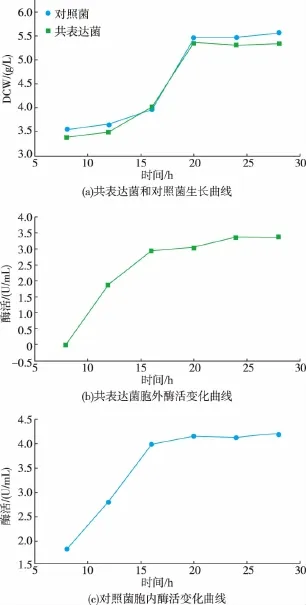
图2 共表达菌和对照菌发酵过程生长和产酶曲线Fig.2 Growth and enzyme production by recombinant E.coli BL21(DE3)
2.3 重组蛋白质的纯化及定性
当GI与PLC共表达时,PLC破坏细胞膜,GI被释放到培养基中的过程是细胞非自然生长状态下渗漏的结果。在这一过程中,可能伴随着GI折叠不完全、结构松散、酶学性能改变等现象。为了考察这些现象是否存在,对GIC进行了分离纯化,并检测了最适温度、最适pH、比活等性质,与实验室保存的GIO纯品性质进行比较。

图3 重组菌摇瓶发酵胞外上清液SDS-PAGE电泳分析Fig.3 SDS-PAGE analysis of culture supernatant of recombinant strains
根据前述1.3.3节方法对粗酶液进行分离纯化。经历了两步纯化后,结果见图4,达到电泳纯。最终获得纯酶GIC比活11.9 U/mg,与作者所在实验室纯化GIO比活12.1 U/mg相近,见表1。

图4 纯酶GICSDS-PAGE电泳图Fig.4 SDS-PAGE analysis of purified GIC

表1 GIC纯化过程参数Table 1 Purification scheme of recombinant GIC
获得GIC纯酶后,分别测定GIC和GIO纯酶最适温度和最适pH,分别选择温度范围55~90℃和pH范围5.5~12进行实验,结果见图5。GIC最适温度为80℃,最适pH为10.0,与GIO一致。且二者随着温度和pH值变化,残留酶活百分比趋势基本一致。

图5 温度和pH对GIC和GIO酶活的影响Fig.5 Effects of temperature and pH on activity of GICand GIO
与磷脂酶C共表达的葡萄糖异构酶和单独表达的葡萄糖异构酶在比活、最适温度、最适pH等性质方面没有明显差别,表明在共表达的过程中,葡萄糖异构酶在被释放出细胞前已经正确地完成了整个蛋白质合成过程,并且不存在折叠不完全等异常情况。
2.4 磷脂酶C与葡萄糖异构酶共表达菌株3 L罐小试
为了考察葡萄糖异构酶与磷脂酶C的共表达是否能够应用于工业生产,用菌株pETDuet/plc/glu进行3 L发酵罐培养。按照1.3.7节所述方法进行pETDuet/plc/glu菌株的3 L罐小试。接罐后约6.5 h,培养基中的甘油耗尽,开始补料。采用指数流加法进行流加补料,细胞干重约为25 g/L时,开始流加乳糖诱导。诱导开始后,每3小时取样一次,结果见图6。培养基中的酶活和细胞干重随着诱导时间的延长逐渐增加。当乳糖诱导约为18 h时,细胞干重不再明显增加;当乳糖诱导约为24 h时,培养基中酶活不再明显增加,此时酶活达到了17.7 U/mL,是摇瓶发酵菌株培养基酶活的5.2倍,释放到培养基中的酶活比例约为85%,电泳结果见图7。
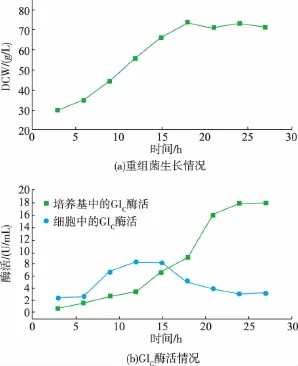
图6 3 L罐小试菌体干重变化和产酶曲线Fig.6 Growth and enzyme production by recombinant E.coli BL21(DE3)under the fermentation condition
对重组磷脂酶C和葡萄糖异构酶共表达的菌株进行的摇瓶发酵和3 L罐小试结果说明,葡萄糖异构酶成功实现了胞外表达,并且这一过程具有表达效率高、发酵过程易调控、后期分离纯化简便等优势,有利于工业化制备葡萄糖异构酶。

图7 3 L罐小试GIC蛋白质电泳图Fig.7 SDS-PAGE analysis of extracellular fraction under the fermentation condition
3 结语
本研究通过共表达对细胞膜有伤害作用的磷脂酶C成功实现了天然胞内蛋白葡萄糖异构酶的胞外表达,并且获得了较高的表达效率。然而过程中的一些实验条件包括发酵条件等只是初步的探究,并未进行深入的优化,仍然存在进一步发掘的潜力。在未来的研究中,可以进一步优化实验工艺,尤其是发酵工艺,以获得更高的胞外酶活、更好的分泌效率以及更短的发酵周期等,为葡萄糖异构酶工业化规模制备奠定基础。
[1]YOON S H,KIM S K,KIM J F.Secretory production of recombinant proteins in Escherichia coli[J].Recent Pat Biotechnol,4(1):23-29.
[2]JONG W S P,SAURI A,LUIRINK J.Extracellular production of recombinant proteins using bacterial autotransporters[J].Curr Opin Biotech,2010,21(5):646-652.
[3]CHOI J H,LEE S Y.Secretory and extracellular production of recombinant proteins using Escherichia coli[J].Appl Microbiol Biot,2004,64(5):625-635.
[4]MERGULHAO F J M,SUMMERS D K,MONTEIRO G A.Recombinant protein secretion in Escherichia coli[J].Biotechnol Adv,2005,23(3):177-202.
[5]SHIN H D,CHEN R R.Extracellular recombinant protein production from an Escherichia coli lpp deletion mutant[J].Biotechnol Bioeng,2008,101(6):1288-1296.
[6]CHIEN L J,LEE C K.Synergistic effect of co-expressing D-amino acid oxidase with T7 lysozyme on self-disruption of Escherichia coli cell disruption[J].Biochem Eng J,2006,28(1):17-22.
[7]MORITA M,ASAMI K,TANJI Y,et al.Programmed Escherichia coli cell lysis by expression of cloned T4 phage lysis genes[J]. Biotechnol Progr,2001,17(3):573-576.
[8]SU Lingqia,CHEN Sheng,YI Li,et al.Extracellular overexpression of recombinant Thermobifida fusca cutinase by alphahemolysin secretion system in E.coli BL21(DE3)[J].Microbial Cell Factories,2012,11(1):8.
[9]DE Maria L,VIND J,OXENBOLL K M,et al.Phospholipases and their industrial applications[J].Appl Microbiol Biot,2007,74(2):290-300.
[10]孟庆飞,温其标.磷脂酶C水解大豆油磷脂提高油脂精炼率的研究[J].中国油脂,2006,31(1):36-38.MENG Qingfei,WEN Qibiao.Improving refining yield of crude oil by using phospholipase C to hyolrolyze soybean gwms[J]. China Oils and Fats,2006,31(1):36-38.(in Chinese)
[11]刘菲菲,张梁,顾正华,等.蜡状芽孢杆菌磷脂酶C基因在大肠杆菌中的异源表达[J].食品科学,2013,34(11):182-187. LIU Feifei,ZHANG Liang,GU Zhenghua,et al.Cloning and heterologous expression of phospholipase C gene from Bacillus cereus in E.coli[J].,2013,34(11):182-187.(in Chinese)
[12]CALIK P,ANGARDI V,HAYKIR N I,et al.Glucose isomerase production on a xylan-based medium by Bacillus thermoantarcticus[J].Biochem Eng J,2009,43(1):8-15.
[13]朱国萍,程阳,宫春红,等.葡萄糖异构酶的生物工程研究进展[J].生物化学与生物物理进展,2000,27(2):127-131. ZHU Guoping,CHENGYANG,GONG Chunhong,et al.Progress in biological engineering of D-glucose isomerase[J].Progress in Biochemistry and Biophysics,2000,27(2):127-131.(in Chinese)
[14]ZHANG Fan,DUAN Xuguo,CHEN Sheng,et al.The addition of Co2+enhances the catalytic efficiency and thermostability of recombinant glucose isomerase from Thermobifida fusca[J].Process Biochem,2013,48(10):1502-1508.
[15]CHENG Jing,WU Dan,CHEN Sheng,et al.High-level extracellular production of r-cyclodextrin glycosyltransferase with recombinant Escherichia coli BL21(DE3)[J].Journal of Agricultural and Food Chemistry,2011,59:3797-3802.
Co-Expression of Phospholipase C and Glucose Isomerase Which Promoting the Extracellular Expression of Glucose Isomerase in E.coli
JIANG Qi, SU Lingqia, WU Jing, CHEN Sheng*
(State Key Laboratory of Food Science and Technology,JiangNan University,Wuxi 214122,China)
Phospholipase C(PLC)hydrolyzes the phospholipids in the membrane.The hydrolysis was partial,which would enhance the cell membrane permeability,and then proteins in the cytoplasm were released into culture medium.In this study,PLC was co-expressed with glucose isomerase(GI).In shake flask fermentation of recombinant E.coli BL21(DE3),the activity of GICin culture supernatant was 3.4 U/mL,which was 93%of the total enzyme activity in culture supernatant and cytoplasm.GICwas released into culture medium.The GICwas purified and characterized.The specific activity of GICwas 12.1 U/mg,the optimum temperature was 80℃,and the activity of GICwas maximal at pH 10.The characteristics of GICwere similar to GIO,In addition,The enzyme activity reached 17.7 U/mL in 24 hours by utilizing fed-batch strategy in 3 L fermentor.
phospholipase C,glucose isomerase,co-expression,released into culture medium
Q 814
A
1673—1689(2017)03—0236—07
2015-03-06
国家杰出青年基金项目(31425020);江苏省自然科学基金项目(BK20140132)。
*通信作者:陈 晟(1981—),女,江苏常州人,工学博士,副教授,硕士研究生导师,主要从事工业微生物、酶工程、发酵工程与技术方面的研究。E-mail:chensheng@jiangnan.edu.cn
姜琪,宿玲恰,吴敬,等.共表达磷脂酶C促进葡萄糖异构酶在大肠杆菌中的胞外表达[J].食品与生物技术学报,2017,36(03):236-242.
